ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(アロニア:研究の最前線) |
ラット網膜におけるヨウ素酸ナトリウム誘発損傷からの |
Yan Xing et al., |
|
|
| 概要 |
加齢黄斑変性症 (AMD) は、高齢者の失明の主な原因の 1 つです。 ただし、乾燥型の加齢黄斑変性症には有効な治療法がありません。 アロニア メラノカルパの果実にはアントシアニンが豊富に含まれています。 この研究では、ラット網膜に対するアロニア果実抽出物の保護効果を、ヨウ素酸ナトリウム誘発乾性加齢黄斑変性症モデルを使用して調査しました。 フルフィールド網膜電図 (ERG) は、b 波の振幅が大幅に減少し、網膜構造がモデルで乱れていることを示しました。 抽出物治療により、損傷が軽減されました。 b 波の振幅は、Scotopic 0.01ERG で 61.5%、Photopic 3.0ERG で 122.0%、Photopic 3.0 フリッカーで 106.8% 増加しました。 網膜構造障害は、外核層の厚さが44.1%増加して改善されました。 マロンアルデヒドのレベルは、モデルと比較して、抽出物で処理されたラットの網膜で大幅に減少しました。 プロテオミクス解析により、α-クリスタリンA鎖、β-クリスタリンB2、β-クリスタリンA3、α-クリスタリンB鎖、γ-クリスタリンSの5つのクリスタリンタンパク質の発現が明らかになりました。 これは網膜神経節細胞を保護し、免疫ブロッティングによっても確認された対照と比較して、抽出物処理でそれぞれ7.38倍、7.74倍、15.30倍、4.86倍、および9.14倍増加しました。 この結果は、おそらくそのアントシアニンに起因するアロニア果実抽出物が、酸化的損傷を軽減し、クリスタリンタンパク質をアップレギュレートして神経系を保護することにより、ラットの網膜を保護できることを示唆しています。 |
| 目次(クリックして記事にアクセスできます) |
| 1.はじめに |
| 2.材料と方法 |
| 2.1. 動物 |
| 2.2. アロニア果実エキス |
| 2.3. ラットにおけるヨウ素酸ナトリウム誘発網膜損傷に対するアントシアニジンの保護効果の評価 |
| 2.4. 網膜電図 |
| 2.5. 網膜のヘマトキシリンおよびエオシン (H&E) 染色 |
| 2.6. マロンジアルデヒド(MDA)の測定 |
| 2.7. 網膜タンパク質の質量分析 |
| 2.8. イムノブロッティング |
| 2.9. 統計分析 |
| 3. 結果 |
| 3.1. ヨウ素酸ナトリウムによる損傷からラット網膜を保護するアロニア果実エキス |
| 3.2. ラット網膜および血清に対するアロニア果実抽出物処理の抗酸化効果 |
| 3.3. ラット網膜におけるクリスタリンタンパク質の発現に対するアロニア果実抽出物処理の効果 |
| 4.討議 |
| 5.結論 |
本文 |
| 1.はじめに |
| 加齢黄斑変性症(AMD)は、進行性の失明疾患として知られています。 世界中で 1 億 7000 万人以上が加齢黄斑変性症に苦しんでいます [1]。 |
| 現在の治療は、血管新生型または「ウェット」型の加齢黄斑変性症には有効ですが、非血管新生型または「ドライ」型には有効な治療法はありません [2]。 乾性加齢黄斑変性症は、この視力を脅かす状態の人の総数の約 90% を占めています [3]。 ドライ加齢黄斑変性症の発生と発症を予防する方法を開発することが不可欠です。 |
| 加齢黄斑変性症発症の特徴には、光受容体の変性に関連する網膜色素上皮 (RPE) の酸化的損傷によって引き起こされる進行性黄斑変性症が含まれます [4,5,6]。加齢黄斑変性症のメカニズムに関する研究は、この疾患の主な損傷が酸化ストレス/網膜色素上皮への損傷によるものであることを示唆しています [4,5,6]。 したがって、加齢黄斑変性症の治療に関するいくつかの研究は、高用量の抗酸化ビタミンと亜鉛サプリメントを使用した酸化ストレスの抑制に焦点を当てています [7,8]。加齢黄斑変性症が網膜の神経系の損傷にもつながる可能性があることを示唆する研究があります。 臨床研究の結果では、滲出性加齢黄斑変性症 (EXAMD) の末期症例で神経節細胞層 (GCL) ニューロンが 47% 失われていることが示されました [9]。 網膜神経節細胞 (RGC) の死は、光受容体がほぼ完全に失われた後のげっ歯類モデルでも観察されています [10]。 |
| 網膜神経節細胞は網膜の求心性ニューロンであり、視神経の軸索を介して視覚情報が脳の網膜受容核に送られ、さらに分析されます。 多くの研究者は、ラットの網膜への部分的な視神経切断 (ponT) が、活性酸素種 (ROS) の過剰生成と、ミトコンドリアの形態と機能の変化につながる可能性があることを示しています [11,12]。 しかし、視神経切断後の長期的には、有髄軸索の腫れやミエリン鞘の肥厚など、二次的な網膜神経節細胞変性が観察され、視覚障害につながる可能性があります [12,13]。 以前の研究では、α-およびβ-クリスタリンが視神経損傷後の網膜色素上皮の生存率を高める可能性があることが示されています [14,15]。 部分視神経切断後の網膜神経節細胞におけるタンパク質発現に関する 2D 蛍光差分ゲル電気泳動 (DIGE) とそれに続く質量スペクトル (MS) を使用した最近の研究では、非損傷領域の網膜神経節細胞では、8週間後の損傷部位との比較して、6 つのクリスタリンタンパク質(訳者注:クリスタリン(Crystallin)は、動物の眼のレンズに当たる器官、水晶体に存在するタンパク質の一種。重量にして水晶体の1/2〜1/3を占める。ほ乳類の場合、α-、β-、γ-の3種のタンパクの混合物である)の発現が劇的にアップレギュレートされていることがわかりました[16]。 これは、これらのクリスタリンタンパク質が網膜神経節細胞を二次変性から保護する可能性があることを示唆しています。 |
| ある研究では、ビルベリーアントシアニンが、ウサギの網膜の光による酸化損傷から保護する効果があることが示されました [17]。 アントシアニジンには抗酸化作用があり、ラジカルを除去することができると報告されており [18]、加齢黄斑変性症による損傷から網膜を保護する効果がある [19]。 アントシアニジンに関する研究では、アントシアニジンが細胞内活性酸素種を減少させることにより、酸化ストレスによる網膜色素上皮の損傷を防ぐことができることが示されています [20]。 ただし、これらの研究のほとんどは、動物ではなく細胞に焦点を当てています。 |
| アロニア (Aronia melanocarpa) の果実には、抗酸化活性の主要な「有効成分」であると考えられている高レベルのアントシアニジンが含まれています [21]。 ラットモデルは、ヨウ素酸ナトリウム (NaIO3) または強い光によって誘発される加齢黄斑変性症の研究において古典的です [17,22]。 この研究では、進行性加齢黄斑変性症[23,24,25] 中の損傷をシミュレートするために広く使用されているヨウ素酸ナトリウムを使用してラットを加齢黄斑変性症モデルとして扱い、げっ歯類モデルに酸化ストレスを発生させて網膜色素上皮および光受容体の酸化的損傷/細胞死を誘発しました。 ヨウ素酸ナトリウム処理ラット網膜に対するアロニア果実抽出物の保護効果を調査し、ラット網膜における上記のクリスタリンタンパク質の発現を測定することにより、網膜神経節細胞の二次変性の防止に対するその機能を分析しました。 |
| 2.材料と方法 |
| 2.1. 動物 |
| この研究で実験に使用されたすべての動物は、眼科および視覚研究における動物の使用に関する視覚および眼科学研究協会 (ARVO) 声明に従って、手順の下で準備されました。 この研究で使用されるプロトコルは、薬用植物開発研究所の動物倫理委員会によって審査および承認されています (No. SLXD-20201218031)。 オスの Sprague-Dawley (SD) ラット 180 ~ 200 g は、National Institutes for Food and Drug Control (北京、中国) から提供されました。 動物は、実験に使用する前に、22°Cで12時間/12時間の明/暗サイクル条件で1週間維持しました。 |
| 2.2. アロニア果実エキス |
| この研究で使用されたアロニア果実抽出物は、Greater Hinggan Gebei Frigid Zone Biotechnology Co., Ltd. (黒竜江省、中国) から購入した紫がかった赤色の粉末でした。 粉末には、10% のデンプン (粉末の調製時に外から添加)、10.3% のアントシアニジン、およびアロニア メラノカルパの果実からの糖類、タンパク質、および食物繊維を含むその他の水溶性栄養素が含まれています。 |
| 2.3. ラットにおけるヨウ素酸ナトリウム誘発網膜損傷に対するアントシアニジンの保護効果の評価 |
| 合計 36 匹の SD ラットをランダムに 3 つのグループ (NOR、MOD、および AC グループ)、各グループ 12 匹のラットに分けました。 ACグループでは、胃管を使用してアロニア果実抽出物の蒸留水を60 mg/kg 体重相当のアントシアニジン濃度で(アロニア果実エキス末の量は体重1kgあたり600mg)28日間、1日1回経口投与しました。 30 mg/kg 体重のヨウ素酸ナトリウムの単回治療が、アロニア水抽出物の給餌開始後 8 日目に、このグループのラットに静脈内注射されました。 |
| 他の 2 群のラットにはアロニア果実抽出液と等量の蒸留水を経口投与した。 MOD グループでは、8 日目にラットに 30 mg/kg 体重のヨウ素酸ナトリウムを注射しました。これにより、乾性加齢黄斑変性症モデルが確立されました。 NOR 群では、ラットに蒸留水を経口投与し、他の処置は行わなかった。 29 日目に、実験のためにすべてのラットを採取しました (図 1)。 |
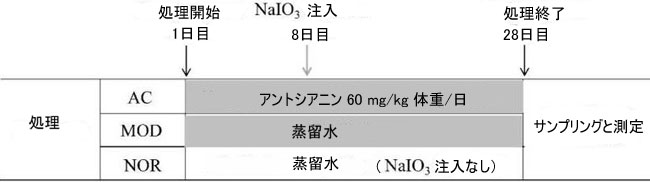 |
| 図 1 ヨウ素酸ナトリウムとアロニア果実抽出物によるラットの治療の概略図 |
| 36 匹の SD ラットをランダムに 3 つのグループ (NOR、MOD、および AC) に分け、各グループに 12 匹のラットを割り当てました。 NOR: 治療なしのコントロール。 MOD: 30 mg/kg 体重の ヨウ素酸ナトリウム尾静脈注射による損傷モデル。 AC: 損傷モデルのアントシアニジン (60 mg/kg 体重) 治療。 |
| 2.4. 網膜電図 |
| 各グループのラットの全視野網膜電図(ERG)は、全視野網膜電図記録システム(D430 Diagnosis、米国)を使用して記録されました。全視野網膜電図の測定前に、ラットを 24 時間暗順応させた [26]。 ケタミン塩酸塩とキシラジン塩酸塩の混合物をそれぞれ 100 mg/kg と 15 mg/kg の投与量で筋肉内注射して、ラットの麻酔を誘発しました。 0.5% トロピカミドと 0.5% 塩酸フェニレフリンを含む点眼薬をラットの眼に 20 分間投与し、瞳孔を散大させました。 発光ダイオード (LED) 電極が両方の角膜に配置されました。 同一の参照電極を中頭皮の下に配置し、接地電極を試験ラットの両下肢の皮下に配置した。 Scotopic 0.01ERG、Scotopic 3.0ERG、Scotopic 3.0 振動電位、Photopic 3.0ERG、および Photopic 3.0 フリッカーの全視野網膜電図が実施されました。 a波とb波のピーク時間と振幅の両方が記録され、b波の振幅が統計的に分析されました。 ラットの各グループから 12 のサンプルを測定しました。 |
| 2.5. 網膜のヘマトキシリンおよびエオシン (H&E) 染色 |
| 安楽死後、各群のラットの両眼を摘出した。 眼球を摘出し、20% イソプロパノール、2% トリクロロ酢酸、および 2% 塩化亜鉛を含む 4% パラホルムアルデヒドで室温で 2 時間固定しました。 角膜、虹彩、水晶体、および部分的な硝子体を取り除いた後、目の残りの部分をアルコールで 24 時間脱水し、パラフィンに包埋しました。 網膜全体のサンプルを厚さ 4 μm の矢状切片に切断し、ヘマトキシリンおよびエオシンで染色しました。 各セクションについて、網膜全体のデジタル化された画像をデジタル カメラ (Leica DMi8、Wetzlar、ドイツ) で 200 倍の倍率で撮影しました。 外核層(ONL)の厚さは、Image Jソフトウェア(米国国立衛生研究所、ベセスダ、メリーランド州、米国)で測定しました。 視神経の両側から始めて、各セグメントを 0.5 mm 離して、各網膜切片の 12 箇所を測定しました。 12 回の測定値を平均して ONL の厚さを算出しました [27]。 |
| 2.6. マロンジアルデヒド(MDA)の測定 |
| 各群のラットから静脈血を採取し、卓上遠心分離機(Eppendorf、Hamburg、Germany)で 3000 rpm で 10 分間遠心分離し、各サンプルの上清をマロンジアルデヒド濃度測定に使用しました。 各群のラットの眼球から採取した網膜サンプルをリン酸緩衝生理食塩水 (PBS、pH 7.4) でホモジネートし、3500 rpm/min で 10 分間遠心分離し、上清をマロンジアルデヒド測定用に収集しました。 各サンプルの上清中のマロンジアルデヒドレベルは、測定キット (Nanjing Jiancheng Bioengineering Institute、南京、中国、#20210304) を使用して、532 nm の波長で分光測光法により決定されました。 ラットの各グループから 8 つのサンプルが検出されました。 |
| 2.7. 網膜タンパク質の質量分析 |
| ラット網膜タンパク質は、以前のプロトコルに従ってタンデム 質量スペクトル(MS) を使用して同定および分析されました [28]。 ラットの各グループから 3 つの網膜サンプルを準備しました。 各群からの網膜組織を、8M尿素緩衝液中での超音波処理によって溶解した。 トリプシンで消化した後、オンラインEASY-nLC 1200ナノ高速液体クロマトグラフィー(HPLC)システム(Thermo Fisher Scientific、ウォルサム、マサチューセッツ州、米国)と組み合わせたOrbitrap Q Ex-active HF質量分析計でペプチドを分析しました。 ペプチド混合物は、逆相ナノ HPLC C18 カラム (プレカラム: 0.1 × 20 mm、3 μm; 分析カラム: 0.15 × 120 mm、1.9 μm) で 600 nL/分の流速で 78- 分グラジエント: 6 ~ 9% の溶媒 B で 2 分間、9 ~ 13% で 8 分間、13 ~ 26% で 40 分間、26 から 38% で 20 分間、38 から 100% で 1 分間、100% で 7 分間 分 (溶媒 A、水、溶媒 B、アセトニトリル、0.1% ギ酸)。 エレクトロスプレー電圧は 2.2 kV でした。 ペプチドは、フルスキャン モードで 120,000、MS/MS モードで 15,000 の分解能を持つデータ依存型 MS/MS 取得モードによって分析されました。 フルスキャンは、Orbitrap で質量/電荷比 250 から 1800 で処理されました。 各スキャンで上位 20 個の最も強いイオンが自動的に選択され、正規化された衝突エネルギー 32% の高エネルギー衝突解離 (HCD) フラグメンテーションが行われ、Orbitrap で測定されました。 一般的な質量分析条件は次のとおりです。自動ゲイン制御のターゲットは、フル スキャンでは 3 × 106 イオン、MS/MS スキャンでは 2 × 105 イオンでした。 最大注入時間は、フル スキャンで 35 ミリ秒、MS/MS スキャンで 80 ミリ秒でした。 動的排除は18秒間使用されました。 サンプル量が限られているため、各サンプルは 1 つの技術的複製で分析されました。 ラットの各グループにおける標的タンパク質の発現は、各グループからの3つの網膜サンプルの測定値の平均によって決定されました。 質量分析の結果は、PEAKS Studio (ウォータールー、カナダ) を使用して分析および定量化されました。 |
| 2.8. イムノブロッティング |
| 網膜組織のタンパク質を NETN バッファー (50 mM Tris-HCl、pH 8.0、150 mM NaCl、0.2% Nonidet P-40、および 2 mM EDTA) で抽出し、各サンプルの 20 μg のタンパク質を 12% SDS-PAGE で分離しました。。 電気泳動後、タンパク質をポリフッ化ビニリデン膜に転写した。 メンブレンは、Tween/Tris 緩衝生理食塩水 (TBST) 中の 5% スキムミルクで洗浄して非特異的結合をブロックし、α-クリスタリン A 鎖および γ-クリスタリン S に対する一次抗体とともにインキュベートしました。 免疫ブロットは、SuperSignal Western Pick Plus (#34577、Thermo Scientific) [29] を使用して実行されました。 |
| 2.9. 統計分析 |
| 結果は、平均 ± 標準偏差として表示されます。 グループ間の違いは、一方向ANOVA、続いてTukeyの検定またはKruskal-Wallisのノンパラメトリック検定、そしてDunnの検定によって評価されました。 p < 0.05 は統計的に有意と見なされました。 すべての統計分析は、Prism 8.0 (GraphPad Software、米国カリフォルニア州サンディエゴ) を使用して実行されました。 |
| 3. 結果 |
| 3.1. ヨウ素酸ナトリウムによる損傷からラット網膜を保護するアロニア果実エキス |
| 全視野網膜電図は、全体的な網膜機能の客観的な尺度です。 全視野網膜電図は、光刺激に対する網膜の電気的反応です。 閃光または明るい外観は、二相性の負/正の波形を引き出します。 桿体と錐体の受容体レベルで発生するa波は、最初の大きな負の波です。 網膜中部に由来するb波は、次の大きな正の成分です。 全身性代謝障害は通常、全視野網膜電図の b 波の振幅、特に暗所視の薄暗いフラッシュ全視野網膜電図を減少させます [30,31]。 |
| ISCEV(国際臨床電気生理学的視覚学会)規格に従って、各群のラット網膜における5つの異なる電流を測定した。 結果は、5 つの異なる測定すべての全視野網膜電図b 波振幅が、NOR グループと比較して MOD グループで大幅に減少したことを示しました (p < 0.01、図 2)。 これは、MOD グループのラットの網膜に重大な損傷があることを示しています。 Scotopic 0.01ERG、Photopic 3.0ERG、および AC グループの Photopic 3.0 フリッカーにおける ERG b 波振幅の減少は、MOD グループと比較して減少しました (p < 0.05、図 2B)。 ISCEV 規格によると、Scotopic 0.01ERG は双極細胞の桿体駆動応答を示し、Photopic 3.0ERG は錐体システムの応答を示し、b 波は錐体上および錐体外の双極細胞から発生します。 そしてPhotopic 3.0のちらつきは、敏感な錐体経路駆動型の反応です。 結果は、ヨウ素酸ナトリウムによって誘発された網膜の損傷が AC グループで軽減され、アロニア果実抽出物がラットの網膜、特に錐体系に対して潜在的な保護効果を示したことを示唆しています。 |
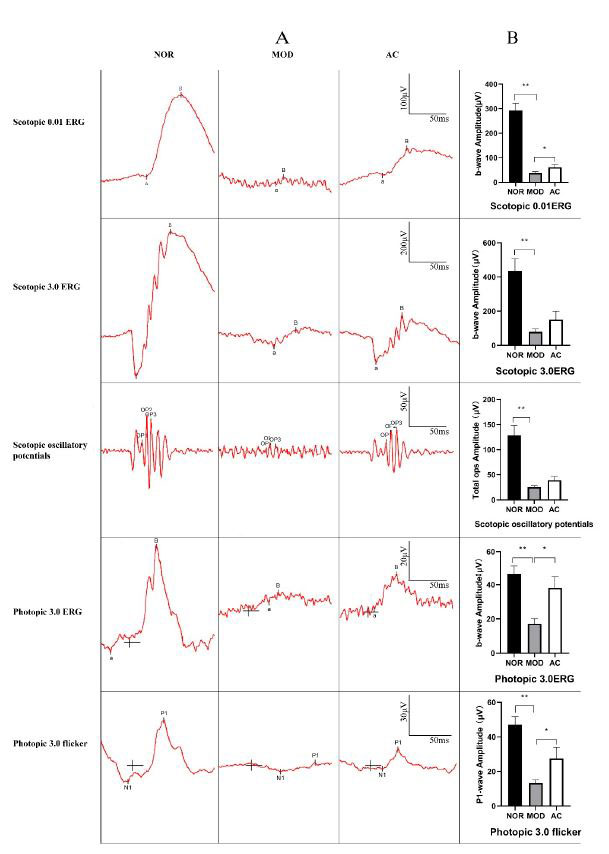 |
| 図 2 異なる処理によるラットの全視野網膜電図 |
| 各群のラットを麻酔し、網膜電図 (ERG) で分析しました。 (A) 全視野網膜電図は、ヨウ素酸ナトリウム注射後 21 日目に記録されました。 (B) Scotopic 0.01ERG、Scotopic 3.0ERG、および Photopic 3.0ERG の b 波振幅の 3 つのグループの平均と、Scotopic 3.0 振動 (3 ops) および P1 波振幅の合計振幅。 示されている b 波振幅のデータは、平均 ± 標準偏差 (n = 12) です。 (*) p < 0.05、(**) p < 0.01 (一元配置分散分析とその後のテューキー検定)。 |
| ラット網膜のヘマトキシリンおよびエオシンで染色された写真は、NOR グループが明確な網膜構造と層を持ち、細胞核がきれいに整列していることを示しています。 MOD サンプルでは、網膜は無秩序な構造を示し、外側と内側の核層 (ONL と INL) が混乱した細胞層の減少を示しました。 比較すると、AC グループは網膜構造と細胞層の改善を示し、核の配置は MOD グループと比較して比較的順調でした (図 3A)。 ヨウ素酸ナトリウム処理は、ONL の厚さを劇的に減少させ、漸進的な ONL の薄化を示唆しました。 しかし、アロニア果実抽出物の投与により、ヨウ素酸ナトリウムによる損傷を軽減し、ONL の厚さを維持することができます (図 3B)。 |
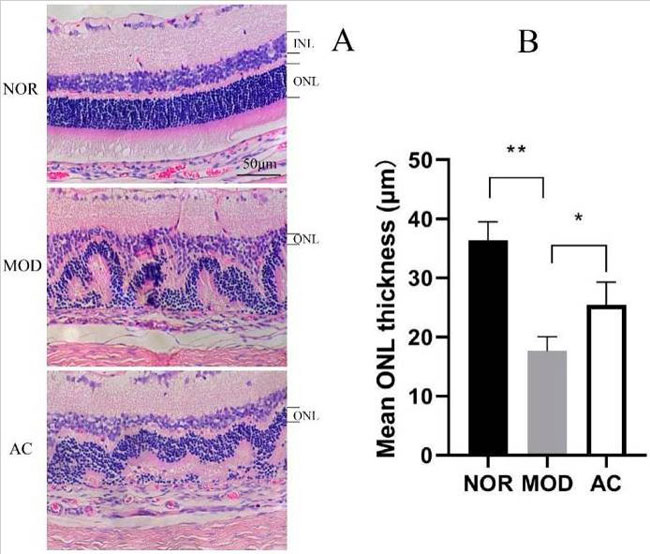 |
| 図 3 ラット網膜の構造および外核層(ONL)の厚さに及ぼすアロニア果実抽出物の影響 |
| (A) 倍率 200 倍で撮影したラット網膜のヘマトキシリンおよびエオシン染色切片の画像。 (B) 網膜の外核層の厚さ。 データは平均 ± 標準偏差 (n = 8) です。 (*) p < 0.05、(**) p < 0.01 (一元配置分散分析とその後のテューキー検定)。 |
| 3.2. ラット網膜および血清に対するアロニア果実抽出物処理の抗酸化効果 |
| マロンアルデヒド (MDA) は、多価不飽和脂肪酸の過酸化生成物であり、酸化ストレスを測定するための「生物学的マーカー」として広く使用されています [32]。 図 4A に示すように、NOR グループのラット網膜のマロンアルデヒドレベルは 2.45 ± 0.19 nmol/mg でした。 NaIO3 処理により、マロンアルデヒドレベルが 3.87 ± 0.74 nmol/mg に大幅に増加しましたが、アロニア果実抽出物処理により、ラット網膜のマロンアルデヒドレベルが 1.88 ± 0.12 nmol/mg に大幅に減少しました (p < 0.01、図 4A)。 |
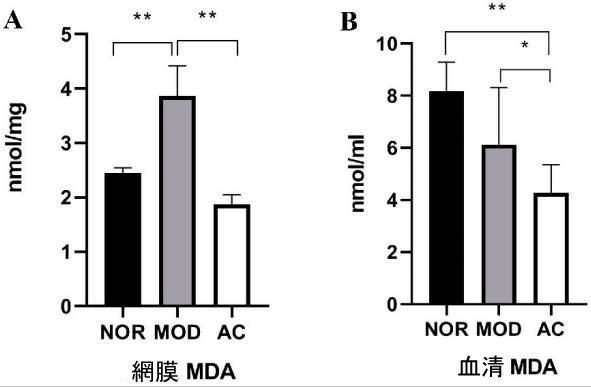 |
| 図 4 ラットの網膜 (A) および血清 (B) におけるマロンジアルデヒド (MDA) レベル |
| データは平均 ± 標準偏差 (n = 8) です。 (*) p < 0.05、(**) p < 0.01 (一元配置分散分析とその後のテューキー検定)。 |
| NOR、MOD、および AC グループのラット血清中のマロンアルデヒド濃度は、それぞれ 8.18 ± 1.12、7.31 ± 1.53、および 4.27 ± 1.09 nmol/mL でした (図 4B)。 MOD グループと NOR グループの間に統計的な差はありませんが、AC グループの血清 MDA レベルは、MOD グループと NOR グループよりも有意に低くなっています (p < 0.05、図 4B)。 これらの結果は、ヨウ素酸ナトリウム処理がラット血清ではなくラット網膜でより多くの酸化ストレスを引き起こし、アロニア果実抽出物の投与がラットの網膜と血液の両方で酸化ストレスを軽減できることを示唆しています。 |
| 3.3. ラット網膜におけるクリスタリンタンパク質の発現に対するアロニア果実抽出物処理の効果 |
| 6 つのクリスタリン タンパク質のグループが、ラットの網膜における網膜神経節細胞の二次変性の防止に一定の効果があることが示唆されています [16]。 ラット網膜神経節細胞に対するアロニア果実抽出物の潜在的な影響を調査するために、網膜サンプルで発現した総タンパク質を分析しました。 その結果、MOD群のラット網膜では、上記クリスタリンタンパク質のうち5つのタンパク質がわずかに増加していた。 NOR グループと比較した MOD グループは、α-クリスタリンA鎖(αA)、β-クリスタリンB2(βB2)、β-クリスタリンA3(βA3)、α-クリスタリンB鎖(αB)の発現、および γ-クリスタリン S (γS) は、それぞれ 1.48、1.43、1.84、1.17、および 1.56 倍増加しました(p > 0.05、図 5A)。 しかし、ACグループの網膜サンプルでは、これらのクリスタリンタンパク質のはるかに高い発現が観察されました。 AC グループでは、5 つのクリスタリン タンパク質、αA、βB2、βA3、αB、および γS の発現が、NOR グループと比較して、それぞれ 7.38、7.74、15.30、4.86、および 9.14 倍増加しました。 (p < 0.05、図 5A)。 結果は、アロニア果実抽出物処理が、ストレス状態で保護クリスタリンタンパク質のはるかに高いアップレギュレーションをもたらしたことを示唆しています。 |
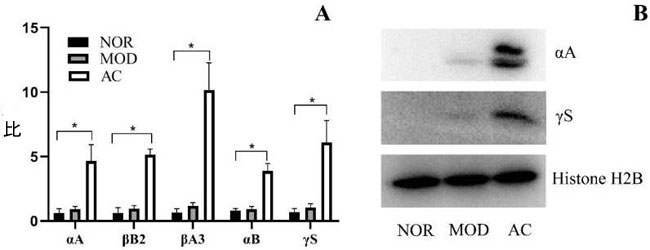 |
| 図 5 ラット網膜におけるクリスタリンタンパク質の発現 |
| (A) α-クリスタリン A 鎖 (αA)、β-クリスタリン B2 (βB2)、β-クリスタリン A3 (βA3)、α-クリスタリン B 鎖 (αB)、γ-クリスタリン S (γS) の発現を質量で決定 分光測定。 NOR群のNo.1サンプル(NOR-1)の各タンパク質の発現量を1倍とし、それ以外のサンプルのクリスタリンタンパク質の発現量をNOR-1に対する比率で表した。 表示されているデータは、平均 ± 標準偏差 (n = 3) です。 (*) p < 0.05 (Kruskal-Wallis 検定とそれに続く Dunn 検定)。 (B) 網膜サンプル中のα-クリスタリン A 鎖と γ-クリスタリン S の存在を示すイムノブロッティングの結果。 各サンプルのタンパク質ヒストン H2B を内部参照として測定し、各サンプルの基本的なタンパク質発現を示しました。 |
| ラット網膜におけるαAおよびγSの量は、イムノブロッティングを使用して検出されました。 図 5B に示すように、NOR グループでは αA と γS の両方が検出されませんでした。 MOD グループでは、非常に少量の 2 つのタンパク質が検出されました。 ただし、αA と γS は、MOD グループと比較して、AC グループのラット網膜で有意に高い発現を示しました。 結果は、図 5A に示す質量スペクトルの結果を確認しました。 |
| 4.討議 |
| アロニア メラノカルパの果実はアントシアニンの最も豊富な供給源の 1 つであり、その含有量は新鮮なアロニア果実 100 g あたり最大 460 mg です [33]。 有色ウサギの可視光誘発網膜変性モデルにおいて、ビルベリーアントシアニン(BAE)の経口投与は、ERG b波の減少を緩和し、外側の核層の菲薄化を抑制すること、および保護効果は抗酸化活性と関連していたことが報告されていました。[17]。 その結果、MOD群と比較して、アロニア果実抽出物を投与したAC群のラットでは、細胞層障害の抑制とともに網膜電図b波の緩和が減少し、外側の核層の菲薄化が観察されました(図2および図3)。 アントシアニジンはポリフェノール化合物のグループであり、人間の健康における治療効果に関連する抗酸化活性が広く証明されています [18,20]。 網膜電図およびヘマトキシリンおよびエオシン染色実験で観察されたラット網膜に対するアロニア果実抽出物の保護効果は、アントシアニンの抗酸化活性による可能性が非常に高いです。 しかし、私たちの研究におけるアロニア果実抽出物治療の保護効果は、マウス網膜変性モデルで30〜300 mg / kg体重のアントシアニジン、プロアントシアニジン、またはポリフェノールを使用した以前の報告ほど重要ではないことに気付きました。 網膜に対するアントシアニンジンの保護効果は用量依存的であり、最適な用量は約 100 mg/kg である可能性があります [17,34,35,36]。 アントシニジンの投与量を増やすと、より良い結果が得られる場合があります。 |
| 光受容体の変性に関連する網膜色素上皮の酸化的損傷は、通常、加齢黄斑変性症発症の特徴であると考えられています [4,5,6]。 一部の研究者は、光受容体が完全に失われた後、ラットの網膜の神経系が損傷を受ける可能性があることを示しました [10]。 通常、光受容体の変性は加齢黄斑変性症に関連する一般的な特徴と考えられているため [4,5,6]、網膜神経節細胞死は加齢黄斑変性症の「二次的」損傷として示唆される可能性があります。 最近の研究では、ラットの網膜への視神経切断は、αA、αB、βA2、βA3、βB2、およびγSを含むクリスタリンタンパク質のグループのアップレギュレーションが、一次変性よりも二次変性に対して網膜神経節細胞により保護効果があることを発見しました。 αB および βA2 クリスタリンの発現は、損傷した領域ではそれぞれ 1.4 倍および 1.2 倍しか増加しなかったが、損傷していない領域ではこれらのタンパク質の 3.5 倍および 2.1 倍増加したことが報告されている [16]。 . 私たちの実験では、クリスタリンタンパク質のより重要なアップレギュレーションが観察されました。 アロニア果実抽出物で処理されたACグループでは、αA、βB2、βA3、αB、およびγSを含む5つのクリスタリンタンパク質が、それぞれ7.38倍、7.74倍、15.30倍、4.86倍、および9.14倍アップレギュレートされました(図5A )、 これは、アロニア果実抽出物の投与がラット網膜の神経系を二次変性から保護する可能性があることを示唆しています。 私たちの実験でのもう 1 つの観察結果は、これら 5 つのクリスタリンタンパク質の発現が ヨウ素酸ナトリウム処理 MOD グループでもわずかにアップレギュレートされたことですが、MOD グループと NOR グループの間に統計的な差はありませんでした (図 5A)。 これらの観察結果は、ラットの各グループからの網膜サンプルの αA と γS の量を測定したイムノブロッティングの結果とも一致していました (図 5B)。 これは、クリスタリンタンパク質の発現が、ラット網膜神経細胞を二次変性から保護する先天的な保護メカニズムである可能性が高いことを示唆している可能性があります。 |
| 二次変性による網膜神経系の抑制に関与するクリスタリンタンパク質のアップレギュレーションの詳細なメカニズムは不明のままです。 クリスタリンタンパク質はシャペロンタンパク質ファミリーのメンバーとして知られているため、それらは通常、細胞内のタンパク質品質管理システムの一部と見なされ、「タンパク質が適切な場所と時間で正しく折りたたまれ、機能する」ことを保証します[37, 38,39]。 したがって、アップレギュレートされたクリスタリンタンパク質の潜在的な保護メカニズムの1つは、ラット網膜のタンパク質フォールディングを調節することによるものです。 私たちの質量分析の結果は、MOD グループと比較して AC グループの網膜において、網膜神経節細胞のアポトーシスを調節する因子であるカスパーゼ-3 タンパク質のダウンレギュレーションも示しました [40] (データは示していません)。 カスパーゼ-3の発現低下は、アロニア果実抽出物が網膜神経細胞をプログラム細胞死からも保護する可能性があることを示唆しています。 |
| 5.結論 |
| 結論として、この研究では、ヨウ素酸ナトリウムを使用したドライ加齢黄斑変性症モデルの確立に成功し、アロニア果実抽出物が抗酸化活性とクリスタリンタンパク質発現の上方制御によって網膜を保護できることを発見しました。 したがって、高アントシアニジンを含むアロニア果実抽出物を補給することは、加齢黄斑変性症および網膜変性に関連するその他の損傷を防ぐためのアプローチである可能性があると予想できます。 |
参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
この文献は、Nutrients 2021, 13, 4411.に掲載されたProtection of Aronia melanocarpa Fruit Extract from Sodium-Iodate-Induced Damages in Rat Retina.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |