ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(ヨーグルト、ビフィズス菌:最新の研究) |
ヒトの健康に対するBifidobacterium longum subsp longum BB536の有益な効果:主要な作用としての腸内微生物叢の調節 |
Chyn Boon Wong et al., |
| Journal of Functional Foods 54 (2019) 506–519 |
|
要約 |
| プロバイオティクスは、人間の健康を促進する上で大きな期待を示しています。 有望な証拠にもかかわらず、多くの利用可能なプロバイオティクスの臨床的有効性に関する情報はほとんどなく、その作用メカニズムはしばしば不明です。 Bifidobacterium longum subsp longum BB536は、胃腸、免疫、感染症の緩和に人間が使用した長い歴史を持つ、臨床的に効果的で定評のある多機能プロバイオティクスです。 このレビューでは、最も関連性の高い臨床および動物研究からのBB536の機能的利点を要約し、その作用メカニズムを理解するための理論的基礎を提供します。 主要な臨床所見は、BB536が腸のコミュニティの生理学的活動を調整するための微生物叢モジュレーターとして作用する可能性があることを指摘しています。 具体的には、BB536は管腔代謝を調節し、腸内細菌叢を安定化し、最終的には宿主とミクロビオームの相互作用内で微調整された恒常性バランスを駆動します。 |
はじめに |
|
ヒトの消化管には、腸内微生物叢と総称される数兆の微生物で構成される複雑で動的な微生物生態系が宿っています(Thursby&Juge、2017)。 これらの腸内微生物は、免疫機能の維持、炭水化物代謝、代謝恒常性など、宿主の生理学の多くの側面を調節しています(Nagpal et al.,2018)。 重要な進歩により、炎症性腸疾患(IBD)を含むいくつかの非常に一般的な健康障害が示されました。 急性感染性下痢、および2型糖尿病や肥満などの慢性疾患は、微生物叢の変化と宿主微生物叢の相互作用の不均衡に関連しています(Liang、Leung、Guan & Au、2018)。 |
| 最近の数十年で、プロバイオティクスによる栄養介入などの外部アプローチによる微生物組成の積極的な調節が、ホストに前例のない健康影響を与える可能性があることを非常に多くの証拠が強く示唆しています(Azad、Kalam、Sarker、Li、&Yin、2018)。 プロバイオティクスは、適切な量を摂取すると、宿主に健康上の利益をもたらす生きた微生物として定義されます(Hill et al。、2014)。 プロバイオティクスの介入による腸内微生物叢の操作は、宿主の腸と免疫の健康に予防効果と治療効果の両方を及ぼすことが報告されています(Scott、Jean-Michel、Midtvedt、&van Hemert、2015年)。 それにもかかわらず、プロバイオティクス製品の使用と多様性が拡大するにつれて、異なる系統の作用メカニズム、安全性プロファイル、起源、および有効性の違いにより、適切なタイプのプロバイオティクスの選択が困難になっています。 最近の研究では、プロバイオティクスの多くの特性が株特異的であり、すべてのプロバイオティクスが同等に安全で効果的ではないことが指摘されています(McFarland、Evans、&Goldstein、2018)。 さらに、多くの利用可能なプロバイオティクス株のin vitroおよびヒトのデータと作用メカニズムの実質的な裏付けを備えた治療特性が欠けていることがよくあります。 |
| Bifidobacterium longum subsp longum BB536(BB536として指定)は、人間に多くの深刻な健康上の利点がある確立されたプロバイオティクス株の1つです(Xiao、2009)。 BB536は、半世紀にわたってその発見以来プロバイオティクスとして使用されており、その有効性を明らかにするために多くの研究が行われてきました。 それにもかかわらず、その有益な特性についての詳細なレビューが不足しており、医療専門家と一般市民の両方が、より良い情報に基づいたプロバイオティクスの選択を支援する上で、より信頼できる健康強調表示を提供できます。 このレビューでは、最も関連性の高い臨床および動物研究からBB536の重要な健康上の利点を要約し、BB536の機能的な健康への影響をよりよく理解するための基本的な枠組みを提供しようとします。 私たちは、プロバイオティクスのより調整されたターゲットを絞ったアプリケーションには、有効性の人間の実証と潜在的な作用メカニズムの包括的な知識が不可欠であると考えています。 ここでは、腸内マイクロバイオームの調節が、ヒトにおけるBB536の主要な有益な作用であることを強調しています。 |
2.ビフィドバクテリウムロンガムBB536 |
| 2.1 起源と特徴 |
| BB536は、1969年に母乳で育てられた健康な乳児の腸から生まれました。 BB536は、機能性食品成分として牛乳ベースのドリンク、ヨーグルト、粉ミルク、栄養補助食品などのさまざまな製品に組み込まれ、40年以上にわたって30か国以上で販売されています。 1996年、BB536を含むヨーグルト製品は、健康のために、腸内ビフィズス菌の存在量の増加、および腸内環境の改善と調整における利益の実証する(He&Benno、2011)十分な科学的証拠を持つ食品として、日本の保健省から特別保健用食品(FOSHU)ステータスを授与されました。 |
| BB536は、グラム陽性、嫌気性、カタラーゼ陰性のrod体で、不規則な形態をしています。 BB536は、人間の腸に非常にアクセスしやすく、粉ミルク、ヨーグルト、発酵乳など、さまざまな最終製品で安定しています(阿部、宮内、内島、八重島、岩槻、2009;阿部、富田、八重島、岩槻、2009;バロンゲ &Grill、1993; Odamaki、Xiao、Yonezawa、Yaeshima、&Iwatsuki、2011; Yonezawa et al、2010); 保管中の優れた安定性と、消費までのプロバイオティクス食品の高い生存性を備えています。 |
| 2.2 安全性 |
| BB536は安全性について十分に評価されており、2009年に米国食品医薬品局(FDA)の一般安全(GRAS)通知インベントリ(GRN No. 268)にリストされています。 GRASステータスは、BB536がプラスミドを含まず、有害な代謝活性を示さない非病原性、非毒性、非溶血性、非抗生物質耐性のプロバイオティクスバクテリアであるという証拠に基づいて付与されました(FDA、2009; 桃瀬、1979;トスカーノ、デヴェッキ、ガブリエリ、ズッコッティ、ドラゴ、2015;小、高橋、小田巻,、八重島、岩槻,、 2010)。 BB536は主にL-乳酸を生成しますが、D-乳酸の生成はごくわずかです(FDA、2009)。 さらに、BB536は、胆汁酸塩の加水分解を触媒する共役胆汁酸塩加水分解酵素(BSH)酵素を保有することが実証されました(Grill、Schneider、Crociani、&Ballongue、1995)。 BB536は、選択された胆汁酸塩(タウロコール酸、グリココール酸、タウロケノデオキシコール酸、グリコケノデオキシコール酸、タウロデオキシコール酸、およびグリコデオキシコール酸)の80〜95%を非共役化することができました。 |
| 特定の二次胆汁酸、特にデオキシコール酸およびリトコール酸の高い生理的レベルがDNA損傷を引き起こし、結腸発がんを促進することが長い間知られています(Bernstein、Bernstein、Payne、Dvorakova、&Garewal、2005; MJ Hill et al。、1975 ; Nagengast、Grubben、およびVan Munster、1995年)。 これを考慮して、二次胆汁酸産生細菌の投与の安全性に関して懸念が生じる場合がある。 注目すべきことに、非共役胆汁酸塩はBB536によって生成される唯一の化合物でしたが、脱水またはヒドロキシル化製品などの二次胆汁酸(ケノデオキシコール酸の脱ヒドロキシル化から生成される肝毒性および発がん性リトコール酸)は、胆汁酸塩の完全な生体内変化では検出されませんでした(Grill 、Manginot-Dürr、Schneider、およびBallongue、1995年)。 |
| BB536の急性および慢性毒性学的特徴に関する研究により、BB536の経口投与は、それぞれマウスおよびSDラットモデルの両方で死および毒性症状を引き起こさないことが明らかになった(Momose、1979)。 結果は、BB536の消費による急性および慢性毒性がないことを示しています。 毒性に加えて、プロバイオティクス投与の安全性について疑問と懸念が提起されており、細菌の転座は菌血症、心内膜炎、敗血症などの疾患を引き起こすプロバイオティクスの潜在的な危険因子であると想定されています。 したがって、転座能力に関するビフィズス菌プロバイオティクスの評価は、特に乳児、高齢者、免疫不全の人にとって重要な安全パラメーターです。 |
| この目的に向けて、健康な4週齢のマウスに7日間高用量(9.3×1011 CFU / kg /日)でBB536を経口投与する生体内試験を実施しました(Abe et al。、2010)。 この研究は、BB536が血液、肝臓、脾臓、腎臓、腸間膜リンパ節に移行せず、腸表面の損傷を誘発しないことを明らかにしました。 さらに、BB536のムチン分解活性に関する追加のin vitro試験により、BB536はコントロール型株としてin vitroでムチンを分解できないことが明らかになった(Abe et al。、2010)。 総合すると、これらの研究は、プロバイオティクスとしての食品でのBB536の安全な使用に対する実質的なサポートを提供します。 |
3. BB536の臨床効果 |
| 3.1 胃腸障害の緩和 |
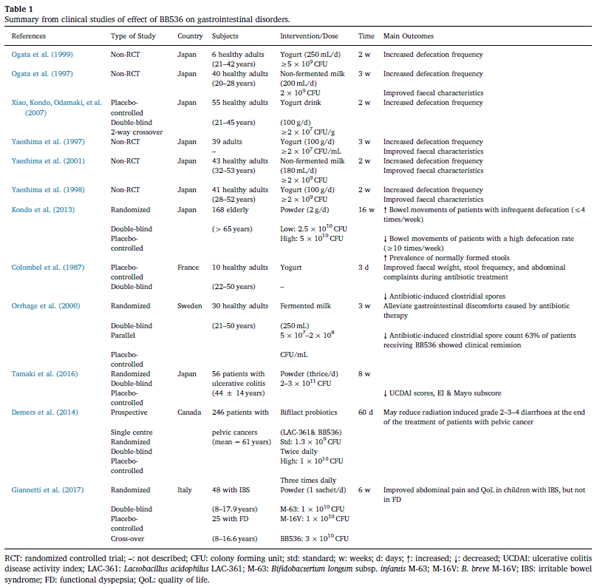
| プロバイオティクスは歴史的に、多くの形態の胃腸障害の治療と予防に使用されてきました。BB536は、消化器疾患の改善に最も効果的なプロバイオティクス株の1つとして長い間認識されてきました(表1)。 臨床証拠の増加により、ヨーグルト、ヨーグルト飲料、非発酵乳など、BB536を含む乳製品の摂取により、便秘のある健康な成人の排便頻度と糞便特性が改善されることが示されています(Ogata et al。、1997、1999; Xiao 、近藤、小damら、2007;八重島ら、1997、1998、2001)。 |
便秘は、臨床診療で遭遇する最も一般的な胃腸障害の1つであり、健康関連の生活の質に大きな影響を及ぼします。 有病率は年齢とともに増加し、長期介護施設の高齢者の間でより頻繁に見られます(De Giorgio et al。、2015)。 BB536は、入院した高齢患者の腸の運動性を調節し、排便頻度を正常化することが報告されています(Kondo et al。、2013)。 経腸栄養法を受けている65歳以上の168人の患者を含む2つの二重盲検プラセボ対照ヒト介入試験では、BB536の投与(それぞれ2.5 x 1010および5 x 10 10 CFUの低用量と高用量の両方で) 16週間は、プラセボ群と比較して、排便頻度の低い患者の排便を有意に増加させた(週4回以下)(近藤他、2013年)。 さらに、BB536群では排便の頻度が高い(週10回以上)患者の排便の有意な減少が観察されたが、プラセボ群では観察されなかった(Kondo et al。、2013)。 正常に形成された便の有病率も、プラセボ群と比較してBB536群で有意に増加しました 。まとめて、これらの発見は、定期的な排便頻度の維持と腸内環境の調整における効果的なモジュレーターとしてのBB536の機能的特性を例示しています。
|
| さらに、排便問題の正常化に効果的なプロバイオティクス株として実証されたBB536の消費は、抗生物質による腸内微生物叢の変化を軽減し、抗生物質療法によって引き起こされる胃腸の不快感を軽減することができました(Colombel、Cortot、Neut、Romond、1987 ; Orrhage、Sjöstedt、&Nord、2000)。 便秘と抗生物質に関連する下痢の緩和に加えて、BB536の消費は証拠の強さは堅牢ではありませんが、潰瘍性大腸炎(Tamaki et al。、2016)、放射線誘発性胃腸機能障害(Demers、Dagnault、&Desjardins、2014)、過敏性腸症候群(Giannetti et al。、2017)などの胃腸疾患の治療にも使用されています。 胃腸疾患の治療におけるBB536の有効性を明らかにするには、さらに臨床試験が必要になります。 総合すると、二重盲検プラセボ対照試験で評価されたこれらの臨床所見は、ヒトの胃腸障害の改善に効果的なプロバイオティクス株としてBB536の健康上の利点を実証しています。 |
| 3.2 腸内微生物学への影響 |
| 腸内細菌叢は、腸の恒常性と健康の重要な決定因子です。 いくつかの研究は、摂動した腸内微生物叢、例えば、微生物多様性の減少-微生物叢の不安定性につながる機能不全の生態系の兆候-が消化器および代謝障害を含むヒトのいくつかの疾患に関連していることを示しています(Gong、Gong、Wang、Yu 、&Dong、2016; Mosca、Leclerc、&Hugot、2016)。 |
| 食事は、腸内細菌叢の構造と活動に影響を与える主要な要因の1つです。 David et al(2014)による注目すべき研究は、動物ベースまたは植物ベースの食事の短期間の消費が微生物プロファイルを変える可能性があることを示唆しています。 植物ベースの食事ではなく動物ベースの食事が胆汁耐性細菌(Alistipes、Bilophila、Bacteroides)の量を増加させ、食事植物多糖を代謝するファーミキューテスのレベルを減少させることが明らかになりました。 これらの変化、特にBilophila wadsworthiaの存在量と活性の増加は、IBD(Devkota et al。、2012)と大腸がん(Han et al。、2018; Yazici et al。、2015)の発症に関連していることがわかりました。 |
| 食事は、腸内細菌叢の構造と活動に影響を与える主要な要因の1つです。 David et al(2014)による注目すべき研究は、動物ベースまたは植物ベースの食事の短期間の消費が微生物プロファイルを変える可能性があることを示唆しています。 植物ベースの食事ではなく動物ベースの食事が胆汁耐性細菌(Alistipes、Bilophila、Bacteroides)の量を増加させ、食事植物多糖を代謝するファーミキューテスのレベルを減少させることが明らかになりました。 これらの変化、特にBilophila wadsworthiaの存在量と活性の増加は、IBD(Devkota et al。、2012)と大腸がん(Han et al。、2018; Yazici et al。、2015)の発症に関連していることがわかりました。 |
| 表1 胃腸障害に対するBB536の効果の臨床研究からの要約。 |
|
| RCT:ランダム化比較試験。 –:説明なし。 CFU:コロニー形成ユニット。 std:標準; w:週; d:日数; ↑:増加; ↓:減少。 UCDAI:潰瘍性大腸炎疾患活動性指数; LAC-361:アシドフィルス菌LAC-361; M-63:ビフィドバクテリウムロンガムsubsp。 乳児M-63; M-16V:B.ブレーブM-16V; IBS:過敏性腸症候群; FD:機能性消化不良; QoL:生活の質。 |
| この研究は、7日間のベースライン、動物ベースの食事の5日間給餌、およびバランスの取れた食事の2週間給餌で構成されていました。 被験者はランダムに3つのグループに割り当てられました。 (i)YABグループとして、動物ベースの食事期間とバランスの取れた食事期間の両方でBB536ヨーグルトを摂取した被験者、 (ii)YBグループとしてバランスの取れた食事期間中にのみBB536ヨーグルトを摂取した被験者、 (iii)介入としてコントロール(CTR)グループとしてヨーグルトを摂取しなかった被験者。 以前のレポート(David et al。、2014)に沿って、動物ベースの食事は、5日間の摂取後に微生物群集の変化を誘発し、Bilophila、Odoribacter、Dorea、およびRuminococcusの相対量が大幅に増加しました( ラクノスピラ科に属する)また、YBおよびCTRグループの両方でビフィズス菌のレベルが大幅に減少しました。 対照的に、BB536ヨーグルトの摂取は健康な腸内微生物叢を再確立しました。動物ベースの食事によって引き起こされる微生物プロファイルの変化は、Ruminococcusを除くYABグループでは示されませんでした。 |
| エンテロトキシン産生Bacteroides fragilis(ETBF)の保菌者である32人の健康な日本人成人(平均年齢39.58±9.18歳)を対象とした別のオープンな無作為化並行群間試験でも、腸の微小環境に対するBB536のプラスの影響が明らかになりました(Odamaki et al。、 2012)。 この研究は、4週間の慣らし期間、8週間のコントロールミルクまたはBB536ヨーグルトの摂食、および12週間のウォッシュアウト期間で構成されていました。 BB536(108 CFU / g以上、1日あたり160 g)を含むヨーグルトの摂取は、健康な成人の腸内微生物叢のETBFの細胞数に識別可能な影響を与えました。 ETBFは、IBD患者の急性および持続性下痢性疾患(Sears、2009)および結腸直腸癌の発症(Orberg et al。、2017; Wu et al。、2009)と関連していることが示唆されています。 ベースライン値と比較して、8週目でのETBFの細胞数の有意な減少がBB536ヨーグルトを投与された被験者で観察されましたが、コントロールミルクグループでは観察されませんでした。 腸の微小環境を整えます。 その結果、これらの結果は、結腸直腸癌の予防と治療におけるBB536の潜在的な役割を暗示し、BB536がヒトの健康な微生物叢を維持できるという概念を支持しています。 |
| これに基づいて、腸環境の改善は、胃腸障害の緩和におけるBB536の臨床的有効性の主な理由の1つである可能性があることに注意することが不可欠です。 BB536の摂取は有益なビフィズス菌の成長を刺激し、糞便の腐敗性物質のレベルを低下させ、それによって便秘患者の排便問題を改善できることが、人間の介入研究の多くで示されています(Kondo et al.、2013; Ogata et al.、1997、1999; 八重島他、1997)。 たとえば、非ランダム化比較試験では、BB536(2×109 CFUおよび2×1010 CFU / 200 mL)を添加した牛乳を7人の1日1回、12人の健康な日本人成人(21〜57歳)に投与しました。 糞便中のアンモニア含有量とベータグルクロニダーゼの活性が大幅に減少する一方で、ビフィドバクテリウムの相対割合は大幅に増加しました(Ogata et al。、1997)。 腸内感染と腸疾患の原因となる病原菌である腸内細菌とクロストリジウムパーフリンジェンスの数は、BB536を添加した牛乳を摂取した被験者では減少する傾向がありました(Ogata et al。、1997)。 |
| さらに、BB536の腸の健康促進特性は、系統特異的であると思われます。 ビフィズス菌発酵乳のヒト腸内微生物叢への影響を調査した研究では、BB536が他のものよりもかなり優れていることが示されました(Ballongue&Grill、1993)。 非ランダム化比較試験では、BB536発酵乳(107 CFU / g、125 g)を1日に3回3週間投与すると、45人の被験者の腸内微生物バランスが大幅に改善され、ビフィドバクテリウムの量が著しく増加し、腐敗菌のレベルは減少しました。 それにもかかわらず、これらの変化は、B.アニマリスおよびB.ロンガム亜種ロンガムの他のすべての株(107 CFU / g以上)を含む発酵乳を同じ期間摂取した被験者では観察されませんでした(Ballongue&Grill、1993) 。 これらの不一致は、ヒトの腸の微小環境の改善におけるBB536の系統特異的および宿主特異的効果をさらに強調するのに役立ちます。 総合すると、これらの臨床試験は、より良い健康のために微生物のバランスを維持するというBB536の健康上の利点の証拠を提供します(表2)。 |
| 表2 腸の微小環境に対するBB536の影響の臨床研究からの要約 |
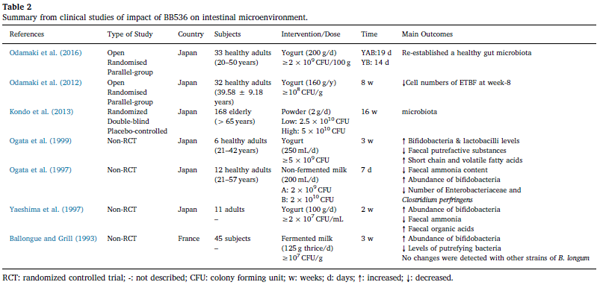 |
| RCT:ランダム化比較試験; -:説明されていません。 CFU:コロニー形成単位; w:週; d:日; ↑:増加しました。 ↓:減少しました。 |
| 3.3 免疫調節 |
|
プロバイオティクスは免疫機能を調節できるため、宿主免疫系が関与する特定の病態を予防または緩和するための興味深い代替手段として適用できることが長い間実証されています(Yan&Polk、2011)。 それにもかかわらず、密接に関連するプロバイオティクス株でさえ、免疫系に異なる影響を与える可能性があることが明らかになりつつあります(Klaenhammer、Kleerebezem、Kopp、およびRescigno、2012年)。 プロバイオティクス細菌は、異なるサイトカイン応答を誘発する可能性があり、異なるビフィドバクテリウム株は、明確で反対の免疫応答さえ誘導する可能性があります(He et al。、2002)。 言い換えれば、生物学的に関連するプロバイオティクス株の微調整された選択は、宿主免疫の調節に必要です。 BB536は、免疫系へのプラスの効果に関する強力な科学的証拠を持つ定評のあるプロバイオティクス株の1つです。 BB536の補給は、免疫調節機能によって健康上の利点をもたらすことが示されており、免疫介在性疾患の予防および治療の可能性への応用について臨床的に評価されています(表3)。 |
| 表3 BB536の免疫調節効果の臨床および動物研究からの要約 |
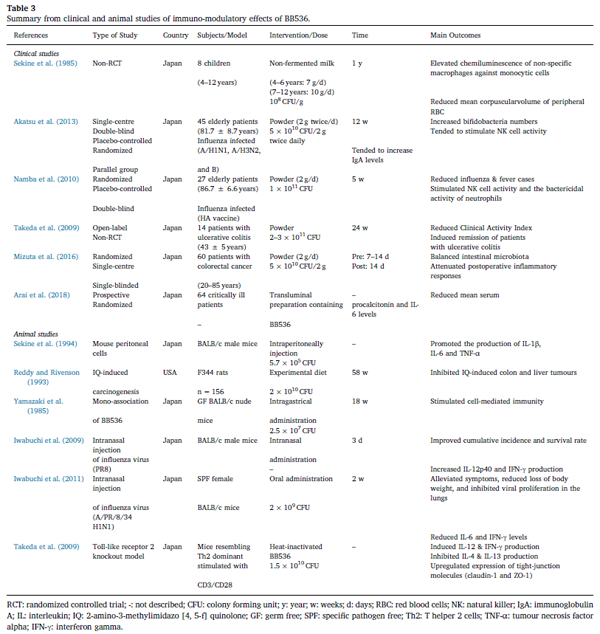 |
| RCT:ランダム化比較試験; -:説明されていません。 CFU:コロニー形成単位; y:年; w:週; d:日; RBC:赤血球; NK:ナチュラルキラー; IgA:免疫グロブリン A; IL:インターロイキン; IQ:2-アミノ-3-メチルイミダゾ[4、5-f]キノロン; GF:無菌; SPF:特定病原体除去; Th2:Tヘルパー2細胞; TNF-α:腫瘍壊死因子 アルファ; IFN-γ:インターフェロンガンマ。 |
初期の研究では、通常の抗腫瘍化学療法に加えて、1年間白血病に苦しむ日本人の子供(4〜12歳)8人(4〜12歳)にBB536(108 CFU / g、1日あたり7〜10g)を与えると、 単球細胞に対する非特異的マクロファージの化学発光および末梢赤血球の平均赤血球容積の減少は、マクロファージ媒介抗腫瘍免疫に対するBB536の免疫刺激効果を示唆しています(関根、吉原、本間、平山、および戸塚、1985)。 いくつかのin vivo動物研究は、BB536が癌に対する宿主免疫を強化し、それによってBB536がマウス腹腔細胞における炎症性インターロイキン(IL-1βおよびIL-6)および腫瘍抑制サイトカイン(TNF-α)の産生を促進することも実証しました(関根、川島 、および橋本、1994)、 F344ラットで2-アミノ-3-メチルイミダゾ[4、5-f]キノロン(IQ)誘発結腸および肝臓の発癌を阻害し(Reddy&Rivenson、1993)、無菌ヌードマウスの単細胞で細胞性免疫を刺激しましたことがBB536に関連付けられています(Yamazaki et al。、1985)。 |
BB536の免疫刺激効果は、インフルエンザウイルス感染のモデルでも実証されています。 前臨床研究により、インフルエンザウイルスに感染したマウスへのBB536の鼻腔内および経口投与は、宿主細胞の免疫を強化し、インフルエンザ様症状を改善し、インフルエンザウイルス感染から保護されたことが示されています(Iwabuchi et al。、2009; Iwabuchi、Xiao、Yaeshima、&Iwatsuki 、2011)。さらなる臨床研究により、BB536は高齢者の衰弱性免疫を改善できることが示されています。 |
2つの異なる臨床試験でインフルエンザワクチン接種を受けた高齢者にBB536を投与すると(表3の詳細を参照)、好中球の貪食活性とナチュラルキラー(NK)細胞の活性を刺激し、インフルエンザと発熱の発生率を低下させる傾向があり、高齢者の免疫グロブリンA(IgA)のレベルを増加させます(赤津ら、2013;難波、波多野、八重島、高瀬、鈴木、2010)。 細胞性免疫反応は高齢者では弱く、免疫系機能の低下がワクチンの有効性の低下とインフルエンザウイルス感染のリスク増加に寄与することが示唆されています。 これらの結果は、BB536が病原性ウイルスに対する高齢消費者の抵抗力を高め、高齢者のインフルエンザワクチンに対する免疫応答を改善する潜在的なアジュバントとして適用できることを示唆しています。 |
BB536は、がんやインフルエンザウイルス感染に対する宿主の免疫を増強するだけでなく、免疫バランスを回復させ、炎症を抑制することにも注目できるのは興味深いことです。 非盲検パイロット試験により、BB536(2–3×1011 CFU)の24週間投与が潰瘍性大腸炎患者の寛解の誘導に有効であることが明らかになりました(n = 14;平均年齢43±5歳)(Takeda et al。、 2009)。 |
| マウスモデルを用いたさらなる調査により、脾細胞のThelper type 1(Th1)-支配的なサイトカインプロファイルの変化とタイトジャンクション分子のアップレギュレーションによる粘膜バリア機能の増強の両方が、潰瘍性大腸炎患者の炎症状態を抑制するBB536の可能なメカニズムであることが示されました(Takeda et al。、2009)。 その後、結腸直腸手術の術後合併症の軽減におけるBB536の有効性が、無作為化された単一中心および単一盲検の臨床試験で評価されました(Mizuta et al。、2016)。 大腸切除(20〜85歳)を受けた日本人患者60人を無作為化して、BB536(5×1010 CFU / 2 g)またはコントロールを1日1回、術前7〜14日間および術後14日間投与した。 BB536の介入は、結腸直腸手術を受けている患者の全身術後炎症反応を軽減し、血液学的および栄養状態の回復を改善し、BB536の投与が結腸直腸切除後健康な回復を促進できることを示唆しました(水田ら、2016)。 |
| より最近では、BB536は敗血症性合併症を予防できることが報告されており、非感染性疾患の重症患者に有益である可能性があります(新井、三島、太田、幸岡、松本、2018)。 敗血症は一般的に致命的な疾患であり、炎症反応の誇張として特徴付けられます(Nee、2006)。 |
| 前向き無作為化試験には、コントロールとして従来の治療のみを受けているか、またはBB536を含む経管製剤を使用している重症患者64人が関与しました。 重症患者の敗血症の特異的マーカーであるプロカルシトニンとIL-6のアップレギュレーションは、BB536を投与された患者で有意に抑制されることが実証されましたが、その効果は抗生物質治療によって打ち消されました(Arai et al。、2018)。 この研究は、BB536が重病患者の敗血症に起因する合併症を予防でき、広域抗生物質の使用が腸内微生物叢でのBB536の定着とその有益な効果を阻害した可能性があることを文書化しています。 |
| 総合すると、これらの研究はBB536の宿主免疫応答を調節する能力を実証し、臨床研究で得られた有望な結果はBB536の健康上の利点をサポートする科学的証拠を強化しました。 |
| 3.4 抗アレルギー |
| スギ花粉によって引き起こされる季節性アレルギー性鼻炎は、日本で最も一般的なアレルギー疾患の1つであり、国民の苦痛と考えられています(Yamada、Saito、&Fujieda、2014)。 スギ花粉症(JCPsis)は、刺激性のスギ花粉への曝露によって引き起こされる免疫グロブリンE(IgE)を介したI型アレルギーです。 JCPsisの有病率は過去20年間で劇的に増加しており、現在日本の人口の約30%に影響を及ぼしています(Fujimura&Kawamoto、2015)。 増えつつある証拠は、プロバイオティクスが宿主の免疫系を調節し、アレルギー疾患の症状を緩和する可能性があることを示唆しています(Yang、Liu、&Yang、2013)。 過去10年間に、プロバイオティクスのいくつかの株がJCPsisの治療のために提唱されてきました(石田ら、2005;川瀬ら、2009;田村ら、2007;シャオら、2006a)。 それにもかかわらず、これらのプロバイオティクス株の多くの臨床効果は詳細に調査されておらず、決定的ではありません。 たとえば、アレルギー性鼻炎の症状を改善するためのプロバイオティクス乳酸菌の影響に関する2つの研究のデータは矛盾しています(Helin、Haahtela、&Haahtela、2002; Tamura et al。、2007)。 これらの研究は、アレルギー性疾患に対するプロバイオティクスの有益な効果は、使用する株と治療のタイミングに依存することを強調しています。 |
| BB536は、JCPsisの治療に関して広範に研究されており、実質的な臨床データと再現性のある結果が得られています(Xiao et al。、2006a、2006b; Xiao、Kondo、Takahashi、et al。、2007; Xiao、Kondo、Yanagisawa、et al。、2007 )。 BB536ヨーグルトまたは凍結乾燥粉末の摂取は、スギ花粉に敏感な患者の鼻および眼のアレルギー症状を潜在的に緩和し、アレルギー性免疫反応を調節することが示されています。 これは過去10年間で最も重い季節である2005年の花粉シーズン中に13週間、1日2回、5×1010 CFU / 2 gの用量でBB536粉末を摂取した44人のJCPsis被験者(26〜57歳)を行った注目すべき無作為化二重盲検プラセボ対照試験は、JCPsisに関連するすべての症状の著しい改善を示し、免疫機能を改善する傾向がありました(Xiao et al。、2006b)。 さらに、BB536は、花粉の分散と一緒に発生したTヘルパー2型(Th2)スキュー免疫反応を大幅に改善し、それにより、胸腺および活性化調節ケモカイン(TARC)の血漿レベルがBB536粉末を投与された被験者で著しく正常化されました(Xiao et al。 、2006b; Xiao、Kondo、Takahashi、et al。、2007)。 花粉シーズン中の血漿TARCレベルの上昇は、JCPsisの病気の重症度と正の相関があることが注目されました。 さらに、BB536の補充はアレルギー反応を改善する可能性があり、それによりスギ花粉特異的IgEおよび好酸球のレベルの低下とインターフェロンガンマ(IFN-γ)のレベルの増加が4週目のベースラインからBB536グループで検出されました(Xiao et al。、 2006a)。 これらの研究は、スギ花粉に敏感な被験者のアレルギー反応の重要なマーカーに対するBB536消費のプラスの影響を一貫して示しています。 |
| 別の二重盲検、プラセボ対照、無作為化試験により40人のイタリアの子供(4〜17歳)が3つのビフィズス菌菌株BB536(3×109 CFU)、B.longum subsp. infantis M-63(1×109 CFU)、およびB. breve M-16V(1×109 CFU)も4週間にわたって、花粉誘発性アレルギー性鼻炎の有意な改善を明らかにしました(Del Giudice et al.,017)。 プロバイオティクス混合物の投与は、花粉誘発性アレルギー反応から子供を保護し、生活の質を改善しました。これらのパラメーターはプラセボ群で悪化しました。 この研究は、BB536が他のビフィドバクテリウム株と組み合わされた場合、顕著な有益な効果を持つことを示唆しています。 |
| さらに、アレルギー性鼻炎に対するBB536の優れた保護効果は、主要なカバおよび草の花粉アレルゲンに対する多感作のマウスモデルを使用したin vivo試験で実証されています(Schabussova et al。、2011)。 2つのプロバイオティクス株であるL. paracasei NCC2461およびBB536(研究ではB. longum NCC3001と表示)の異なる時点で適用された5 x 108 CFUの用量での免疫抑制特性を比較しました。 両方のプロバイオティクス株による治療は、アレルゲン特異的免疫応答を著しく抑制しました。 ただし、感作とチャレンジの前に適用すると、BB536のみが長期持続的な保護効果を示し、BB536はアレルギー性鼻炎の予防と治療の両方に適用できることを示しています。 総称して、臨床および動物研究からの発見は、プロバイオティクス株選択性の概念を支持し、アレルギー状態および免疫応答の改善におけるBB536の顕著な有益な効果を指摘しました(表4)。 |
| 表4 アレルギー性疾患に対するBB536の効果に関する臨床および動物研究からの要約 | 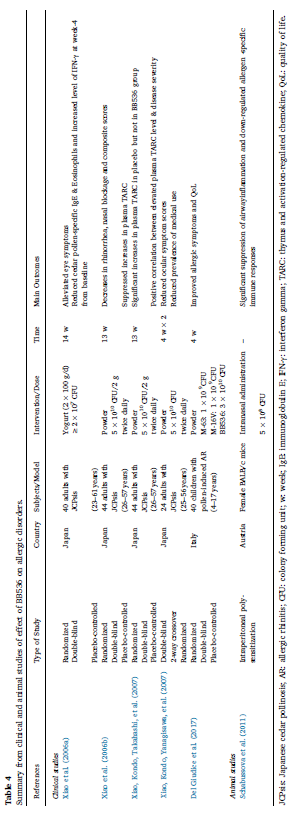 |
| JCPsis:スギ花粉症; AR:アレルギー性鼻炎; CFU:コロニー形成単位; w:週; IgE:免疫グロブリンE; IFN-γ:インターフェロンガンマ; TARC:胸腺および活性化調節ケモカイン; QoL:生活の質。 |
| 3.5 抗感染症 |
| BB536は、微生物感染の治療と制御にも有効であることが示されています。 いくつかのin vitroおよび動物研究は、BB536がグラム陰性病原体(例えば、大腸菌、クレブシエラ肺炎、C perfringes、Ba fragilis、Salmonella typhi、およびPseudomonas aeruginosa)の成長を阻害できることを実証しています(Araya-Kojima、Yaeshima、Ishibashi、島村、早澤、1995;イントゥリ、スティバラ、フルネリ、ブランディーノ、2016;松本ら、2008;難波、八重島、石橋、早沢、山崎、2003;山崎、上村、百瀬、川島、上田、1982 )。 |
| 競争的排除は、病原性感染に対するBB536の有効性の主な理由の1つです。 BB536は、大腸菌、腸炎菌、および腸チフス菌(Inturri et al。、2016)および緑膿菌(松本et al。、2008)に感染したマウスモデルの上皮細胞への病原性細菌の付着を妨害します。 また、BB536は、大腸菌O157:H7感染症のBB536単一関連無菌マウスで実証されているように、乳酸や酢酸などの代謝物の放出を介して病原菌の致死活性を阻害する可能性があります(Namba et al。、2003)。 |
| BB536の抗感染活性を文書化したこれらの結果は、いくつかの臨床研究(Chitapanarux、Thongsawat、Pisespongsa、Leerapun、およびKijdamrongthum、2015年; Kageyama、Tomoda、&Nakano、1984; Lau et al。、2018; Tomoda、Nakano、&影山、1988)に表されています。 BB536の投与により、白血病患者のカンジダの過成長が予防されたことが報告されました(Kageyama et al。、1984; Tomoda et al。、1988)。 侵襲性真菌感染は、白血病患者の罹患率と死亡率の主な原因であり、腸内微生物叢の摂動とカンジダの過成長は、抗白血病化学療法を受けている患者の注目すべき現象です(Bhatt、Viola、&Ferrajoli、2011)。 驚くべきことに、BB536(107 CFU / mL、1日あたり200 mL)を3か月間投与すると、56人の患者の腸内バランスが回復し、血漿内毒素(既知の毒性因子)および尿中インディカン(腸内腐敗の尺度)のレベルが正常化されました( 20〜60歳)抗白血病療法を受けている(影山ら、1984)。 データは、BB536が腸内微生物叢の自然なバランスを回復し、日和見病原菌の侵入を防ぐことができるという概念をサポートしています。 |
| ヘリコバクターピロリ感染に対するBB536の効果を調べる別の無作為化二重盲検プラセボ対照試験では、標準的なトリプルセラピー(エソメプラゾール、アモキシシリン、クラリスロマイシン)と組み合わせたBB536の1日2回投与が63人の患者のピロリ感染(平均年齢52.61±11.30)のヘリコバクターピロリの根絶率を改善することが明らかになりました 。(Chitapanarux et al。、2015)。 ピロリ菌は慢性胃炎の主な原因であり、胃癌の重要な危険因子と考えられています(上村ら、2001)。 H. pyloriを根絶するために、抗生物質療法が一般的に推奨されます。 ただし、このようなレジメンは、コンプライアンスの低下、副作用、耐性の出現に関連することがよくあります(Deltenre、Ntounda、Jonas、&De、1998)。 特に、BB536の補給は、プラセボ対照と比較して、非潰瘍性消化不良患者の抗ピロリ抗生物質療法関連下痢を有意に改善しました(Chitapanarux et al。、2015)。 いくつかのランダム化比較試験は、プロバイオティクス治療が合併症を伴うピロリ菌治療を減らすことができるようであることを示しましたが、ピロリ菌の根絶率の有意な改善はほとんどの研究で検出されず、ピロリ菌の増殖に対するプロバイオティクスの阻害活性が非常に株特異的であることを示唆しています( Lionetti et al。、2010)。 したがって、これらのデータは、BB536がH. pylori感染と戦って抗生物質関連の合併症から保護するのに優れていることを示しています。 |
| より最近では、BB536は、2〜6歳の健康な就学前児童の上部呼吸器感染を改善できることが報告されました(Lau et al。、2018)。 この研究は、マレーシアの上気道疾患の流行しやすい季節の間に、BB536(5×109 CFU / g; n = 109)またはプラセボ(n = 110)の凍結乾燥粉末を10か月間投与した219人の被験者を対象とした無作為化二重盲検並行並行プラセボ対照研究でした。 BB536は、呼吸器疾患の発生から子供を保護する際に、より一般的な抗感染効果を発揮し、BB536を投与された子供では、プラセボ群と比較して咽頭痛の持続時間が著しく改善されたことが注目されました。 一方、BB536を補充した子供は、プラセボ対照と比較して、他の症状(発熱、鼻水、咳)の日数もわずかに減少しました。 総合すると、これらの臨床研究は、BB536がヒトのグラム陰性病原性およびウイルス感染症に対する有望な抗感染剤として使用できることを示唆しています(表5)。 |
| 表5 さまざまな感染症に対するBB536の効果の動物および臨床研究からの要約 | 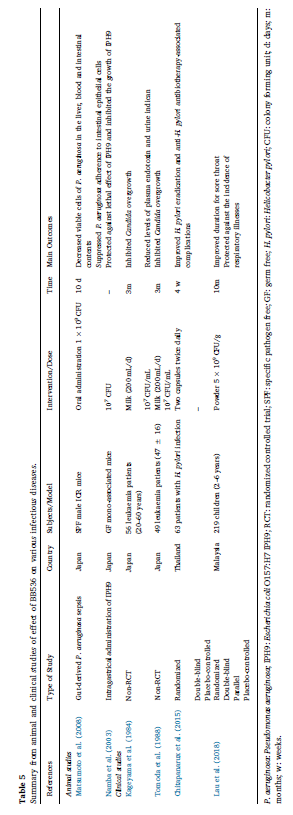 |
| 緑膿菌:緑膿菌; IPH9:大腸菌O157:H7 IPH9; RCT:ランダム化比較試験; SPF:特定病原体除去; GF:無菌; H.ピロリ:ヘリコバクターピロリ; CFU:コロニー形成単位; d:日; m:月; w:数週間。 |
4. BB536の主要な作用としての腸内微生物叢の変調 |
| 40年以上にわたる研究により、BB536は人間に複数の健康上の利点を与えることが明らかになり、その結果はBB536の多機能活動が主にBB536と宿主の腸内微生物叢との相互作用に起因することを示唆しました(図1)。 |
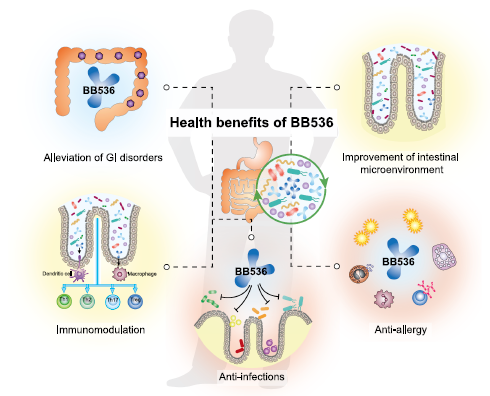 |
| 図1.腸内微生物叢の調節は、ビフィドバクテリウムロンガム亜種の主要な有益な作用です。 longum BB536は、人間の健康を促進します。 BB536は、腸内微生物叢と連携して、胃腸の健康を改善し、宿主の免疫恒常性を調節し、アレルギー障害と感染状態を緩和します。 |
| 4.1。 腸内微生物代謝に関する微生物クロストーク |
| BB536の影響の根底にある分子基盤を解明する試みにおいて、優勢なヒト腸由来微生物叢(HGM)の15系統を保有するノトバイオティクスマウスが採用されました(Sugahara et al。、2015)。 ヒト化ノトバイオティクスマウスモデルは、プロバイオティクス系統が腸内環境を改善するメカニズムを研究する可能性を提供する強力なツールです。 |
| 個々の腸内微生物叢の変動を含む、人間の研究における交絡因子をよりよく制御します。 また、マウスモデルの腸内微生物叢の組成はヒトのものとは大きく異なるため、腸内微生物叢の研究結果を従来のマウスモデルからヒトに翻訳する際に落とし穴がある可能性があることにも留意されたい。 この目的のために、HGM関連のノトバイオティクスマウスにBB536またはPBSを毎日14日間投与し、BB536の腸環境への影響を、メタボローム、メタゲノム、およびメタトランスクリプトーム分析などの多面的なアプローチを使用して評価しました(Sugahara et al。、2015 )。 ビオチンの前駆体である酪酸およびピメレートの糞便レベルの有意な増加がBB536-HGMマウスで観察されたが、BB536-モノ関連マウスでは観察されず、BB536はHGMとの微生物クロストークを通じて腸代謝を調節できることを示唆している。 さらに、ピメレート代謝を通じて進行するビオチン合成経路に関与するBacteroides caccaeの細菌転写産物は、HGMグループよりもBB536-HGMグループで有意に高く、BB caccaeとの相互作用によりBB536がビオチン生合成を調節したことを示しています(菅原ら 。、2015)。 BB536は前駆体ピメレートの生成を促進し、Ba caccaeがそれをさらにビオチンに代謝することを可能にし、それにより宿主の腸の恒常性に貢献しました(図2A)。 |
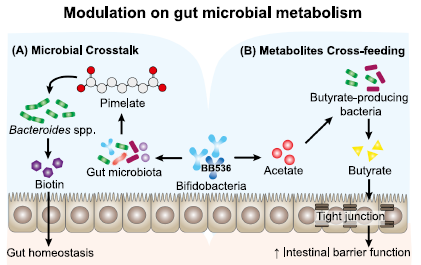 |
| 図2. Bifidobacterium longum subsp longum BB536による、ヒト腸内微生物叢との微生物クロストークによる腸代謝の調節。 |
(A)BB536は、前駆体ピメレートの生成を促進することによりビオチン生合成を調節し、バクテロイデスカッカエがそれをさらにビオチンに代謝することを可能にし、それにより宿主の腸の恒常性に寄与する。 (B)BB536は、クロスフィードメカニズムを介して、共生酪酸産生細菌(例:Eubacterium rectale)の代謝活性に影響を与えます。 炭水化物発酵でBB536によって生成されるアセテートは、Eu.rectaleの成長を維持する基質として機能し、酪酸塩の生成を刺激します。 |
| 4.2 腸内微生物叢のバランスの回復 |
| 腸内細菌叢の多様性とバランスは、免疫機能、代謝機能、神経機能を含む人間の健康の多くの側面と密接に関連していることが示唆されています(Valdes、Walter、Segal、&Spector、2018)。 腸内細菌叢の個体数の変動は、多くの健康障害と強く関連しています。 例えば、腸内微生物叢の多様性の喪失と不均衡、特にラクトバチルス属、ビフィドバクテリウム属、アッカーマンシア・ムニシフィラおよびフェカリバクテリウム・プラウスニツィイなどの有益な生物のレベルの低下は、胃腸障害、アレルギー、メタボリックシンドロームおよび慢性疾患などのリスクをもたらす可能性があります(Clemente、Ursell、Parfrey、Knight、2012)。 注目すべきは、BB536のプロバイオティクス介入が腸内微生物叢の有益な分類群の豊富さを促進し、さまざまな健康上の合併症を有する被験者のビフィズス菌の跋扈(有病率]を刺激することができることは明らかです(Akatsu et al。、2013; Ballongue&Grill、1993;水田 et al。、2016; Ogata et al。、1997)。 BB536の介入により刺激されるビフィズス菌の豊富さおよびおそらく関連する活性の増加は、微生物叢の組成に対する選択的な圧力として作用する可能性があります。 これに基づいて、BB536は腸内微生物叢のバランスを回復することにより、人間の宿主にプラスの効果を発揮すると考えられています。 |
| BB536の微生物恒常性への影響の顕著な例は、動物ベースの食事摂取の人間の試験で示されています(Odamaki et al。、2016)。 動物ベースの食事によって引き起こされる腸内微生物叢の変動、特にBilophila wadsworthiaの量の増加は、BB536ヨーグルトの摂取によりバランスが取り直されました(Odamaki et al。、2016)。 Bil wadsworthiaは、虫垂炎患者から一般に回収されるグラム陰性亜硫酸塩還元および硫化水素産生細菌です(Bernard、Verschraegen、Claeys、Lauwers、およびRosseel、1994)。 Bil wadsworthiaの存在量が増加すると、腸の炎症、腸のバリア欠損、胆汁酸代謝異常、および微生物叢の機能プロファイルの変化が促進される可能性があることが報告されました(Natividad et al。、2018)。 Bil wadsworthiaの過剰表現は、動物ベースの食事に関連していることが判明しました(David et al。、2014)。 飽和動物由来脂肪からのタウリン共役胆汁酸の産生の増加は、Bil wadsworthiaなどの亜硫酸還元細菌の拡大を刺激する有機硫黄の利用可能性の向上に寄与すると推測されます(Devkota et al。、2012)。 驚くべきことに、BB536はBil wadsworthiaの成長を抑制し、腸内微生物叢の変動を減衰させるのに効率的でした(Odamaki et al。、2016)。 BB536のプロバイオティクス介入が動物食による有害な影響から保護し、人間の健康を改善する方法は未解決の問題ですが、Bil wadsworthiaの拡大を制限し、腸内微生物叢のバランスを回復することにより、BB536が多くの健康障害を改善する可能性があります。 |
| 総合すると、これらのデータは、不均衡状態にある腸の微生物バランスの回復(腸の共生)が、プロバイオティクスBB536が微生物の健康を促進し、人間の健康を改善する1つのメカニズムである可能性を示唆しています。 |
| 4.3 免疫恒常性 |
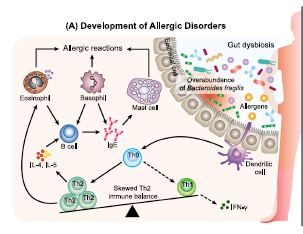
| 免疫恒常性の維持は、宿主の生存に不可欠です。 腸内微生物叢は、宿主の免疫系の重要な調節因子です(Belkaid&Hand、2014)。 腸内細菌叢のバランスの変化は、腸内細菌叢異常と呼ばれる状態であり、異常な免疫反応を特徴とする多くの疾患(炎症性疾患やアレルギー性疾患など)と密接に関連しています(Kosiewicz、Dryden、Chhabra、およびAlard、2014年)。 最近の研究により、腸内細菌叢と特定のT細胞サブタイプ(Th1、Th2、Th17および制御性T(Treg)細胞)の発生との関連が明らかになっています(Lee&Kim、2017)。 腸内微生物叢の異常は、歪んだT細胞応答とアレルギー疾患の誘発に密接に関連していることが明らかになりました(Hong、Kim、&Surh、2017)。 たとえば、季節性アレルギー性鼻炎、JCPsisは、スギ花粉への曝露がアレルギー被験者の腸内微生物叢の変動に寄与するが、健康な被験者ではないという観察によって示されるように、腸の片利共生に対する耐性の喪失によって特徴付けられることがわかった。 特に、Ba fragilisの存在量は、JCPsis被験者の微生物コンソーシアム内で大幅に増加しました(Odamaki et al。、2007a、2007b)。 Ba fragilisの有病率のこのような増加は、スギ花粉に特異的なIgEのレベルと正の相関があり(Odamaki et al。、2008)、腸内微生物叢の障害が宿主免疫の混乱とアレルギー性疾患の発症に寄与する可能性があることを示唆しています(図。 3A)。 |
| 注目すべきことに、変動した腸内細菌叢、特にBa fragilisの過剰は、BB536を投与されたJCPsis被験者で大部分が修正されました(Odamaki et al。、2007b)。 JCPsis被験者からの末梢血単核細胞を使用したさらなるin vitro研究では、Ba fragilisがT細胞関連サイトカインを誘導する能力を持っていることが示されました。 Ba fragilisの株は、ビフィズス菌よりも高レベルのTh2サイトカイン(IL-6)および低レベルのTh1サイトカイン(IL-12およびIFN-γ)を有意に刺激することがわかりました(Odamaki et al。、2007a)。 この結果は、Ba fragilisの存在量とアレルギー性疾患の発症におけるTh2の歪んだ免疫バランスとの関連についての理解への道を開く可能性があります。 BB536の補給は腸内微生物叢の安定化を促進し、ビフィズス菌の量を増やし、その結果、Th1 / Th2バランスを回復し、アレルギー症状を軽減することが示唆されています(図3B(i))(Odamaki et al。、2007a; Xiao et al 。、2006b; Xiao、Kondo、Takahashi、et al。、2007)。 さらに、BB536はTh2の歪んだ免疫バランスに直接影響することも報告されています(図3B(ii))。 in vitroおよびin vivoの両方の研究は、BB536のゲノムDNAから同定された新規免疫刺激配列オリゴデオキシヌクレオチド(ODN)BL07SがIgE産生を阻害することを示しました(Takahashi et al。、2006a、2006b、2006c)。 卵白アルブミン感作マウスモデルを用いたさらなる研究により、BB536の免疫刺激性ODN BL07SはTh2サイトカイン産生(IL-4およびIL-5)を有意に抑制し、脾細胞培養におけるTh1サイトカイン(IFN-γ)のレベルを増加させ、BB536が抗原を妨げたことを示した Th2は、細菌成分を介して免疫応答を歪めました(Takahashi et al。、2006a、2006b)。 総合すると、これらの発見は、BB536が花粉感作の有病率を低下させ、アレルギー症状を緩和するメカニズムを明らかにしました。 BB536は、ホストとミクロビオームの相互作用内で微調整された恒常性バランスを駆動することにより、免疫機能障害を改善したことは明らかです。 |
 |
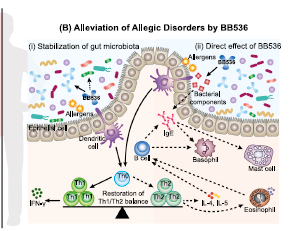 |
図3. Bifidobacterium longum subsp longum BB536の免疫調節効果 |
(A)腸内微生物叢の変動、特にBacteroides fragilisの過剰は、宿主免疫の摂動とアレルギー疾患の発症に寄与します。 アレルギー反応では、アレルゲンは樹状細胞に取り込まれ、ナイーブT細胞(Th0)に提示され、Tヘルパータイプ2(Th2)細胞に変換されます。 Th2細胞はインターロイキン(IL)-4およびIL-5を分泌し、その後メモリーB細胞を刺激して、免疫グロブリンE(IgE)抗体の産生が支配的なアレルゲン特異的体液性応答に切り替えます。 これらのIgE抗体はマスト細胞および好塩基球に付着し、それによりアレルギー性症状のその後の暴露および発症に感作します。 (B)BB536は、ホストとミクロビオームの相互作用内で免疫恒常性を調節し、間接的および直接的なメカニズムを介してアレルギー性疾患を緩和します。 (i)BB536は、Bacteroides fragilisの有病率を改善することにより腸内微生物叢の安定化を促進し、その結果、Th1 / Th2バランスを回復し、アレルギー症状を緩和します。 (ii)BB536は、その細菌成分を介して、抗原誘発性IgEを介したTh2歪んだ免疫バランスに直接影響を与えます。 実線の矢印線:刺激。 破線の矢印:抑制。 |
5 結論 |
| Bifidobacterium longum subsp longum BB536は、ヒト宿主に一貫した有益な健康効果をもたらすことができる臨床的に有効な実証済みのプロバイオティクス株の1つになりました。 |
BB536の明らかな利点の1つは、このプロバイオティクスが十分に特性化されたヒト起源の株であり、安全性と臨床効果の実績を持つ人間の健康に広く使用されていることです。 これらの有益な健康効果はBB536に固有のものではないかもしれませんが、BB536の機能的利点に関する証拠は、プロバイオティクスとして見事に市場に出されている多くの文書化されていない株とは対照的です。 |
| 動物研究と臨床研究の両方から得られた有望な結果は、まだ範囲が限られているにもかかわらず、人間の健康に対するBB536の影響の根底にある潜在的なメカニズムの概要を提供しています。 |
| 腸内マイクロバイオームの調節は、ヒトの腸におけるBB536の健康増進活動の重要な要素である可能性が高いことが明らかになりました。 BB536は、腸内細菌叢と連携して、宿主の恒常性を調節し、胃腸の健康を改善し、アレルギー性疾患を緩和します。 |
| これらの事実を考えると、BB536は人間の健康の治療と管理において有用で価値のあるプロバイオティクスの候補として役立つかもしれません。 BB536、腸内細菌叢および宿主間の相互作用が人間の健康にどのように影響するか、および人間のプロバイオティクスとしてのBB536の可能性を最大限に引き出すための分子的基礎についての新たな洞察を得るには、大規模で縦断的な統合的および機構的研究によるさらなる調査が重要です。 |
|
この文献は、Journal of Functional Foods Volume 54, March 2019, Pages 506-519に掲載されたBeneficial effects of Bifidobacterium longum subsp. longum BB536 on human health: Modulation of gut microbiome as the principal actionを日本語に訳したものです。タイトルをクリックして原論文の全文を英文で読むことが出来ます |