ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(発酵乳、腸内細菌の科学:研究の最前線) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
腸内微生物叢と酸化ストレスの相互作用: 神経変性と神経保護に関する展望 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Shruti Shandilya et al., |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 概要 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
背景 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 脳機能に対する腸内微生物叢の影響に関する最近の研究は、それらの間の関係に関する重要な情報を収集するのに役立ちました。 神経障害の病因は、腸-脳軸の調節不全に関連していることがわかっています。 一部の腸内細菌の代謝産物は、神経変性の最も重要な危険因子の 1 つである活性酸素種レベルの増加に直接関連していることがわかっています。 それらの病的な関連性に加えて、腸内細菌の代謝産物は、これらの生命を脅かす脳障害の発症を減らすのに重要な役割を果たすこともわかっています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| レビューの目的 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 最近行われた研究では、腸内微生物叢と脳の間の 2 つの最も重要な関連性が明らかになりました。「腸内微生物叢 - 酸化ストレス - 神経変性」と腸内微生物叢 - 抗酸化物質 - 神経保護です。 このレビューは、神経変性に関与する腸内微生物叢を介した酸化ストレスに焦点を当て、神経保護における腸内微生物叢とその代謝産物の関与を示す研究に焦点を当て、実施された集団研究の深い洞察を読者に提供することを目的としています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| レビューの重要な科学的概念 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| このレビューは、3 つの主要な主要概念に焦点を当てています。 第一に、臨床および前臨床分野からの証拠の増加は、機能不全の神経学的プロセスをもたらす腸内微生物叢媒介酸化ストレスの影響を示しています。 したがって、酸化ストレスに対する脳の脆弱性に影響を与える腸内微生物叢の潜在的な役割、およびアルツハイマー病とパーキンソン病の新進の原因について説明します。 第二に、腸内微生物叢の寄与する役割は、それ自体の代謝産物を介して、または二次代謝産物を生成することによって酸化ストレスと炎症を軽減する上で観察されており、抗酸化および抗炎症プロバイオティクスによる腸内微生物叢集団の調節も有望な神経回復力を示しています。 第三に、シリコツール(訳者注:コンピューターを用いてを意味する)とデータベースの高スループットは、腸内微生物叢、その代謝物、脳の健康の相関関係も提供するため、魅力的な視点と治療オプションの有望な新しい道を提供します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| レビューの図解 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
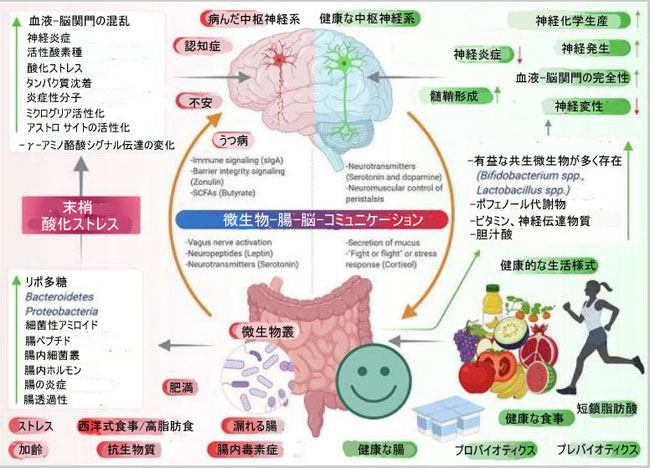 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 目次(クリックして記事にアクセスできます) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1.序章 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.活性酸素種と酸化ストレス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.生理学的条件下での脳における活性酸素種の役割 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.なぜ脳は酸化ストレスに弱いのか? | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.腸内細菌叢、酸化ストレスおよび神経変性 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.1.生理学的条件下での腸-脳軸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.2.腸内細菌叢が媒介する酸化ストレスと神経変性 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.3.アルツハイマー病における腸内微生物叢媒介酸化ストレス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.4.パーキンソン病における腸内微生物叢媒介酸化ストレス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.5.腸内微生物叢と外傷性脳損傷 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.中枢神経系の抗酸化物質と神経保護 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7.神経保護における腸内微生物叢 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.細胞特異的応答による神経保護における腸内微生物叢代謝産物 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.1宿主分子との腸内微生物叢の相互作用 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.1.1.胆汁酸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.2.食物分子との腸内微生物叢の相互作用 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.2.1.アミノ酸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.2.2.食物繊維 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.2.3.ポリフェノール | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9.ビタミンBとビタミンK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.プロバイオティクスの抗酸化作用と抗炎症作用 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 11.腸-脳研究を進めるための in silico戦略 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 12.結論 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
1.序章 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 体の第 2 の脳としても知られる腸神経系 (ENS) は、胃腸 (GI) 管と中枢神経系 (CNS) 間の双方向通信を理解する上で重要なリンクとして機能します [1]。腸ニューロンは、腸の健康維持に関与するニューロン (迷走神経)、内分泌、および免疫経路を介して中枢神経系と通信します [2]。ただし、このコミュニケーションを調節できる調節因子は腸内細菌叢です。腸内微生物集団 (真菌、古細菌、ウイルス、細菌、寄生虫) の中で、特に腸内細菌は体内のヒト細胞の数と同じ数を構成し [3]、興味深いことに、ヒトの肝臓と同等の代謝能力を持っています [4]。この分野での継続的な研究は、免疫系の機能 [5] や調節代謝 [6] だけでなく、さまざまな器官の発達においても腸内微生物叢が非常に重要であることを示しています [7]。食事[8]、薬物[9]、年齢や遺伝学を含む宿主因子[10]などのさまざまな環境要因は、腸内細菌叢の組成を変化させるだけでなく、それらのシグナル伝達活動の変化を引き起こす可能性があります。また、粘膜表面で分泌される最も豊富な抗体である免疫グロブリン A (IgA) は、腸内の共生細菌を覆い、多様で安定した腸内細菌群集を維持します [11]、[12]。さらに、腸内微生物叢は、腸および血管内皮 (血液脳関門) タイトジャンクションを調節するために必要な血清タンパク質ゾヌリンに影響を与えることが知られています [13]。腸内細菌叢の変化は、ゾヌリン経路に直接影響を与え、リーキーガットを引き起こします [13]。さらに、腸内細菌叢は、腸の蠕動運動を制御する要因の 1 つです [14]。内分泌因子の中で、コルチゾール値の上昇は腸内細菌叢の寄与因子として観察されており、ストレスやうつ病の原因と考えられています [15]。このように、腸内微生物叢は、環境から来る膨大な数の化学信号を感知、調節、循環させることにより、人間の健康に直接影響を与えます。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内細菌と神経疾患との関係は、1900 年代に Elie Metchnikoff らによって最初に仮説が立てられ、現在では多くの研究グループによって認識されています。神経変性疾患 (NDD) は、神経系に直接影響を与えることが知られているさまざまな免疫、代謝、および神経化学的要因を生成および変更する能力を腸内の微生物が持っているという事実を無視して、神経系の欠陥によって引き起こされると最初に考えられていました [16]。神経変性疾患は主に、酸化的損傷、活性酸素種 (ROS) 産生の増加、神経炎症、および破壊的なエネルギー代謝によって引き起こされますが、一方で腸内微生物群にも影響を与えます [17]。驚くべきことに、腸内微生物組成は、健康状態から病的状態への身体の代謝の変化に伴って変化します [18], [19]。これは、宿主と環境の間の腸内微生物叢の交差点、およびさまざまな神経学的および心理的障害との病的な関連を示しています。 腸内細菌叢と神経炎症は、さまざまな神経障害の病態生理学における一貫した要因です。 このレビューでは、神経変性における腸内微生物叢を介した酸化ストレスの役割を紹介することを強調しました。これには、活性酸素種産生のメカニズムの説明、脳が酸化ストレスの影響を受けやすい理由が含まれます。 また、アルツハイマー病 (AD)、パーキンソン病 (PD)、外傷性脳損傷 (TBI) に焦点を当てて、腸内微生物代謝物が酸化ストレスによって誘発される脳の損傷にどのように影響するかを調べます。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 神経変性疾患の病理学における腸内微生物叢の役割にもかかわらず、当然のことながら、腸内微生物叢は、食物繊維、ポリフェノール、または胆汁酸やステロイド ホルモンなどの宿主分子の変換からそれらによって生成された代謝産物を放出することによって、脳を損傷から保護する可能性もあります。腸内のそれらの組成は、神経回復力を促進するためにプレバイオティクスによって調節することができます [20]。 腸内微生物叢の神経保護の役割は、最近の研究で十分に文書化されており、韓国の発酵食品から分離された Lactobacillus buchneri KU200793 がより高い抗酸化活性を示したことが観察されました。 また、SH-SY5Y(訳者注:ヒト神経芽細胞腫)細胞を 1-メチル-4-フェニルピリジニウム (MPP+) から保護することができ、そのプロバイオティクスおよび神経保護効果を示唆しています [21]。 同様に、Lactobacillus delbrueckii ssp bulgaricus B3 および Lactobacillus plantarum GD2 から分離されたエキソポリサッカライドが、SH-SY5Y 細胞を アミロイドβ(1-42) 誘導アポトーシスから保護したことが報告されています [22]。 アルツハイマー病の薬理学的治療のための有望な天然化学成分としての役割を強化します。 さらに、Rumnicoccus albus の加熱殺菌株による SH-SY5Y およびマウスモデルの治療は、過酸化水素 (H2O2) で処理された SH-SY5Y 細胞およびヒ酸ナトリウムで処理された動物モデルにおいて、神経保護効果を示し、活性酸素種レベルを低下させ、スーパーオキシドジスムターゼ (SOD) およびグルタチオン (GSH) レベルを増加させるのに非常に効果的であることがわかった[23]。 同様に、Lactobacillus plantarum MTCC1325 の抗アルツハイマー病作用は、D-ガラクトース (D-Gal) によって誘発されたアルビノラットで研究されています [24]。 L. plantarum は、D-ガラクトースの記憶障害およびマウスのスコポラミン誘発性アルツハイマー病から保護することが示されています [25]。 まとめると、これらの研究は、腸内微生物叢の有望な役割、それらの抗酸化的役割、そしてその後の神経保護的役割を反映しています。これらの基本的な発見に基づいて、神経保護における抗炎症性および抗酸化性プロバイオティクスとともに、腸内微生物叢、その代謝産物の役割について説明します。 また、腸と脳の相互作用を研究するために利用されるデータベースと in silico 戦略に関する情報を収集しました。 全体として、このレビューは、酸化ストレス誘発性神経変性と腸内微生物代謝産物を介した抗酸化メカニズムに基づく神経保護における腸内微生物叢の二重の役割について深い洞察を与えます。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2.活性酸素種と酸化ストレス |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 酸化ストレスは、抗酸化物質レベルよりも活性酸素種レベルの増加による細胞内のレドックスシグナル伝達経路の破壊に対応しています。 この不均衡状態は有害な影響をもたらし、多くの神経疾患の主な原因となっています。 好気性代謝に関与するすべての化学反応は、活性酸素種として知られる不安定で短命な反応性中間生成物の形成をもたらします [26]、[27]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 生体分子の酸素 (O2) は、完全に還元することができない 2 つの不対電子を持っているため、その不完全な還元により、過酸化水素、スーパーオキシド アニオン、一酸化窒素、ペルオキシナイトライト アニオン、ヒドロキシルおよびペルオキシル ラジカルのような高度に求電子的で寿命の短い活性酸素種が形成されます [28]。 . 活性酸素種は、活性酸素種生成酵素を介して、または薬物、毒素、放射線などの外因性因子の存在下で、通常の細胞プロセスの中間体として生成されます [29]。神経組織は代謝率が高いため、他の臓器に比べて大量の活性酸素種を生成します。ミトコンドリアは、脳内の活性酸素種生成の主要な細胞部位と考えられており、ATP 生成 (ミトコンドリアの酸化的リン酸化) のプロセス中に、副産物としてアニオン スーパーオキシドが生成され、スーパーオキシドジスムターゼによって 過酸化水素と 酸素に急速に変換されます [30]。 酸素の量が多いほど、スーパーオキシドの形成が多くなり、漏れやすい電子伝達系 (ETC) (複合体 I および III) からの電子の追加により、過酸化水素やヒドロキシルラジカルなどの 活性酸素種がさらに増加します [31]。ミトコンドリア活性酸素種産生はまた、強力なシナプス伝達がスーパーオキシド産生を促進するため、ニューロン活動を示します [32]。一方で、ミトコンドリア活性酸素種は細胞内カルシウム (Ca2+) レベルによって調節できますが [33]、さらに、ミトコンドリア活性酸素種産生の増加は、ミトコンドリア膜電位の増加にも関連しています。ミトコンドリア内膜、ミトコンドリアマトリックス酵素に加えて、アコニターゼは、フェントン反応で鉄硫黄クラスターによって促進され、過酸化水素をヒドロキシルラジカルに変換することにより、活性酸素種の生成にも寄与します。外部 NADH デヒドロゲナーゼ、プロリン デヒドロゲナーゼ、ジヒドロオロト酸デヒドロゲナーゼ、複合体 IV などの他の多くの酵素も、ミトコンドリア活性酸素種産生の寄与因子として報告されています [34]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ミトコンドリア内膜に加えて、モノアミンオキシダーゼ (MAO) はミトコンドリア外膜に存在し、ニューロンを含むほとんどの細胞型に存在し、活性酸素種の主要な供給源の 1 つです。 モノアミンオキシダーゼは、分子酸素を利用して分子からアミン基を除去することにより、補因子 FAD (フラビン アデニン ジヌクレオチド) を必要とするモノアミンの酸化的脱アミノ化を触媒し、そこで反応の副産物として 過酸化水素が生成されます [35]。 すなわち モノアミンオキシダーゼ-A と モノアミンオキシダーゼ-B は、グリア細胞と神経細胞の酸化還元状態を調節することが知られています。 モノアミンオキシダーゼ-A は主にカテコールアミン作動性ニューロンに見られ、ノルアドレナリンとセロトニンの酸化に関与していますが、モノアミンオキシダーゼ-B は特にセロトニン作動性ニューロンとグリア細胞で発現し、β-フェニルエチルアミンを酸化します [36]。第三に、ニューロンに存在する一酸化窒素合成酵素 (NOS) nNOS のアイソフォームも脳内の 活性酸素種の供給源の 1 つであり、Ca2+ 結合タンパク質であるカルモジュリンによって制御されます。一酸化窒素合成酵素は L-アルギニンから L-シトルリンへの酸化を触媒し、補因子として NADPH、テトラヒドロビオプテリン、および酸素を利用して一酸化窒素 (NO) を生成します [37]。 一方では、一酸化窒素はシナプス伝達を調節する重要なシグナル伝達分子として機能しますが、スーパーオキシドと相互作用して反応性の高いペルオキシナイトライト化合物を形成することにより、レドックスの恒常性維持を妨害することもできます。 これらは、細胞内でニトロソ化ストレスを引き起こすことに直接関与しており、それぞれ低濃度および高濃度でアポトーシスおよびネクローシス細胞死に関連しています [37]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 他の酵素に加えて、生理学的状態における脳内の活性酸素種の主要な内因性供給源の 1 つには、NADPH オキシダーゼ (NOX) が含まれます。これは、NADPH の酸化を触媒し、その主な生成物としてスーパーオキシドを生成します。 NOXは主に原形質膜と多形核好中球のファゴソームに位置することがわかっていますが、マウスとラット組織の以前の免疫組織学的評価では、脳の皮質と海馬領域に豊富に存在することが示されました [38]、[39]。 Ca2+ は NOXの主要な活性化因子として作用し、ニューロンにおける酵素複合体のシナプス後局在化につながり、ニューロン活動におけるNOXの関与を示しています [40]。 NOXの 7 つのパラログ、すなわち NOX (1-5)、デュアル オキシダーゼ DUOX (1 および 2) が報告されており、これらはサイズとドメイン構造が異なりますが、主に活性酸素種生成に関与しています。 タンパク質は活性化され、成熟、安定化、および他のタンパク質との相互作用で膜を横切る転位を受けます。たとえば、NOX (1–3) は G タンパク質 Rac とともに p22 (phox) 膜貫通タンパク質と相互作用することによって活性化されます。 同様に、NOX-4 は p22 (phox) 膜貫通タンパク質のみと相互作用することによって活性化されますが、NOX-5 および DUOX (1 および 2) は Ca2+ に直接結合することによって活性化されます。 活性化されると、NOX (1–3) と NOX-5 は主にスーパーオキシドの生成を触媒し、NOX-4、DUOX (1 および 2) は 過酸化水素の直接生成に関与します [41]。 NOX パラログは、脳の皮質、海馬、小脳に広く分布していることがわかっており、NOX-2、NOX-3、NOX-4 が最も顕著です。 以前の報告では、ミトコンドリア活性酸素種と NOX-活性酸素種の間の相乗的関係も明らかにされており、互いの活性酸素種生成をサポートしています [42]。 現在までに、アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症 (ALS)、ハンティングトン病 (HD) などの慢性中枢神経系疾患の進行を媒介する NOX の極めて重要な役割が研究によって明らかにされています [41]。 したがって、アイソフォーム選択的 NOX 阻害剤の開発は、急性および慢性中枢神経系障害の治療のための有望な治療アプローチとなり得る。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 細胞質内では、リポキシゲナーゼなどの非ヘム鉄酵素が分子酸素の存在下でアラキドン酸の過酸化を触媒し、スーパーオキシドとヒドロキシル ラジカルを生成します [43]。 キサンチンオキシダーゼ、シトクロム P450 モノオキシゲナーゼ、シクロオキシゲナーゼ、D-アミノオキシダーゼなどの細胞質内の他の多くの酵素も重要なかっせ栄酸素種プロデューサーです [44]。 ミトコンドリアと連携して、ペルオキシソームのようなオルガネラも活性酸素種産生の場所であり、そこではグリコール酸オキシダーゼとキサンチンオキシダーゼによって触媒される脂肪酸のベータ酸化がスーパーオキシドと過酸化水素をもたらす [27]。 また、活性酸素種産生の増加は、展開タンパク質応答 (UPR) により小胞体で発生します [45]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 酸化ストレスは、脂質過酸化、タンパク質の酸化、核酸損傷という 3 つの主な反応を媒介することにより、細胞損傷を引き起こします [46]。 実際、酸化ストレスは加齢中の正常な生理学的プロセスの一部と考えられていますが、アルツハイマー病、パーキンソン病、ンティングトン病、虚血性脳卒中、うつ病、硬化症などの脳の慢性疾患に関与していることが知られています [46]。 さらに、2型糖尿病(T2D)、非アルコール性脂肪肝疾患、非アルコール性脂肪性肝炎、肥満、心血管疾患、癌などの生活習慣に関連した代謝障害において重要な役割を果たしています[47]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.生理学的条件下での脳における活性酸素種の役割 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 高レベルの活性酸素種は有害であると考えられていますが、生体分子の損傷を引き起こし、幅広い細胞機能障害を引き起こしますが、安全な安定したレベルでは、活性酸素種は有用な生物学的役割を果たします. 通常の生理学的条件下では、細胞外活性酸素種は自然免疫応答を誘発することによって感染症を軽減するのに役立ちます。 一方、細胞内で生成されるフリーラジカルは、シグナル伝達経路、アポトーシス、および酸化ストレスに対する防御システムを刺激するのに役立ちます。 活性酸素種は、酸化ストレスにつながる炎症を引き起こす核転写因子 NF-κB の活性化にも重要な役割を果たします。 次亜塩素酸 (HOCl) のようなフリーラジカルは、病原体に対する強力な酸化剤として使用されるミエロペルオキシダーゼの作用によってリソソームで生成されます。 したがって、活性酸素種は、細胞内シグナル伝達を調節するシグナル伝達経路の強力な参加者として機能します [27]、[44]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 中枢神経系に関しては、生理学的条件下での多くの反応で副産物として生成される 活性酸素種は、細胞内シグナル伝達経路の調節に役立つだけでなく、細胞の増殖、分化、および成熟も調節します [48]。以前の報告では、活性酸素種の産生とレドックスのバランスが、前駆神経前駆細胞からの神経細胞の分化と軸索の形成を仲介するのに役立ち [49]、ニッチでの神経細胞の拡大にも役立つことが示されました [50]。さらに、酸化還元シグナル伝達 (活性酸素種および酸化状態) は、酸化還元とともに (NF-κB)、活性化 T 細胞の核因子、活性化タンパク質 1 (AP-1) などの転写因子の機能も調節することが観察されています。したがって、チロシンリン酸化タンパク質 PKC の状態は、神経発生に関与するシグナル伝達カスケードに影響を与える上で重要な役割を果たします [51]。また、過酸化水素のような活性酸素種は、細胞内 Ca2+ シグナル伝達を強化することにより、皮質ニューロンの興奮性を調節することが観察されています [52]。同様の研究では、過酸化水素が皮質ニューロンと PC12 細胞の ERK と cAMP 応答要素結合タンパク質 (CREB) のリン酸化を増加させることが観察されました [53]、[54]。これは、活性酸素種が神経系のシグナル伝達カスケードに影響を与える上で重要な役割を果たし、シグナル伝達経路のメッセンジャーとして機能できることを示しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 活性酸素種は、海馬、大脳皮質、視床下部、扁桃体、脊髄などの脳のさまざまな部分で二次メッセンジャーとして機能し、シナプス可塑性の維持に役立ちます [55]。 活性酸素種は、哺乳類の学習と記憶に関連する海馬の長期増強 (LTP) にも必要であることがわかっており、シナプス増強における活性酸素種の関与が示されています [56]、[57]。 活性酸素種はまた、痛みの調節の感情的側面を担う脳の領域である扁桃体の中心核の興奮性の増加に関与することにより、痛みに関連する行動にも影響を与えます [58]。同様に、脊髄では、神経因性および炎症性疼痛に関連する神経可塑性プロセスも、シグナル伝達分子として作用する活性酸素種によって制御されます [59]。動物研究からの報告は、NOX を介した活性酸素種の直接生成が、特に海馬と視覚野でシナプス可塑性メカニズムを維持するのに役立つ重要な生理学的プロセスとして作用することを示しました [60]。さらに、過酸化水素の用量依存効果は、レドックス変化をもたらし、シナプス可塑性を調節する必須のプロセスであることが観察された [61]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
4.なぜ脳は酸化ストレスに弱いのか? | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 基底酸素の約 20% が脳によって消費され、2,500 ~ 3,000 億個のグリアによってサポートされている、数兆個のシナプスによって接続された約 860 億個のニューロンの ATP 主導の活動をサポートします [62]。 脳が酸素を奪われると毎分ニューロン (~190 万) とシナプス (~1400 万) は死に始めることが報告されている [63]。 酸素 は脳の機能に不可欠ですが、酸化ストレスがどのように神経変性を引き起こすかという事実には常にあいまいさがあります。酸化還元シグナル伝達中に生成される酸素由来のフリーラジカルおよび非ラジカル種は、脳の健康を等しく維持し、その潜在的な正の機能を示しています [64]。 NADPH オキシダーゼ (NOX-2) の作用によって生成される 酸素および過酸化水素が、PI3K/AKT 経路を介して神経前駆細胞の増殖の維持に関与していることが報告されています [49]。 また、NOX-2 の欠失は、脳の認知機能の調節におけるその重要性を示している [65]。 最近、NOX-2由来の過酸化水素が、その再生においても軸索経路発見をサポートする内因性化学誘引物質として作用することが報告されている[66]。 したがって、多様な反応種が脳によって使用され、酸化ストレスの影響を受けやすくなるシグナル伝達機能を実行します。脳を酸化ストレスに対してより脆弱にする生化学的イベントは他にもたくさんあります。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| a) 双方向のシナプス可塑性を維持するには、活動電位中の Ca2+ トランジェントが必要です。 膜を横切る高い Ca2+ トランジエントが中断されると、遊離細胞内 Ca2+ 濃度が上昇し、ニューロンの NOS (NO. の産生)、ホスホリパーゼ A2、および細胞骨格に損傷を与えるカルピンが活性化されます。 高一酸化窒素はシトクロム C オキシダーゼにも結合し、ミトコンドリア呼吸を阻害します。 さらに、一酸化窒素は (機能不全のミトコンドリアからの) 酸素.- と反応して ONOO– を形成します。 ミトコンドリアの Ca2+ 過負荷は、ミトコンドリア透過性遷移孔 (mPTP) を介して Ca2+/過酸化水素の流出を媒介することにより、ATP 生成を無効にします。 これはネクロトーシスにつながります。 このように、Ca2+ 恒常性の崩壊により、脳は酸化ストレスを受けやすくなります [67], [68]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| b) グルタミン酸はニューロンの興奮毒性アミノ酸として作用し、大量に摂取すると壊死によって細胞を損傷します。 この損傷により、細胞外環境に大量のグルタミン酸が放出され、隣接するニューロンの受容体にも結合して、細胞内で Ca2+ および Na+ が持続的に放出されます。 ONOO– のような反応性種は、グルタミン合成酵素を不活性化することにより、グルタミン酸からグルタミンへの変換を阻害します。 また、グルタミン酸は、Xc-トランスポーターを介した細胞内グルタミン酸から細胞内システインへの交換を阻害し、グルタチオンの枯渇を引き起こし、フェロトーシスを介したニューロンの死につながります [69]、[70]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| c) 脳は、多くの酵素の補因子として作用する Fe2+ や Cu+ などのレドックス遷移金属イオンが豊富です。 脳損傷を引き起こすストレス要因は、フリーラジカル反応を触媒できる金属イオンを放出します。 さらに、放出された鉄は、脳脊髄液 (CSF) に鉄結合能力がほとんどまたはまったくないため、長期間存在します [71]、[72]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| d) ドーパミン、セロトニン、ノルエピネフリンなどの神経伝達物質は、自動酸化を受けて 活性酸素種を生成します。 簡単に説明すると、ドーパミンは酸素と反応してセミキノンと酸素を生成し、酸素はさらに酸素と反応してキノンを生成します。 そのように生成されたキノンは、酸素によってキノールと過酸化水素に再酸化され、そこで Mn2+ が過酸化水素と反応して OH を生成します。 このような自動酸化は、ミトコンドリアとリソソームの機能不全を引き起こします [73], [74]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| e) 脳は、グルコースによって誘発される酸化ストレスにも敏感です。 ペントースリン酸経路にグルコースの大部分を利用するために、ニューロンは律速解糖酵素であるホスホフルクトキナーゼを分解します。 解糖速度の欠如は、タンパク質の糖化と最終糖化産物 (AGE) の形成をもたらします。最終糖化産物は、炎症による酸化ストレスによってタンパク質とミトコンドリアの機能を損なう [75]、[76]、[77]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| f) 脳は多価不飽和脂肪酸、特にドコサヘキサエン酸 (DHA) の含有量が高く、脂質の過酸化と脳による過酸化脂質の使用により、酸化ストレスを受けやすくなります。 4-ヒドロキシノネナールのような脂質過酸化の生成物は、Ca2+ レベルを増加させることによってグルタミン酸トランスポーターを不活性化するため、神経毒性があります。 脂質過酸化物はまた、α-ケトグルタル酸デヒドロゲナーゼを不活性化し、血管収縮剤として作用し、プロテアソームに損傷を与え、アルツハイマー病のような神経変性疾患の一貫した要因であることがわかっています[78]、[79]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| g) 脳の常在免疫細胞であるミクログリアは、脳の発達と機能にとって重要ですが、食作用の通常の活動中に、酸素- および他の反応種を生成します。 アクティブなミクログリアは NOX-2経由して酸素- を生成します。 したがって、ミクログリアの活動は、総酸素バイオアベイラビリティに依存し、酸素を生成するためにより多くの酸素を消費することによるシナプスへの損傷を示しています。 過酸化水素や一酸化窒素どの反応性種は、その部位にミクログリアを引き付けて、神経変性を引き起こす局所的な炎症を引き起こします [80]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| h) 脳は、他の組織と比較して適度な抗酸化防御システムにより、レドックスの恒常性が乱れがちです。 ニューロンのカタラーゼ含有量は、肝細胞のそれよりもはるかに低い (50 倍)。 さらに、ペルオキシソーム内にそれらが存在すると、他の細胞内コンパートメントで生成された過酸化水素に作用するその活性が制限されます。 同様に、ニューロンは非常に低いレベルのグルタチオンを持っているため、フェロトーシスに敏感になり、求電子剤の代謝に抵抗します [81]、[82]、[83]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| i) ヘモグロビンが過剰な過酸化水素と反応して酸化促進剤の鉄イオンとヘムが放出される場合、ヘモグロビンは脳にとって神経毒性があると見なされます。 遊離ヘムは、脂質過酸化の最強のプロモーターです。 さらに、その一酸化窒素との結合は血管収縮を引き起こします [84]、[85]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| j) 脳には低レベルのシトクロム P450 酵素、CYP2E1 が含まれていますが、 反応を触媒しながら電子が漏れるため、脳は酸化ストレスを受けやすくなります。 研究では、エタノールの消費と喫煙によりそのレベルが上昇する可能性があることが示されました [86]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| k) ポリ ADP リボース ポリメラーゼ (PARP-1) のような DNA 修復酵素は、NAD+ を切断し、ADP リボースを核タンパク質に結合することによって DNA 損傷を修復します。 しかし、この酵素の過剰活性化は、神経保護作用のある NAD+ の枯渇につながり、エネルギー産生を制限し、神経細胞死をもたらす一過性受容体電位メラスタチン (TRPM2) Ca2+ チャネルを開く [87]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| l) ヒストンタンパク質によって保護されていない一本鎖であるため、RNA は酸化に対してより脆弱です。 酸化された RNA は、リボソームによるタンパク質合成を停止させ、修復されないままにしておくと、折り畳まれていない切断されたタンパク質になる可能性があります。 酸化還元活性遷移金属とともに酸化された RNA がフェントン反応を触媒することが報告されました。 酸化された CuZn-SOD mRNA が筋萎縮性側索硬化症の前臨床徴候であることも報告されています。 ただし、神経変性におけるその強力な役割を調査する必要があります [88]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
5.腸内細菌叢、酸化ストレスおよび神経変性 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
5.1.生理学的条件下での腸-脳軸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 胃腸 (GI) 管には、何兆もの共生微生物と約 1000 種の共生微生物が含まれており、膜バリア機能の維持に重要な役割を果たしています [2]。 これらの微生物は小腸と結腸に常駐しており、宿主生物内の分子の流れを一定に保ち、さまざまな代謝機能を調節している [58]。 微生物群集は生後 2 年以内に宿主の消化管内で安定しますが、その構成は個人によって異なり、年齢、健康状態、遺伝学、ライフスタイルなどの外的要因によって変化する可能性があります [89]。 消化管の管腔側は食事成分と腸内微生物叢にさらされており、さらに、腸組織は免疫細胞の 70% によって収容され、腸と脳をつなぐニューロンによって神経支配されており、腸と脳の間の絶え間ないコミュニケーションが関与しています [2]。 . 腸内微生物叢と脳の間のコミュニケーションには、4 つの主要な経路が含まれます。最初の重要なモードには、消化管の筋肉および粘膜層を脳幹に接続する迷走神経の活性化が含まれます。 最近の報告では、腸内病原体とプロバイオティクスが、迷走ニューロンの活性化を介して脳内のγ-アミノ酪酸 (GABA)、オキシトシン、および脳由来神経栄養因子 (BDNF) シグナル伝達を変化させることにより、不安、摂食、うつ病などの宿主の行動を調節することが示されている [90]。 [91]。 ある報告では、トリプトファンから得られる細菌代謝産物であるインドールが、迷走神経の活性化を介してラットの不安様行動を増加させることが示されましたが、これらの効果を媒介する特定の代謝産物を特定する必要があります [92]。 脳活動に直接的または間接的に影響を与える通信の 2 番目の経路には、腸内壁に存在する腸クロム親和性細胞 (EC) によって放出されるセロトニンを介したシグナル伝達が含まれます。 プロバイオティクスのBifidobacterium sppで治療した場合、うつ病のマウスモデルでセロトニンとセロトニン前駆体レベルが増加し、うつ病状態が改善されることが研究で示されました [93]。 同様に、胞子形成細菌 (Clostridium spp.) からの代謝産物が腸クロム親和性細胞からのセロトニン産生を刺激できることが報告された [94]。 第三に、腸内微生物叢はミクログリアの発生、成熟、活性化に重要な役割を果たしています。 ある研究では、無菌(GF)マウスは、従来のマウスよりも多くの未熟ミクログリアを保有していることが報告されており、さらにBifidobacterium sppで処理した場合。転写活性化を通じてミクログリア細胞を活性化する [95]。 ミクログリア機能の変化は、行動および神経変性疾患で観察されており、ミクログリアを介した神経変性疾患に対する腸内微生物叢の影響を示しています。 腸内細菌叢は、サイトカインやケモカインなどの全身免疫系を介して神経系にも影響を与えます。 研究によると、無菌マウスは従来のマウスと比較して血液-脳関門 (BBB) の透過性が高いため、脳が微生物産物にアクセスしやすくなり、その後神経病理学的状態につながることが示された [96]。 最後になりましたが、腸内微生物叢は化学信号を脳に直接伝達することによって通信します。 たとえば、腸内細菌による食物繊維の発酵は、中枢神経系の神経可塑性を調節することが示されている短鎖脂肪酸 (SCFA) を生成し、マウスの抑うつ行動を改善することも報告されている [97]。 さらに、Bacteroides, Bifidobacterium, Parabacteroides およびEscherichia spp. などの腸内微生物叢は、神経伝達物質 γ-アミノ酪酸を生成することができます。これは、腸内微生物叢が宿主生物の神経伝達物質の濃度を調節することを示唆しています [98]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
5.2.腸内細菌叢が媒介する酸化ストレスと神経変性 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Firmicutes, Bacteroidetes, Actinobacteria, および Proteobacteria. を含む共生細菌の4つの主要な門がヒトの腸に定着します。 その中で、Lactobacillus, Streptococcus, Mycoplasma およびClostridium などのFirmicutesとBacteroidetesが全体の90%を占めています。 腸内の共生細菌と病原性細菌の両方が、ミトコンドリアの活性を調節することによって細胞の活性酸素種を変更することができます [99]。 共生細菌は、マクロファージおよび好中球上の G タンパク質共役受容体 (GPCR) に結合するホルミル化ペプチドを生成し、上皮細胞の炎症を引き起こします。 このプロセスにより、NOX-1 によるスーパーオキシドが生成され、細胞の活性酸素種が増加します [100]。 腸内Lactobacilli と Bifidobacteriumは、硝酸塩と亜硝酸塩を一酸化窒素に変換する能力を持っており、腸上皮を一酸化窒素の豊富な供給源にしています。同様に、Streptococcus と bacilliは、NOS を使用して L-アルギニンから一酸化窒素を生成します [101]。 ナノモル濃度の一酸化窒素は神経保護作用があると考えられており、ノルアドレナリン作動性、非コリン作動性腸管ニューロンの神経伝達物質です。 高濃度では、スーパーオキシドや過酸化水素などの活性酸素および窒素種 (RONS) の生成によって引き起こされる有害な影響をもたらし、神経炎症、軸索変性、神経変性疾患に関連する非常に反応性の高いヒドロキシルラジカルをさらに形成します [18]。 腸内細菌によって生成される短鎖脂肪酸のような有益な代謝産物は、ミトコンドリアの活動に影響を与えることによって活性酸素種を減らすのに役立ちます。これについては、別のセクションで詳しく説明します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
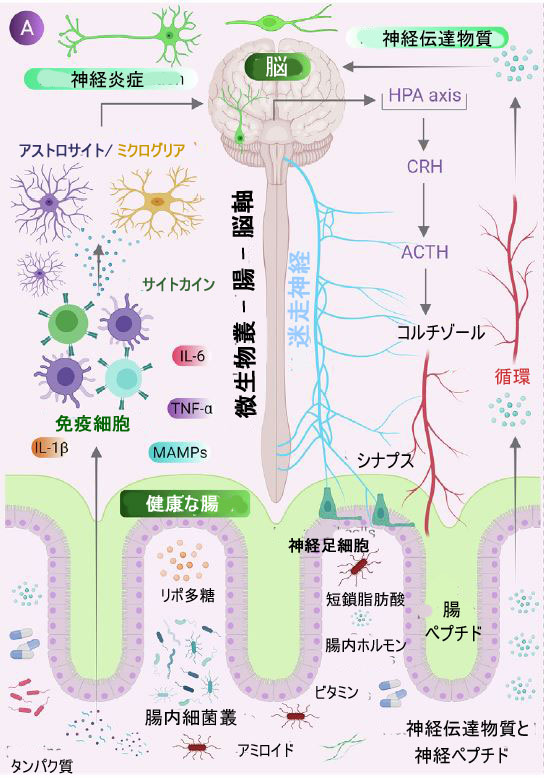
| 膜関連分子パターン (MAMP) は、すべてのクラスの微生物の構造的完全性と基本的な機能を維持し、脳によってさえ検出されます。 これらは、ペプチド、ヌクレオチド、炭水化物、脂質を含む多様な化学基です [102]。 このような分子パターンが宿主によって検出されない場合、それは急性から慢性の炎症を引き起こす可能性があり、脳の発達と機能を変化させることがわかっています。 これらの高度に保存された構造モチーフは、自然免疫系の細胞に存在するパターン認識受容体 (PRR) に結合し、ミトコンドリアの活性酸素種産生と NF-κB 経路の活性化を誘導し、炎症反応を引き起こし、神経ストレスと細胞死を引き起こす [103]。 最近の研究では、細菌の細胞壁成分であるペプチドグリカンが発生中の脳に移行し、遺伝子発現に影響を与え、社会的行動の変化を引き起こすことが報告された [104]。 同様に、グラム陰性菌の細胞壁に存在するリポ多糖は、マウスの胎児の脳の発達、急性うつ病、および認知障害を損なうことがわかった[105]。 さらに、膜関連分子パターンの急性および慢性暴露は、パーキンソン病、自閉症スペクトラム障害 (ASD)、およびシヌクレイノパシー モデルにおける疾患症状の発症の原因となる要因であることが観察された [106]。 膜関連分子パターンとは別に、日和見病原体によって産生される細菌毒素も、宿主の神経系に悪影響を及ぼします。 毒素 B、エンテロトキシン、Clostridium sppによって産生されるイプシロン毒素などの致死毒素は、破壊された 血管-脳関門を介して脳に到達することにより、神経細胞の生存能力を低下させ、神経伝達物質の放出を阻害することが発見された [107]。 Staphylococcus sppが産生するエンテロトキシンとセレウリドおよび Bacillus spp は、迷走神経を刺激することによって嘔吐および病気の行動を誘発することがわかった [108]、[109]。 Salmonella や E. coli などの病原性細菌は、硫黄含有アミノ酸を分解することができるため、腸内で硫化水素 (H2S) を生成します。 硫化水素レベルの増加は、乳酸の増加や ATP 産生の減少、シクロオキシゲナーゼ 2 (COX-2) 活性の阻害 [110]、ミトコンドリアによる酸素消費の減少、炎症誘発性サイトカインの発現増加 [111] など、さまざまな代謝活動の変化を引き起こし、高血圧と神経炎症を刺激することが知られている[112]。 図1は、神経変性における腸内微生物叢媒介酸化ストレスの役割を示しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 図1神経変性における腸内微生物叢の役割 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (A) 腸と脳の間の通信には、神経経路、代謝経路、内分泌経路、および免疫経路が関与します。神経伝達物質、アミノ酸、短鎖脂肪酸 (SCFA)、アミロイド、リポ多糖 (LPS)、微生物関連分子パターン (MAMP) などの腸内微生物分子は、循環を介して宿主免疫系と相互作用し、宿主の代謝と神経系に影響を与え、 、また、脳への腸神経系を介して迷走神経を直接活性化することにより、脳に影響を与えます。ストレスのような状態は、視床下部ニューロンに副腎皮質刺激受容体ハーモン (CRH) を分泌させ、副腎皮質刺激ホルモン (ACTH) の放出を引き起こし、続いてコルチゾールの放出を活性化し、腸の健康に影響を与える腸バリアの完全性に影響を与えます。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (B) 腸内毒素症(Dysbiosis)の状態が発生すると、抗炎症性分子 (短鎖脂肪酸、水素 など) が減少し、炎症誘発性分子 (リポ多糖、アミロイド) にかわります。 また、腸内で有益な細菌が病原菌に変化します。 これにより、腸および血液-脳関門の透過性が増加し、その後末梢免疫応答が増加し、中枢神経系 (CNS) の酸化ストレスが増加します。活性酸素種 (ROS) の産生の増加は、ミトコンドリア、小胞体 (ER)、ニューロンのペルオキシソームなどの細胞オルガネラで観察され、神経毒の凝集とともに神経変性を引き起こします。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| アルツハイマー病やパーキンソン病 などの神経変性疾患の病因には、ニューロン内タンパク質のミスフォールディングとその凝集が関与しています。 さらに、酸化ストレスもその病原因子の 1 つと考えられていますが、正確な下線を引くメカニズムはまだ不明です。 神経変性疾患病理学と腸内微生物叢との関係を示す最初の証拠は、Heiko Braak グループによって示されました。 彼らは、パーキンソン病患者の腸内の粘膜下マイスナー神経叢およびミエンテリック アウエルバッハ神経叢で α-シヌクレイン凝集を発見し、腸内での α-シヌクレイン凝集の開始における腸内微生物叢の役割を示唆している。 その後、シナプスを介して 中枢神経系ニューロンに上昇し、神経変性を引き起こす [113]。 興味深いことに、微生物叢を介したタンパク質障害と神経炎症は、それらの中間的な関係を示す「マプロノーシス」と呼ばれています [114]。 これまでのところ、健康な人と比較して、神経学的疾患の状態で腸内微生物叢の構成が変化していることを多くの研究が示していますが、細菌および細菌因子が疾患の進行にどのように影響するかについてのメカニズムを特定するには、さらなる研究が必要です. ここでは、アルツハイマー病、パーキンソン病、および外傷性脳損傷に焦点を当てて、腸内細菌、酸化ストレス、および 神経変性疾患の間のリンクを描いた最近の研究について説明します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
5.3.アルツハイマー病における腸内微生物叢媒介酸化ストレス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| アルツハイマー病は認知症の主な原因であり、世界中で 5,000 万人を超える人口が罹患しており、その有病率は高齢者で高く、1,000 人あたり約 80 人が 85 歳以上です。 アルツハイマー病の無症候性病理は、記憶喪失や認知障害などの症状が現れる約 20 年前に始まると考えられている [115]。 アルツハイマー病に関連する脳の病理学的変化には、タンパク質アミロイドβの細胞外蓄積(Aβ-アミロイド斑)とタウタンパク質の細胞内蓄積(タウもつれ)が含まれる[116]。 このタンパク質の異常な蓄積は、アミロイドβおよびタウタンパク質のクリアランスのためにミクログリアの活性化につながりますが、その後の老化に伴い、慢性炎症が起こり、神経細胞死を引き起こし、萎縮につながります[117]。 発症の 20 年前に アミロイドβ 蓄積を引き起こし、その後 18 年前に脳によるグルコース消費が減少し、萎縮が疾患発症の 13 年前に始まったというまれな遺伝子変異にもかかわらず [118]、 多くの横断的研究では、アルツハイマー病の病因として修正可能な危険因子が文書化されている[119]。 考えられる危険因子の中で、酸化ストレスと腸内微生物叢の役割は科学界を引き付けており、神経変性プロセスの直接的なもっともらしい結果と見なされています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 脳の高いエネルギー需要に加えて、酸化ストレスとアルツハイマー病の関連性は、脳の抗酸化防御システムの変化、すなわちスーパーオキシドジスムターゼ(SOD)とカタラーゼ酵素の活性とレベルの変化を示した多くの研究に反映されている[120]。 同様に、マロンジアルデヒド、4-ヒドロキシノネナール、F2-イソプロスタン (脂質酸化損傷) などの酸化ストレス バイオマーカー。タンパク質カルボニルと3-ニトロチロシン(タンパク質酸化の生成物)、8-ヒドロキシデオキシグアノシン(核酸酸化)は、血液と脳脊髄液で高レベルで発見された[120]、 また、それらの濃度は、認知障害および脳重量の濃度に比例することがわかった [121]。 アルツハイマー病脳における活性酸素種産生は、ミトコンドリア(シトクロムCオキシダーゼの欠損)[122]、展開タンパク質応答 (UPR)による小胞体[123]、神経斑における金属イオンの蓄積[124]などの細胞小器官の機能不全によっても特徴付けられる。 そして、ミクログリアの過剰活性化とそれに続くNADPHオキシダーゼの過剰発現による[125]。 また、アミロイドβ沈着と酸化ストレスの間には相互関係があります。つまり、アミロイドβ凝集は酸化ストレスを誘発し(ミトコンドリア、小胞体(ER)、ゴルジ装置などのオルガネラでも)、酸化ストレスはAβ蓄積を誘発します[126]。 さらに、ニューロンにおけるタウタンパク質の凝集は、NADH-ユビキノンレダクターゼ活性の低下につながり、活性酸素種産生の増加とミトコンドリア機能障害をもたらす[127]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 最近の事実と数字は、アルツハイマー病が限定的な脳の炎症の結果であるだけでなく、末梢の炎症の結果でもあることを示している[128]。 これは、腸の腸内環境異常が年齢とともに増加する炎症、血液-脳関門の破壊、免疫系の活性化、それに続く神経変性を引き起こすという事実によって裏付けられています。 一方、健康でバランスの取れた腸は、活性酸素種によって引き起こされる有害な影響を減らすのに役立ちます [19]。 アルツハイマー病に罹患している個人は、Bifidobacterium sppやFirmicutesなどの共生細菌の個体数が減少し、Escherichia、Shigella spp、Bacteriodetesの量が増加し、その後炎症とアミロイドβの蓄積が増加することが確認されている[129]。 同様に、広域抗生物質の組み合わせで治療した場合、APP / PS1マウスモデルでアミロイドβプラーク形成の減少が見られた[130]。 同様に、アルツハイマー病の5xFADマウスモデルは、アミノ酸異化作用の変化とともに微生物叢集団の炎症誘発性種へのシフトを示し、逆に、抗生物質による治療はその効果を逆転させ、疾患の重症度と形質転換された腸集団との間の関連の可能性を示唆している[131]。 腸内で形成された微生物アミロイドタンパク質は、Toll-like receptor (TLR) を活性化し、分化クラスター 14 (CD14) を活性化して免疫応答を促進しアミロイドβ クリアランスの障害を伴う見過ごされたミスフォールドアミロイドβ につながり、続いてサイトカイン産生が増加し、腸と血液-脳関門が破壊される [114] . また、アルツハイマー病患者では腸内ホルモンのレベルが低下し、逆に硫化水素やトリメチルアミンなどの腸内微生物叢の代謝物が増加し、その厳しさが増すことが示されました[132]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 加齢に伴う腸内微生物の生物多様性の減少も、認知症の原因の 1 つと考えられています。 年齢が上がるにつれて、Bifidobacterium spp. が減少し、Proteobacteriaが増加することが示されています。認知症は短鎖脂肪酸の減少ではなく、脂質代謝の干渉によるものです。 Bifidobacterium spp.は、コレステロール値の調節において重要な役割を果たしており、その糞便排泄を直接促進し、血清レプチン値を増加させることで間接的に海馬の可塑性と記憶機能の維持に関与しています [133], [134]。 Lactobacilli や Bifidobacterium などの腸内細菌は、抑制性神経伝達物質γ-アミノ酪酸 を代謝します [135]。 ある研究では、アルツハイマー病の APPSwe/PSEN1DeltaE9 バイジェニック マウス モデルで、海馬のシナプス可塑性が変化していることが観察されました。 ここで、γ-アミノ酪酸産生の減少が、グルタミン酸作動性神経伝達の同時増加とともに見出された [136]、[137]。 ただし、cynobacteria門が認知障害を引き起こす神経毒を産生することが報告されていますが、アルツハイマー病との関係は観察されていません[138]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 別の可能性のある接続リンクである脳アミロイド蓄積と腸内微生物叢には、プリオン伝播と同様の方法で微生物アミロイドの交差播種のメカニズムが関与しており、したがって形成された異なるアミロイド配座異性体は、それらの細胞標的において異なるレベルで毒性を誘発し、アルツハイマー病表現型の存在を仮定している[ 139]。 腸内細菌叢に加えて、口腔内の共生細菌とアルツハイマー病との関連も研究されました。 興味深いことに、口腔衛生の悪さと歯の喪失は、アルツハイマー病の早期発症のリスクを高めることがわかった[140]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
5.4.パーキンソン病における腸内微生物叢媒介酸化ストレス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| パーキンソン病は、ドーパミン作動性ニューロンの進行性変性、リン酸化タンパク質 α-シヌクレインの凝集、過剰な活性酸素種産生、ミトコンドリア機能不全およびミクログリア活性化によって病理学的に特徴付けられる [141]。 症状的には、脳の黒質および線条体領域の損傷により、患者が随意運動(震え、筋肉の硬直、歩行困難、および猫背の姿勢)を制御できないことを特徴としています。 これは、世界中の高齢者人口の 1% 以上に影響を与える 2 番目に一般的な神経変性疾患です [142]。腸とパーキンソン病の関係に関する最初の報告は、1817 年に James Parkinson による震え麻痺に関するエッセイで言及されました [143]。 Braak の仮説は、腸で病状が始まり、その後脳に影響を与えるというこの見解を支持しました。 蓄積された証拠は、腸の炎症、迷走神経の背側運動ニューロンを超えるリン酸化α-シヌクレインの早期蓄積、便秘の問題、および腸透過性の増加がパーキンソン病患者で一般的であることを示しており、腸内微生物叢とパーキンソン病の病因との間の強い関係を示唆している[144] 。 さらに、迷走神経切除術を受けた個人ではパーキンソン病を発症するリスクが低下することが観察された [145]。 さらに、グルタチオンのレベルが低く、鉄分と過酸化水素水のレベルが高いと、黒質緻密部 (SNc) ニューロンが酸化ストレスを受けやすくなる [146]。 また、この領域での脂質過酸化とドーパミン酸化は神経細胞死につながります。 研究では、ミトコンドリア呼吸鎖の機能不全が過剰な活性酸素種産生につながることも示されています。 複合体1の阻害剤がドーパミンニューロンに対する細胞傷害効果を誘発するという事実[147]、またα-シヌクレイン、ホスファターゼおよびテンシンホモログ(PTEN)によって誘発される推定キナーゼ1(PINK1)およびパーキンの病状を有する患者が検出されたという事実によって裏付けられている。酸化ストレスの増加を伴うミトコンドリア機能不全を有する[148]。 アルツハイマー病と同様に、異常タンパク質α-シヌクレインの凝集は酸化ストレスの増加と相互に関連しており、逆もまた同様です。 腸内微生物叢の役割になると、いくつかの病原菌は腸の細胞にミトコンドリア機能不全を引き起こす毒素を放出し、腸神経系(ENS) は神経変性をもたらす [149]。 病原菌はパーキンソン病患者の腸内で増加し、その微生物産物はパーキンソン病の病因に直接関与しています。 この事実を裏付けるように、最近、病原菌である Escherichia coli がcurliとして知られるアミロイドタンパク質を産生することが報告されました。 腸と脳でα-シヌクレインの凝集を促進し、マウスの運動障害を引き起こすことが観察された[150]。 一方、マウスが腸制限アミロイド阻害剤で治療された場合、便秘の改善とともに運動機能の改善が示されており[151]、パーキンソン病症状の病因における腸の関与を示し、Braakの仮説を支持している。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内細菌によって引き起こされる腸の炎症は、パーキンソン病の進行性神経変性と直接相関する可能性があります。 パーキンソン病患者では血清代謝プロファイルと腸の組成が変化していることが判明しているが、重度のパーキンソン病状態では、腸内細菌科のレベルが非常に低いレベルの抗炎症性細菌が腸内で増加することが観察され、クローン病における腸の炎症との関連も示されている[ 152]。 これは、クローン病患者がパーキンソン病を発症するリスクが非常に高いことを示しています。 同様に、マウスモデル(PTEN誘導キナーゼ1(PINK1)ノックアウトマウス)におけるCitrobacter rodentium感染は、腸の炎症を誘発することによりパーキンソン病症状を悪化させることが観察された[153]。 炎症を誘発することに加えて、腸内微生物叢は、その代謝産物であるβ-グルコロネート、トリプトファン、および短鎖合脂肪酸がパーキンソン病患者の場合に変化することがわかった[154]などの代謝効果も発揮する。 最近、標準的なレボドパ治療の場合に観察されるように、腸内微生物叢の1つの顕著な属性も報告されており、バイオアベイラビリティを低下させるか、薬物の不活化を増加させることにより、抗パーキンソン病薬の有効性を低下させる能力を獲得することが示されている[155]、 [156]。 これらの研究を含めて、腸内細菌による水素産生の減少も、パーキンソン病の寄与因子の1つであることがわかっている[157]。 最近の報告によると、パーキンソン病のラットおよび MPTP マウスモデルに 50% 水素 飽和水を与えると、黒質および酸化ストレスマーカーの神経細胞損失の減少に成功することが判明し、二重盲検無作為化試験がヒトで実施された場合、パーキンソン病患者の運動評価の改善が観察された[157]、[158]。 さらに、この概念に向けて、ヒト α-シヌクレイン遺伝子を過剰発現させた パーキンソン病の無菌マウスモデルに関する最近の研究では、ミクログリア活性化の減少および運動機能の改善とともに短鎖脂肪酸レベルの低下が示されました。 それに加えて、パーキンソン病患者の腸内細菌叢を 無菌α-シヌクレイン過剰発現マウスモデルに移植すると、運動症状が悪化し、パーキンソン病患者における機能不全の腸内細菌叢の役割が示唆された [151]。 同様に、神経毒で処理されたマウスモデルは、病原性Enterobacteriaceaeのレベルの増加を伴う腸内微生物叢組成の変化を示した[159]。 さらに、殺虫剤ロテノンで処理したマウスでは、Fermicutes/Bacteroidesの比率の増加が観察された[151]。 驚くべきことに、Proteus mirabilis のようないくつかの特定の細菌種が、マウスの神経変性を促進することがわかった [160]。 まとめると、これらの研究は、パーキンソン病のヒトモデルと動物モデルの両方で、腸内微生物叢が神経機能障害と神経炎症を悪化させることを示唆しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
5.5.腸内微生物叢と外傷性脳損傷 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 外傷性脳損傷は、米国で年間約 140 万件発生しており、死亡および障害の主な原因の 1 つである [161]。 外傷性脳損傷で発生する障害には、脳の一次的な機械的損傷だけでなく、細胞および分子レベルで起こる損傷後の二次的損傷も含まれます。 また、ミトコンドリア機能障害、酸化ストレス、炎症、ミクログリア活性化、興奮毒性などの代謝異常を引き起こし、一時的または生涯にわたる認知障害を引き起こす可能性がある [162]。 さらに、外傷性脳損傷の重症度は脳だけに焦点を当てているだけでなく、多臓器損傷である可能性があり、異種の病態と見なされます。 脳損傷の不均一な性質のため、外傷性脳損傷誘発神経病理学の治療法はまだ不足しており、新しい治療計画の検討に関連しています。 このアプローチに向けて、腸内共生療法は、腸内細菌叢と外傷性脳損傷の間の両面関係を回復する能力があるため、多くの注目を集めています [163]、[164]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 増え続ける証拠は、腸内微生物叢と外傷性脳損傷損傷との間に双方向の関係が存在することを明らかにしています。 外傷性脳損傷の全身症状の 1 つは、腸の運動性と透過性の破壊、粘膜の損傷、腸絨毛の組織病理学的変化として観察されており、腸内微生物叢の組成の乱れを示している [165]、[166]、[167]。 逆に、腸内毒素症は、外傷性中枢神経系損傷の病態生理学、血液-脳関門透過性の変化、ミクログリアの活性化にも影響を与え、深刻な影響をもたらすことが観察されています [168]、[169]、[170]。 最近の報告では、軽度の反復的な脳損傷に20日間さらされたマウスは、白質損傷、認知機能の低下、および軽度の一過性の腸内毒素症の進行性の出現を示すことが示された[171]。 同様に、脳損傷前後の外傷性脳損傷のマウスモデルにおける腸内微生物叢の枯渇は、CA1海馬ニューロン密度の増加、連合学習障害の減弱、および病変体積の減少をもたらすことが観察された[172]。 最近の研究では、脊髄内の炎症と病変の病理を引き起こした外傷性脊髄損傷の後に腸内毒素症が起こると主張している [173]。 腸内細菌叢集団における同様の調節であるが、Firmicutes の減少と Bacteroidetes の増加の反比例の関係が損傷の 2 時間後に発生し、中程度の外傷性脳損傷のげっ歯類モデルで約 7 日間持続することがわかった [175]。 同様に、最近の報告では、マウスの外傷性脳損傷の 24 時間後に、Lactobacillus gasseri, Ruminococcus flavefaciens, および Eubacterium ventriosum の 3 種が急速に大幅に減少し、ヒトの腸内微生物叢でEubacterium sulci, と Marvinbryantia formatexigens が大幅に増加したことが示された [176]。 . さらに、72時間以内に Bacteroidales, Fusobacteriales, および Verrucomicrobialesの減少が、Clostridiales と Enterococcusの増加とともに、重度の外傷を負った多発性外傷患者で観察された[177]。 腸内細菌叢は、外傷性脳損傷からの回復においても非常に重要な役割を果たしています。 最近の報告で観察されているように、腸内微生物叢も外傷性脳損傷からの回復に非常に重要な役割を果たしています。 外傷性脳損傷の前、最中、後に広範囲の抗生物質によって誘発された腸内毒素症により、神経細胞の喪失が増加し、神経新生を抑制し、恐怖記憶反応の変調とともにミクログリアと末梢免疫反応を変化させた[178]。 したがって、外傷性脳損傷患者に対する腸内微生物叢の影響は、臨床的に最も重要です。なぜなら、外傷性脳損傷患者は、定期的な抗生物質の投与と入院の長期化により、腸内細菌叢の変化を受けやすくなるためです。 さらに、腸内微生物叢の変調の検出は、外傷性脳損傷の重症度を特定するための診断ツールを提供する可能性があり、したがって、標的治療アプローチを提供します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
6.中枢神経系の抗酸化物質と神経保護 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 中枢神経系は酸化ストレスに対して非常に脆弱であり、神経障害を引き起こします。 高レベルの活性酸素種は、高い酸素需要と過酸化感受性脂質細胞の急増により、中枢神経系の機能中に生成されます。 この酸化的代謝は、シナプス可塑性などの重要な機能を調節するための酸化還元シグナルを伝達するための反応種を生成します [29]、[179]。 酵素的または非酵素的、内因性または外因性の抗酸化物質は、活性酸素種の生成を防止するか、フリーラジカルを除去するか、フリーラジカル生成物を不活性化することにより、脳を酸化ストレスから保護します。 抗酸化防御メカニズムの第一線には、スーパーオキシドジスムターゼ、グルタチオンペルオキシダーゼ (GPx)、グルタチオンレダクターゼ、カタラーゼなどの内因性酵素の使用が含まれる [180]。 一方、抗酸化防御の第 2 段階では、チオレドキシン、フェリチン、トランスフェリン、セルロプラスミン、アルブミン、メタロチオネインなどの内因性の非酵素分子を使用します。 また、酵素補因子、すなわちコエンザイムQ と α-リポ酸、および代謝物、すなわちビリルビン、メラトニン、および尿酸は、抗酸化防御メカニズムにおいて重要な役割を果たしている [181]、[182]。 同様に、ビタミンA、E、C、フラボノイド、フェノール酸、カロテノイドなどの天然の食物化合物も、酸化ストレスによって誘発される神経変性に対する強力な抗酸化防御を有することが知られている[182]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| スーパーオキシドジスムターゼ などの抗酸化酵素は、スーパーオキシドの 酸素と 過酸化水素 への不均化を触媒することにより、スーパーオキシド アニオンの濃度を低下させます。 ルタチオンペルオキシダーゼは 過酸化水素と過酸化脂質を減らします。ペルオキシレドキシンなどのチオール特異的ペルオキシダーゼはヒドロキシペルオキシドの量を減らし、カタラーゼは過酸化水素を 水と通常の分子酸素に変換します [29]、[183]。 フリーラジカルは、抗酸化経路に関与する遺伝子の転写を活性化し、細胞を悪影響から保護します。 グルタチオンは中枢神経系に少量存在し、還元型のフリーラジカルと反応して過酸化水素を水に変換し、GS-SG に自己触媒作用を及ぼし、レダクターゼによって再びグルタチオンに再生する [183]。 グルタチオンが不足すると、ペルオキシダーゼの活性が制限され、ニューロンが酸化ストレスを受けやすくなる可能性があります。 これまでに実施された in vivo および in vitro 研究では、スーパーオキシドジスムターゼ およびカタラーゼが、アミロイドβ 誘発性のニューロン毒性に続くニューロンの生存を改善することが示された [184]、[185]、[186]、[187]。 同様に、チオレドキシンとチオレドキシンレダクターゼの両方が脳で広く発現しており、ハンティングトン病およびアルツハイマー病の酸化ストレスモデルに対して神経保護効果を発揮することが知られています [188]、[189]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 別の非常に重要な経路は、Kelch 様 ECH 関連タンパク質 1 核因子赤血球 2 関連因子 2 抗酸化応答要素 (Keap1-Nrf2-ARE) です。 Keap1-Nrf2-ARE はニューロンで高度に発現しており、神経変性疾患に関連する酸化ストレスに対する防御メカニズムと関連しています。 転写調節を介してスーパーオキシドジスムターゼ、チオレドキシン、ペルオキシレドキシン、およびグルタチオンの活性を調節する[190]。 NF-κB の重要性は、活性酸素種によって活性化される中枢神経系の酸化還元センサーとしても報告されています。 適度なレベルの活性酸素種は、NF-κB 活性化をもたらす NF-κB 阻害剤をリン酸化します。 活性化された NF-κB は、抗アポトーシスの発現を調節し、カスパーゼ依存性の細胞死経路を阻害します。 ただし、高レベルの活性酸素種は、NF-κBを不活性化することにより、NF-κBの結合を阻害します。 このメカニズムは、細胞のアポトーシスを促進し、生存促進経路を停止させます [29]。 腸内微生物の役割が非常に動的である場合、中枢神経系の抗酸化代謝の調節は厳密に制御されています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
7.神経保護における腸内微生物叢 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
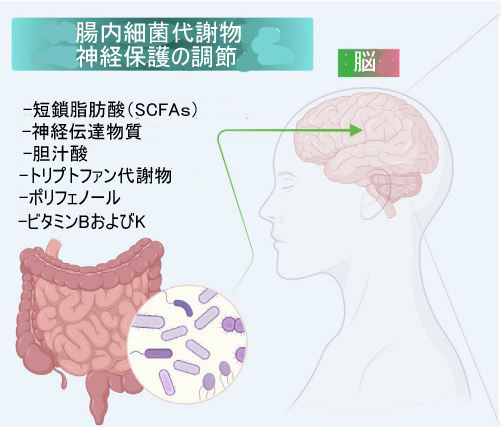
| 腸内微生物 - 微生物および微生物 - 宿主の相互作用は、吸収性ビタミン、ポリフェノール、短鎖脂肪酸、脳由来神経栄養因子 、拡散性抗酸化物質および酸化ガスなどのさまざまな代謝産物の生成を介して、外因性および内因性の活性酸素種のレベルを調節します。 腸内微生物はまた、血液-脳関門への代謝産物の透過性、タイトジャンクションの完全性、および腸のバリアを制御し、免疫系を調節し、病原体の腸内コロニー形成を妨げる[191]。 副交感神経系の迷走神経は、腸の代謝産物を感知し、腸の情報について中枢神経系に伝達して、特定の反応を生成します。 ストレス条件下では、迷走神経緊張が抑制され、過敏性腸症候群 (IBS) や腸内細菌叢による炎症性腸疾患 (IBD) などの有害な影響を示す [192]。 アルツハイマー病の病因に関与するアミロイドβタンパク質は、腸神経系の大腸菌やサルモネラ菌などの腸内細菌によって発現される[183]。 有益な腸内微生物は、ドーパミン、セロトニン、および γ-アミノ酪酸も生成します。 これらは、腸神経系活動を調節し、相関する可能性がある中枢神経系神経伝達物質である [179]。 いくつかの研究では、腸内微生物がミクログリアの活性化と成熟を管理し、活性化されたミクログリアが大量の誘導型一酸化窒素合成酵素 (iNOS) を放出して一酸化窒素産生を調節することが明らかになりました。 腸内毒素症は炎症性一酸化窒素合成酵素を誘発し、神経炎症を引き起こす [183]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
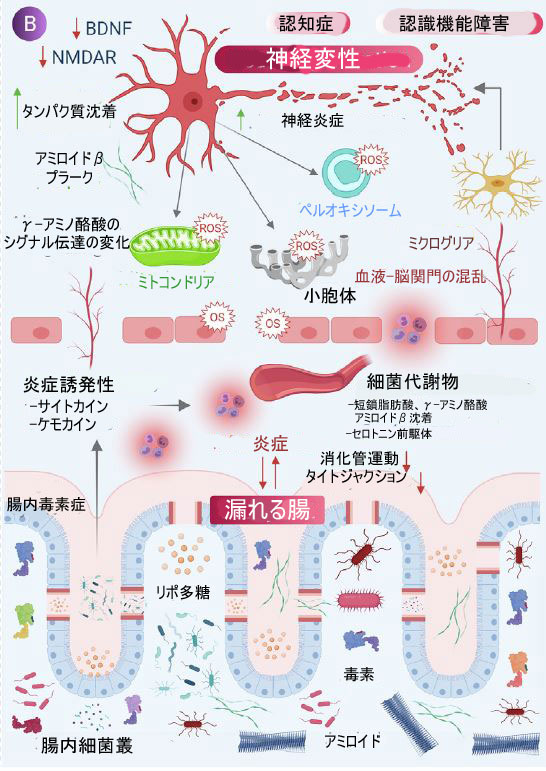
| 腸内微生物叢の潜在的な役割と、神経障害の媒介における酸化ストレスの重要な役割を見てきたように、近年、酸化ストレスによって誘発される神経変性疾患を治療するための抗酸化物質ベースの治療法を開発する必要性と主な科学的焦点が高まっています。 抗酸化物質は、ROS/RNS によって誘発される酸化ストレスに対抗できる化学物質または天然物質である [193]。 抗酸化物質の強力な治療効果は、糖尿病、関節炎、白内障、骨粗鬆症などの疾患で観察されていますが、中枢神経系障害に使用される抗酸化療法は限られており、依然として深いメカニズムの理解が必要です [194]。 抗酸化療法とは対照的に、腸内微生物叢の二分的な役割が観察されており、一方は神経変性の基本的な根底にあるメカニズム(腸内毒素症と神経炎症)に関与しています。 一方、腸内微生物叢とその代謝物は、神経保護の治療的役割を示唆する多くの関連経路を調節します。 タンパク質やビタミンなどの微生物分子は、多段階の生合成経路を介して生成され、宿主系に有益または有害な影響を与える可能性があります [16]。 したがって、プレバイオティクスとプロバイオティクスを含む適切な食事で維持される健康な腸内微生物叢は、神経細胞の健康を維持するための前提条件です [195]。 図 2 は、腸内細菌叢の代謝産物の役割と、腸内細菌叢の調節手順および 神経変性疾患への影響を示しています。 ここでは、細菌の代謝物 (自然代謝物、変換された代謝物、食事による代謝物) が、酸化ストレスによって誘発されるニューロンの損傷と戦うのにどのように役立つかを確認します。 さらに、神経保護における抗酸化および抗炎症プロバイオティクス/プレバイオティクスの役割についてさらに議論します. 表 1 は、神経保護の役割を示す腸内微生物叢に関する最近の研究を示しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
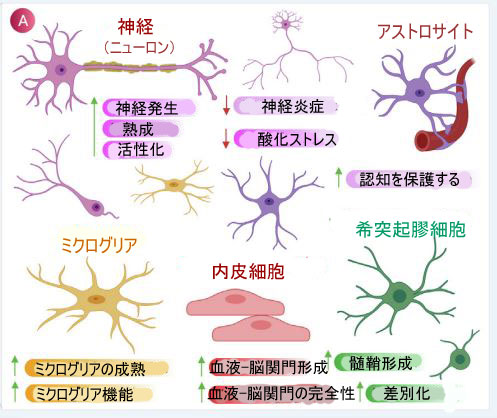
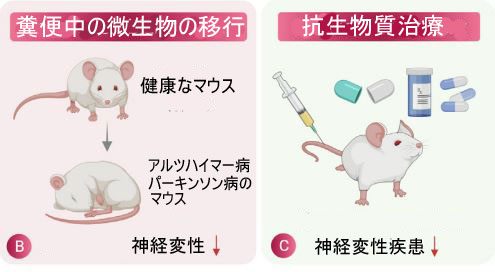
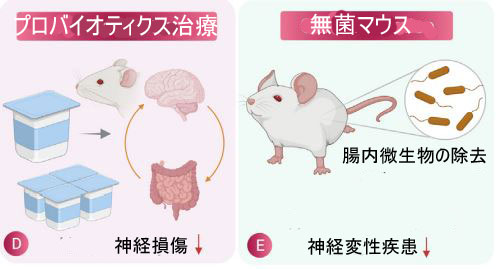
| 図2 神経保護における腸内微生物叢の役割 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (A) 炎症および酸化ストレスの軽減において、腸内微生物叢によって放出される有益な代謝産物に応答した脳細胞特異的効果のスキーム。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (B) 糞便中の微生物の移動 (FMT) は、健康な個人から病的な状態にある個人への糞便細菌の移動を含み、アルツハイマー病 (AD)、パーキンソン病 ( PD)、ハンチントン病 (HD)、および多発性硬化症 (MS) などの状態。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (C) および (D) 抗生物質治療とプロバイオティクス治療はそれぞれ、神経変性疾患の病因を減少させる上でかなりの有効性を示しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (E) 無菌マウス (腸内細菌叢のないマウス) は、神経変性状態の減少を示し、腸内細菌叢が脳の生理機能に及ぼす影響を研究するためにも使用され、神経変性における腸内細菌叢の関与を示しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
8.細胞特異的応答による神経保護における腸内微生物叢代謝産物 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
8.1宿主分子との腸内微生物叢の相互作用 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
8.1.1.胆汁酸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 胆汁酸は肝臓で生成され、腸管腔で放出され、主に脂質と脂溶性ビタミンの可溶化に関与し、エネルギー代謝のシグナル伝達に関与し、脳の生理学と病態生理学において重要な役割を果たすことも知られている [196] . 胆汁酸は、血液-脳関門を通過する脳内の受容体に直接結合するか、腸受容体に結合して線維芽細胞増殖因子 (FGF) およびグルカゴン様ペプチド 1 の放出を間接的に誘導することにより、さまざまな脳領域のニューロン活動および迷走神経ニューロン活動に影響を与えます [ 197]。 ウルソデオキシコール酸 (UDCA) やタウロウルソデオキシコール酸 (TUDCA) などの胆汁酸は、それぞれ第 III 相臨床試験 [198] と動物実験 [199] で証明されているように、神経保護特性を持っており細胞毒性がありません。 最近のデータは、パーキンソン病の慢性マウスモデルにおいて、タウロウルソデオキシコール酸がオートファジー、α-シヌクレイン凝集、およびタンパク質酸化を軽減するのに役立つことを示している[200]。 さらに、武田薬品の G タンパク質共役受容体 5/サーチュイン-3 (TGR5/SIRT-3) 経路を介して、くも膜下出血の影響を受けたラットのニューロンのアポトーシスを防ぐのに役立ちます [201]。 同様に、ウルソデオキシコール酸は前頭側頭型認知症の荷電多胞体タンパク質 2B (CHMP2B) イントロン 5 モデルで神経保護効果を示した [202]。 腸管腔内の腸内微生物叢は、デヒドラターゼの作用によって一次胆汁酸(コール酸とケノデオキシコール酸)を二次胆汁酸に変換し、胆汁酸塩加水分解酵素によるアミノ酸の脱抱合および他の酵素プロセスを伴います。 これにより、核内受容体の結合、溶解度、および循環が変化します [203]。 二次胆汁酸レベルの変化は、アルツハイマー病[204]、パーキンソン病[205]、自閉症スペクトラム障害[206]、および多発性硬化症[207]のヒトおよびマウスモデルで発見されています。また、細菌で修飾された胆汁酸は、筋萎縮性側索硬化症や脳卒中の場合に神経を保護することがわかった [208]。 腸内微生物群集の調節は、胆汁酸のレベルと特性の変化を引き起こす可能性があり、これは神経変性または神経保護的である可能性があります。 腸内細菌叢を介したデオキシコール酸の増加が、マウスの腸クロム親和性細胞で神経伝達物質セロトニンの放出を誘導することも報告されている [94]。 胆汁酸代謝産物は、脱髄 [209] を改善し、酸化ストレスを軽減 [210] し、オリゴデンドロサイトとミクログリアにそれぞれ作用することで神経保護の役割を強化することがわかった。 それでも、微生物叢が操作した胆汁酸の潜在的な役割と効果は不明であり、明確に定義されていません。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
8.1.2.ステロイドホルモン | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ステロイド ホルモンによるシグナル伝達は、脳の発達と機能 (記憶、意思決定、性行動) にとって重要です [211]。 体全体を循環している間、ステロイドホルモンは腸内腔で微生物叢に出会う[212]。 腸内細菌は、β-グルクロニダーゼとβ-グルコシダーゼの作用によって媒介される脱抱合反応でステロイドホルモンを修飾し、ホルモンを再活性化して排泄を防ぎます。 したがって、腸内微生物叢は、分解および活性化経路を介して活性および不活性ステロイドホルモンのレベルに影響を与える [213]。 アンドロゲンとエストロゲンは、腸内微生物叢の影響を受けることがわかっています。 多数の腸内細菌がエストロゲンを代謝することがわかっており [214]、エストロゲンは糞便サンプルでも酸化還元反応を起こし [215]、腸内微生物叢の役割を示唆している。 腸内微生物叢には、テストステロン [216] とコレステロールをアンドロゲン [217] に変換する能力もあることが観察されています。 微生物の影響を受けたエストロゲンは神経保護的であることが判明し、ミクログリアに対する抗炎症効果を示している[218]。 また、腸内微生物群集の変化がエストロゲンレベルの低下をもたらすことが観察され、慢性炎症[219]および認知障害[220]をもたらした。 エストロゲン分子は、オリゴデンドロ サイトの分化と髄鞘形成にも影響を与える [221]。 最近の報告では、MPTP パーキンソン病マウスモデルにおけるプロゲステロン治療でさえ、神経保護、抗炎症、および免疫調節効果を示したが、神経保護の役割が腸または脳で始まるかどうかはまだ不明であることが示されている [222]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
8.2.食物分子との腸内微生物叢の相互作用 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
8.2.1アミノ酸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 食事性アミノ酸は腸内微生物によって代謝されることもあり、脳への影響は食事摂取の種類と頻度によって異なる [223]。 ただし、腸内微生物叢がコードするアミノ酸は宿主内を循環するが、中枢神経系に影響を与えるのは腸内細菌によって代謝される食事性アミノ酸である [224]。 チロシン、トリプトファン、フェニルアラニンなどの芳香族アミノ酸は、腸内細菌によって短鎖脂肪酸、インドール誘導体、神経伝達物質、有機酸、アミン、アンモニアに代謝される [225]。 チロシン代謝の最終産物は、チラミン中間体からの 2 つのカテコールアミン、ドーパミンおよびノルアドレナリンの形成です。 In vitroの研究では、多数の腸内細菌がミリモル範囲でノルアドレナリンを産生できることが示された [226]。 最近の報告によると、非アドレナリンは B-3 アドレナリン受容体を刺激することによって星状細胞からのグルタチオンの供給を増加させ、過酸化水素によって誘発されるニューロン死からニューロンを保護することが示された [227]。 チロシンはまた、腸内微生物叢によって 4-エチル フェノールのようなフェノールで代謝され、その後宿主内で硫酸化されて 4-エチル フェノール サルフェートになることがわかっています。 これは、自閉症スペクトラム障害のマウスモデルで上昇していることが判明しており、自閉症スペクトラム障害の子供の尿バイオマーカーでもある[228]。 トリプタミンやキヌレニンなどのインドール誘導体は、神経活性分子である腸内細菌によるトリプトファン代謝の産物である [92]。 インドール誘導体であるインドールプロパン酸は、神経炎症を軽減する抗酸化物質として作用し、アルツハイマー病の病理を減少させる潜在的な役割を果たしていることが観察されている[229]。 キヌレニン代謝物は、不安、記憶、ストレス様行動に影響を与えることがわかった[230]。 キヌレニン代謝経路の障害が炎症、興奮毒性のあるグルタミン酸産生、およびフリーラジカル攻撃を促進することも観察されており、バランスのとれたキヌレニンの神経保護的役割と、アルツハイマー病、パーキンソン病、および ハッチントン病 におけるその抗炎症的役割を示唆している [231]。 トリプトファン代謝産物は、アストロ サイトのアリール炭化水素受容体を調節することによってアストロ サイトの炎症反応を減少させ、ミクログリアとの相互作用にも影響を与えます。 同様に、インドキシル-3-硫酸がミクログリアの活性化を制御し、その後アストロサイトとの相互作用を制御することも観察されている[232]。 アミノ酸グルタミン酸は、腸内細菌のグルタミン酸脱炭酸酵素によっても変換され、抑制性神経伝達物質であるγ-アミノ酪酸を形成します。γ-アミノ酪酸は、マウスモデルでうつ病や不安症状を軽減することが観察されています [90]。 アミノ酸アルギニンの場合、グルタミン酸受容体を介して作用し、シナプス可塑性と記憶形成の維持に関与する 4 つのポリアミンであるアグマチン、プトレシン、スペルミジン、スペルミンに代謝される [233]。 アグマチンは、脳内のα-2アドレジェンおよびイミダゾール受容体のリガンドとして作用しながら、CNS障害の場合に治療効果を示します[234]。 スペルミジンは、ハッチントン病の 3-ニトロプロピオン酸 3-NP モデルでも神経保護効果を示した [235]。 さらに、アグマチンは Nrf-2 シグナル伝達経路を刺激し、リポ多糖 (LPS) によって誘導される 活性酸素種 産生を改善することがわかった [236]。 in vitro および in vivo の研究では、アグマチンが星状細胞とミクログリアを酸化ストレスによって誘発される損傷から保護することも明らかになった [237]。 これらの研究は、神経科学における腸内微生物内分泌学の潜在的な役割を示唆しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
8.2.2.食物繊維 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 複合炭水化物多糖類のような未消化の食物繊維は、腸内微生物酵素グリコシド加水分解酵素および多糖類リアーゼによって作用され、嫌気性発酵によって短鎖脂肪酸に変換される [238]。 酪酸、酢酸、およびプロピオン酸は、結腸上皮細胞のエネルギー源として機能する短鎖脂肪酸を構成します。 これに加えて、それは体循環にも入り、その後神経発達や機能を含む多くの臓器の生理学的機能に直接的または間接的に影響を与える [238]。 報告によると、アルツハイマー病マウスモデルの腸内細菌叢が、プロバイオティクスの混合物で処理するか、抗短鎖脂肪酸を使用することによって調節されると、疾患の進行に対抗するのに役立つことが明らかにされている [239]。 同様に、短鎖脂肪酸は、パーキンソン病の無菌マウスモデルで運動症状を悪化させるのに効果的であることがわかった[240]。 短鎖脂肪酸、酢酸塩は 血液-脳関門を通過し、ニューロンを活性化し、神経伝達物質と神経栄養因子のレベルを調節する [241]。 ある研究では、プロピオン酸と酪酸がニューロンの細胞内カリウムレベルに影響を与えることが示された [242]。 酪酸は、エピジェネティックな遺伝子活性化 (ヒストン脱アセチル化酵素) を調節する酵素の強力な阻害剤として作用し、アルツハイマー病、パーキンソン病、ハッチントン病、脳卒中および記憶障害のマウスモデルで強力な抗炎症剤として作用することがわかっている [238]。 興味深いことに、短鎖脂肪酸は アミロイドβ ペプチド間の相互作用を妨害して神経毒性オリゴマーを形成し、アルツハイマー病を予防する [243]。 また、野生型マウスからパーキンソン病の動物モデルへの糞便微生物叢の移植と、酪酸の投与が、運動症状とドーパミン欠乏症を著しく改善したことも報告されている[244]。 細胞特異的な応答を見ると、酪酸は in vitro で星状細胞の神経炎症と酸化を減少させることが示されており、酢酸はこれらの細胞によってエネルギー源として使用されている [245]。 ある研究では、短鎖脂肪酸が内皮細胞上の短鎖脂肪酸受容体に結合することによりタイトジャンクションタンパク質の発現を増加させ、血液-脳関門の透過性を低下させ、リポ多糖誘発性の発作と脳卒中を予防するのに役立つことが観察された[246]。 短鎖脂肪酸は、ミクログリアの活性化を減少させることによって脳内の酸化ストレスを軽減し[247]、アルツハイマー病およびパーキンソン病の神経炎症と戦う。 まとめると、食物繊維から得られる短鎖脂肪酸は、個人の腸の健康に応じて、脳の健康を改善するのに役立ちます。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
8.2.3.ポリフェノール | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ポリフェノールは、植物に存在する生理活性分子であり、植物の成長、保護、および繁殖において基本的な役割を果たします [248]。 ポリフェノールは、フラボノイド、フェノール酸、タンニンに分類されます。 ポリフェノールの分子構造、すなわちヒドロキシル基の位置と芳香環の置換の性質により、フリーラジカルを除去する能力が得られ [248]、神経変性疾患の治療のための抗酸化療法として広く研究されている [249]。 吸収されなかったポリフェノールは、加水分解とエステル化を介して腸内微生物叢の作用によって宿主内で生物学的に利用可能な生物活性代謝物に変換され[250]、さらに末梢組織に到達する前にメチル化、硫酸化、ヒドロキシル化などの修飾が行われる[251]。 ブドウやワインから得られるレスベラトロール、クルクミン、緑茶からのエピガロカテキン-3-ガレートなどのさまざまなポリフェノールに関する研究は、内因性および外因性の両方の活性酸素種を緩和する主要な経路である Keap1/Nrf-2/ARE などのプロテインキナーゼ経路を活性化することにより、神経保護効果を発揮します。 . また、3-ヒドロキシ安息香酸や 3-(3'-ヒドロキシフェニル)プロピオン酸などの細菌のポリフェノール代謝産物がアミロイド凝集を阻害し、アルツハイマー病の進行の阻害を促進することも報告されている [252]。 同様に、フラボノイドのケルセチンが BACE-1 阻害剤(訳者注:BACE1(β-site Amyloid precursor protein Cleaving Enzyme 1,β-セクレターゼ)は,アミロイドβ前駆タンパク質(APP)を切断する酵素の一つで,この反応によりアルツハイマー病に特徴的な老人斑の主要構成成分であるアミロイドβタンパク質(Aβ)を生じます。老人斑形成を抑制するアルツハイマー病治療薬候補としてBACE1阻害物質の研究が盛んに行われています。)として作用することが観察されている [253]。 さらに、天然のフラボノイドプロアントシアニジンは、インビトロでドーパミン作動性ニューロンのロテノン誘発酸化ストレスを緩和する[254]。 また、腸内微生物叢によって産生されるフェルラ酸のようなポリフェノールは、神経新生を促進し、アルツハイマー病[255] および脳虚血 [256] のマウスモデルで神経保護の役割を示しています。 エクオールとエンテロラクトンも、ポリフェノールの 1 つである植物エストロゲンを代謝することによって腸内細菌によって生成される誘導体であり [257]、エストロゲン受容体によって媒介される古典的な神経保護経路に影響を与える可能性がある。 興味深いことに、ポリフェノールは腸内微生物叢の組成を調節することもでき [258]、さらにそれらを抗炎症および神経保護代謝産物に変換します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
9.ビタミンBとビタミンK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内微生物叢はビタミン、特にビタミン B と K の重要な供給源としても機能し、ビタミン B と K は腸内微生物の代謝に不可欠であるだけでなく、宿主の生理学的経路にも影響を与えることが示されている [259]。 Escherichia coli、Klebsiella pneumoniae、Propionibacterium、Eubacterium などの腸内細菌は、ビタミン K を生成します。 B2 (リボフラビン) は、枯草菌と大腸菌によって生成され、B9(葉酸)はBifidobacterium, Lactococcus lactis およびStreptococcus thermophilusによって生成され、B12 (コバラミン) は、Lactobacillus reuteri および Propionibacterium freudenreichi によって生成される [260]。 食事からのビタミンは小腸から吸収されますが、微生物叢由来のビタミンの取り込みは結腸で行われます。 新生児の出血性疾患を予防するために、腸内微生物叢が確立される前に、ビタミン K が血栓症のプロセスに不可欠な薬剤として投与される [261]。 さらに、ビタミン B と K は脳の発達と機能に重要であることがわかっている [262]。 神経変性疾患の場合、神経細胞の健康を改善する上でビタミン B と K が効果的な役割を果たしていることが多くの研究で示されています。 ビタミン K 欠乏症は、アルツハイマー病の病因と相関していることが判明しており、同様に、食事によるビタミン K の摂取量の増加は、高齢患者の記憶機能の改善に役立っています。 最近の報告では、ビタミン K2 (メナキオン-4) が強力な抗酸化特性を持っていることが示されています。 ロテノン誘導p38活性化、活性酸素種産生およびカスパーゼ-1活性を有意に阻害し、続いてミトコンドリア膜電位を回復させることが判明し、神経炎症誘導パーキンソン病の治療におけるその可能性を示している[263]。 同様に、ビタミンK2は、PC12マウス神経芽細胞腫細胞のbaxおよびカスパーゼ-3活性化を調節し、6-OHDA誘導アポトーシスから保護するのに有効であることが判明した[264]。 特に、ある最近の報告では、パーキンソン病患者における低レベルのビタミンK2が調節不全の炎症反応およびカスケード凝固シグナルと関連していることが示されている[265]。 ビタミン B の欠乏は、脚気や多発性ニューロパシーなどの神経障害と関連することも判明した [266]。 同様に、葉酸欠乏症は、年配の女性の認知障害および進行性認知症と関連していることが判明した [267]。 また、B6、B9、および B12 の高摂取は、アルツハイマー病の認知機能低下に関連する脳領域の萎縮率を低下させる [268]。 さらに、神経保護における腸内微生物叢産生ビタミンの潜在的な関連性を証明するには、より建設的な研究が必要です。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
10.プロバイオティクスの抗酸化作用と抗炎症作用 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 活性酸素種の形のフリーラジカルは、通常の細胞代謝プロセスの副産物です。 フリーラジカルは、遺伝物質の損傷、酵素の不活性化、複雑な炭水化物の解重合、および脂質過酸化を引き起こす可能性があります。 この細胞内分子の破壊は、細胞死につながります。 フリーラジカルのバランスをとるために、体にはグルタチオンなどの特定の抗酸化物質があります[269]。 また、抗酸化作用と抗炎症作用を持つフェルラ酸 (FA) などの酸化ストレスに対処するために、いくつかの製品が医薬品市場に流れ始めました。 脳由来神経栄養因子(BDNF)、神経成長因子(NGF)、および抗炎症特性を持ついくつかの神経ペプチドの産生を強化することにより、神経幹細胞の増殖を助けます[270]、[271]。 2017年、Westfallらは、フェルラ酸はLactobacillus plantarum NCIMB 8826、Lactobacillus fermentum NCIMB 5221、Bifidum animalisなどのプロバイオティクス細菌によっても、細菌のフェルラ酸エステラーゼ酵素によって大量に生成されることを報告した[272]。 その治療効果により、アルツハイマー病治療での使用が注目されており、フェルラ酸による前処理は アミロイドβ 線維を減少させ、アルツハイマー病マウスの神経炎症を治療することが示されている [273]。 フェルラ酸産生プロバイオティック細菌は、活性酸素種を除去することにより、βアミロイド線維の形成と凝集を阻害します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 他のプロバイオティクス細菌タンパク質、サーチュイン-1 (SIRT1) タンパク質デアセチラーゼは、抗酸化特性を持つことが示されています。 このタンパク質は、宿主の抗酸化経路の遺伝子を調節し、神経保護効果があることが実証されています。 最近の研究では、SLAB51 (Streptococcus thermophilus、Lactobacillus acidophilus、L. plantarum、L. paracasei、L. delbrueckii subsp. bulgaricus、L. brevis、Bifidobacterium longum、B. breve、B. Infantis) という名前のプロバイオティクス細菌の製剤の能力が調査されました。酸化ストレスを軽減し、その効果の分子メカニズムを発見した [274]。 処理されたトランスジェニック 3xTg-アルツハイマー病-マウスでは、脳内のサーチュイン-1タンパク質の発現と活性が回復することが観察され、アミロイドβペプチドの形成が減少したことが示されました。 しかし、未治療の アルツハイマー病マウスでは、サーチュイン-1の発現が大幅に減少していることがわかりました。 さらに、サーチュイン-1の強化された活性は、p53タンパク質のアセチル化を減少させ、アポトーシス経路を抑制することにより、ストレスを受けた細胞の生存を改善します。 他の研究では、プロバイオティック サプリメントが サーチュイン-1 経路を活性化し、抗酸化効果を誘発することも示されました。 SLAB51 はまた、グルタチオンおよびカタラーゼ抗酸化酵素の活性を高め、酸化ストレスを引き起こす障害を軽減します。 同様の発見がヒトでも観察されました。 アルツハイマー病患者の脳内のSIRT1濃度は有意に減少していることが判明し、これはアルツハイマー病患者の大脳皮質におけるアミロイドβおよびタウの蓄積と密接に関連している[275]。 プロバイオティクス株であるLactobacillus casei 01を補充したヒト研究で、SIRT1レベルの改善が報告されている[276]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内微生物叢と宿主免疫細胞の間には非常に壊れやすい関係があります。 免疫細胞は、宿主に優しい細菌と病原性細菌を特異的に区別するように特殊化されています。 この関係が損なわれると、不必要な免疫反応が引き起こされ、慢性炎症が引き起こされる可能性があります [277]。 この分化は、リポ多糖、ペプチドグリカン、フラジェリンなどの細菌細胞表面抗原に基づいて教育されたマクロファージ表現型を生成する役割を担う腸上皮細胞によって開始されます。 上皮細胞が病原体の攻撃を受けやすくなると、抗原は血管系に移行し、それに応答して、IL (インターロイキン)-1、IL-6および腫瘍壊死因子アルファ (TNF-α) が産生され、腸と脳に敗血症性ショックと炎症を引き起こします。 一部の細菌毒素は血液-脳関門を通過することもできる [269]、[278]。 アミロイドーシスにおける微生物叢の役割を調べるために、プロ (CXCL2、CXCL10、IL-1β、IL-6、IL-18、IL-8、インフラマソーム複合体 NLRP3、TNF-α) およびアンチ - 認知障害のある患者におけるいくつかの腸内微生物叢の炎症性 (IL-4、IL-10、IL-13) サイトカイン活性が観察されました。 Escherichia/Shigella の数が増加し、Eubacterium rectale が減少することは、認知障害のある患者およびアミロイド陽性の患者における炎症誘発性サイトカインおよび抗炎症性サイトカインの濃度の変化と有意に相関していました。同時に、IL-6、CXCL2、NLRP3、および IL-1β のレベルの増加と IL-10 のレベルの減少が アルツハイマー病患者で観察されました。 この研究は、腸内微生物叢が神経疾患の末梢炎症を開始、悪化、または緩和する可能性があることを示唆している[279]。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
11.腸脳研究を進めるための in silico 戦略 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| テクノロジーの進歩に伴い、脳に関する知識は指数関数的に増加しています。 神経科学者は、分子生物学、分子遺伝学、脳画像、およびその他の新しい技術から多くのデータを生成しており、さまざまな分析のために神経科学データを共有することに大きな関心を持っています。 ニューロイメージングは、神経変性疾患や精神障害の予測と検出にも非常に役立ちます。 神経科学データの収集と分析のために、脳機能に基づいたデバイスをさらに開発するために、さまざまなバイオインフォマティクス ツールが開発されています。 今日まで、研究者は、さまざまなストレス状態による神経変性に関与する分子経路について基本的な知識を持っています。 統合されたデータベースは、公開されている情報源から神経変性疾患に関連するさまざまな種類のデータを収集するのに役立ちます (表 2)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| マイクロバイオームの包括的な知識と人間の健康におけるそれらの重要性は、ハイスループット技術の強化に伴い急速に増加しています。 微生物叢は宿主の代謝産物と相互作用し、それらを二次代謝産物に変換します。 腸内微生物叢は、人間の健康に非常に重要な役割を果たす代謝物も生成します。 代謝産物と腸内微生物叢の相互作用を理解することで、健康と病気の状態における微生物叢の機能を知り、自然療法を開発することが重要である [280]。 最近では、腸内細菌が中枢神経系の生理機能を変化させる可能性があり、腸内細菌叢が神経炎症の潜在的な要因である可能性があることが証明されている[281]。 2019 年、Lu と Claud は、初期の定着と微生物叢の発達が早産児の脳の発達に影響を与えることを報告した [282]。 この指数関数的に増加する情報を管理および促進するために、人間の健康における微生物叢の役割を深く理解するために、微生物叢データベースが開発されています。 微生物叢と代謝物の相関関係を確立するために、自動化されたマイクロバイオームとメタボロームの統合分析パイプラインである M2IA などの新しいパイプラインが開発されている [283]。 他の微生物叢データベースは、疾患に関連する微生物、微生物叢による薬物の調節、腸内微生物に対する食事の影響に関する有用な情報を提供します。 微生物叢による治療薬、食事成分、ハーブ製品、プロバイオティクス、環境化学物質などの活性物質の変化、および活性物質による微生物叢の変化に関する知識を広めるために、Microbiota-Active Substance Interactions (MASI) データベースが開発されている [284]。 これらのデータベースは、細菌の代謝産物と神経細胞の健康および病因との相関関係を確立するのに非常に役立ちます。臨床試験では、パーキンソン病に対するL-ドーパ治療の結果は、募集対象によって異なることが報告されています。 この変動は、微生物叢によるものです。 L-ドーパは、異なる腸内細菌種、すなわちEnterococcus faecalisおよびEggerthella lenta A2のチロシンデカルボキシラーゼ(TDC)およびドーパミンデヒドロキシラーゼ(Dadh)によって代謝される[285]。 細菌の L-ドーパ脱炭酸酵素を不活性化するための薬剤、(S)-α-フルオロメチルチロシン (AFMT) が発見されました。 L-dopa と AFMT の組み合わせは、パーキンソン病の治療に使用されています。 Zhuangらは、Bacteroides, Actinobacteria, Ruminococcus, Lachnospiraceae, and Selenomonadalesなどの分類学的レベルで、アルツハイマー病患者の腸内微生物叢が健康な個人と比較して変化していることを示した[286]。 メタゲノム データ/16S RNA シーケンス データは、腸内細菌異常症とアルツハイマー病の間の確固たる関連性をサポートしています。 リポ多糖や短鎖脂肪酸などの細菌産物は、アミロイド病理と関連している [287]。 これらのデータベースの開発の主な目的は、神経変性に関連するさまざまな未解決の謎に対する解決策を提供するために、アナリストによるさまざまな分析のための情報を取得するのに役立つ一連の構造化レコードにデータを整理することです。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
12.結論 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内微生物叢と脳の相互接続は、神経科学研究に革新的な進歩をもたらしました。 今日までに発見された研究のほとんどは腸内細菌の神経科学研究に関連していますが、腸内微生物叢は広大であり、腸内微生物とその群集の役割を特定して特徴付け、腸と脳のシグナル伝達経路に影響を与えるために科学者の注意が必要です。 神経変性と神経保護の過程で腸と脳の間のコミュニケーションを仲介する際に腸内微生物叢が使用する経路をさらに理解する必要があり、病理学と病気の治療における新しい仮説への道を開く可能性があります。 糞便中の微生物の移動やプロバイオティクスなどの新しい治療レジメンは、脳の健康をある程度改善するのに役立ちましたが、市販のプロバイオティクス株に経験的な証拠が欠けているため、特定の神経変性疾患の治療のために特定のプロバイオティクスを研究する必要性が生じています。 さらに、インシリコ戦略の可能性が高まることで、神経変性と神経保護における微生物エフェクター分子を深く理解するのに役立ちますが、残念ながら、これらの研究は非常に限られています。 バイオインフォマティクス ツールは時代のニーズです。これは、有害な代謝産物の産生を仲介する特定の細菌酵素を標的とすることができるような薬物の設計を強化する可能性があります。腸内微生物代謝産物の神経保護および神経変性の役割は引き続き明らかにされており、新しい治療法を開発する機会を提供しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
この文献は、Journal of Advanced Research 38 (2022) 223–244に掲載されたInterplay of gut microbiota and oxidative stress: Perspective on neurodegeneration and neuroprotection.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||