ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(ヴィーリ:研究の最前線) |
一酸化窒素およびインターロイキンは、ヴィーリエキソポリサッカライドによって活性化されるRAW264.7マクロファージの細胞増殖に関与しています |
Junhua Wu et al
|
|
抄録 |
ヴィーリは伝統的に健康食品と見なされてきました。 ヴィーリエキソポリサッカライド(VEPS)は抗酸化物質として機能しますが、分子および細胞のメカニズム、特にその免疫機能はほとんど不明のままです。 VEPSの免疫学的役割を評価するために、VEPSをSevageの方法で分離し、陰イオン交換クロマトグラフィーで精製しました。細胞増殖、食細胞、一酸化窒素(NO)、インターロイキン(IL)-1β、およびIL-6の放出、逆転写ポリメラーゼ連鎖反応(RT-PCR)およびiNOSタンパク質による誘導性一酸化窒素合成酵素(iNOS)遺伝子発現リポ多糖(LPS)/ VEPS刺激および非刺激RAW264.7マクロファージの走査型電子顕微鏡によるウエスタンブロット法および形態を分析しました。VEPSは50〜200μg/ mLで細胞増殖を増加させました。LPSおよびVEPSへの暴露後、貪食およびNO、IL-6、およびIL-1βの放出の徴候に対するニュートラルレッドの取り込みが増強されました。iNOS、IL-6、IL-1βの遺伝子発現とiNOSのタンパク質発現は、VEPSによって増加しました。VEPSで処理したRAW264.7細胞は平らになり、マクロファージの活性化を強く示しています。VEPSは、NO、IL-6、およびIL-1βが関与するマクロファージの活性化を促進すると結論付けました。 NOやその他のサイトカインの放出は、最終的にリンパ球を活性化し、ヒトの非特異的(自然)および特異的免疫を増加させる可能性があります。 |
はじめに |
|
フィンランド発祥の半固体ヨーグルトであるヴィーリは、乳酸菌(LAB)とベルベットのような表面を形成する表面成長菌Geotrichum candidumの微生物作用に起因する、ロープ状のゼラチン状の一貫性と酸味を持っています。さらに、ヴィーリには酵母:Kluveromyces marxianusおよびPichia fermentansも含まれています。中温性LAB株の中で、スライム形成LAB cremoris は、リン酸塩含有細胞外多糖(EPS)を生成します。ヴィーリの EPSの基本構造は、主にDグルコース、D-ガラクトース、L-ラムノース、リン酸塩で構成され、平均分子量は約2,000 KDaで、繰り返し単位は「→4-β-Glcp-(1→4)- β-D-Galp(1→4)-β-D-Glcp-(1→”、および-L-Rhapと-DGalp- 1-pのグループ Galp [1、2]の両側に取り付けられています。抗酸化、抗炎症、抗癌、抗老化、自然免疫の強化を含むさまざまな機能上の利点があると言われています[3-6]。 |
VEPSはRAW264.7の細胞増殖を増加させました |
RAW264.7細胞に対するVEPSの細胞毒性効果を調べて、細胞増殖、NO放出、食作用、iNOS遺伝子発現、および細胞活性化の形態の分析のためのVEPS治療の適切な濃度範囲を確立しました。MTTアッセイにより、VEPSの濃度が50〜200μg/ mLの範囲内で、適切な濃度8 x 104 cell / mLで非細胞毒性が観察されました。VEPSはRAW264.7細胞増殖を有意に増加させ、LPSはRAW264.7マクロファージ細胞増殖を最大3倍刺激しました(図1)。 相乗効果も一緒に見られました(データは示していません)。 MTTの結果は、顕微鏡下での細胞数と一致していました。
|
|
| 図1. VEPSの異なる濃度での細胞増殖分析、LPSとコントロールの比較。 結果は平均値±標準偏差(n05)を表しています。 **対照群と比較して、p <0.01および* p <0.05。 |
VEPSはRAW264.7マクロファージ細胞の食作用を増加させました |
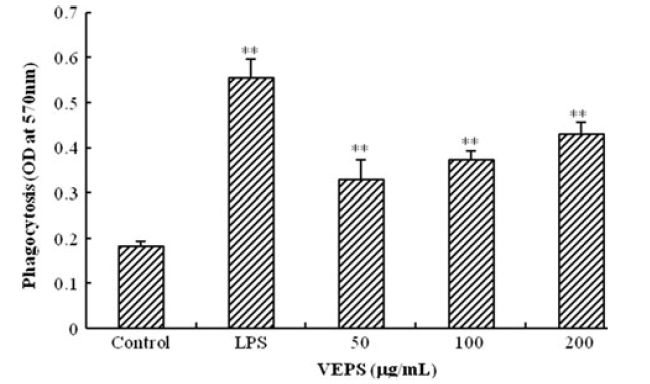
RAW264.7の貪食活性は、中性フクシン(赤)取り込み法によって測定されました。VEPSの最終濃度が50から200μg/ mLに増加すると、OD値が増加し、VEPSがRAW264.7マクロファージの食作用を増加させることが示されました。
|
|
| 図2. VEPSの影響を受けたRAW264.7細胞の食作用とLPSとコントロールの比較。 ニュートラルレッドによる食作用後の吸光度は570 nmで記録されました。 **対照群と比較してp <0.01。 |
RAW264.7細胞でのVEPS誘発NO産生p>
|
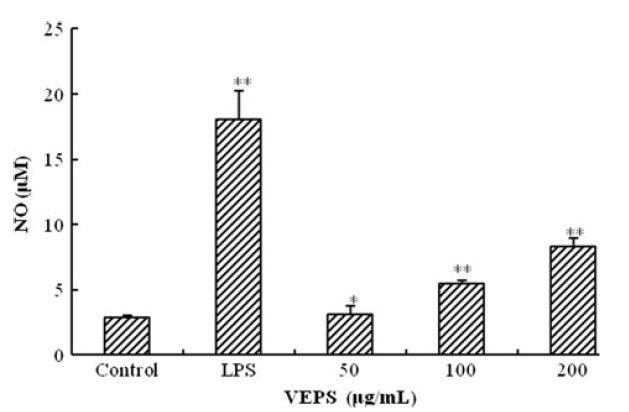
対照と比較したRAW264.7細胞のNO産生に対するVEPSの効果は、培地に放出された亜硝酸塩の量を測定することにより評価されました。NO産生がVEPSによって調節されたかどうかを調べるために、上清のグリースアッセイによってNO産生を定量化しました。NOは、刺激されていないRAW264.7細胞では非常に低かったが、VEPSでは適度に増加し(図3)、LPSでは大幅に増加した。 |
|
図3. VEPSの影響を受ける一酸化窒素(NO)と、グリース法を使用して測定されたLPSとコントロールの比較。 |
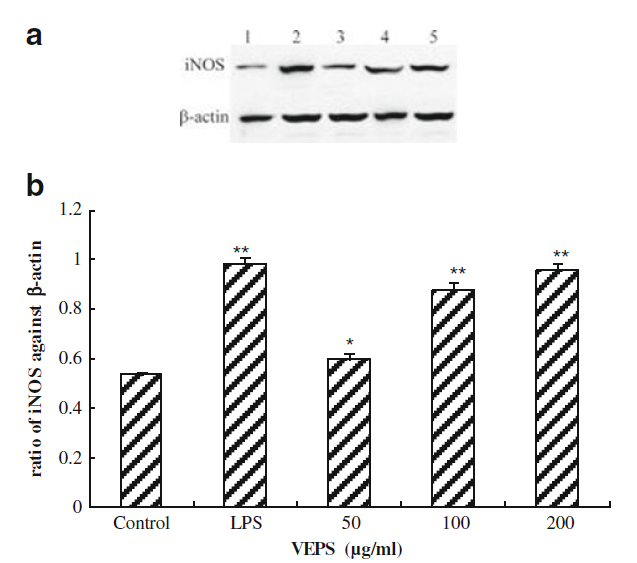
VEPSはiNOSの遺伝子およびタンパク質発現を増加させました
|
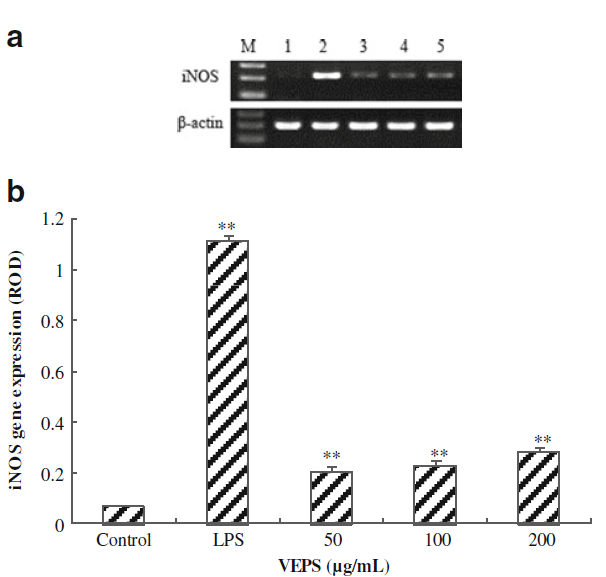
ハウスキーピング遺伝子であるβ-アクチンを差し引いたRT-PCRで示されるように、LPSとVEPSは明らかにiNOSを誘導しました。iNOSの遺伝子発現はVEPSによって増加しました(図4)。
|
|
図4。LPSおよびVEPSを含むマクロファージRAW264.7細胞におけるRT-PCRによるiNOSの遺伝子発現。 B ハウスキーピング遺伝子β-アクチンを収縮させた後、相対光学密度(ROD)をプロットしました。 **対照群と比較してp <0.05。
|
|
図5. VEPSの濃度が異なるマクロファージRAW264.7細胞でのウエスタンブロットによるiNOSのタンパク質発現。 |
VEPSはIL-6およびIL-1βの分泌と遺伝子発現を増加させました
|
ELISA法により検出された分泌インターロイキン、IL-6およびIL-1βは、LPSよりも弱いにもかかわらず、VEPSがIL-6およびIL-1βを増加させたことを示しています(図6)。
|
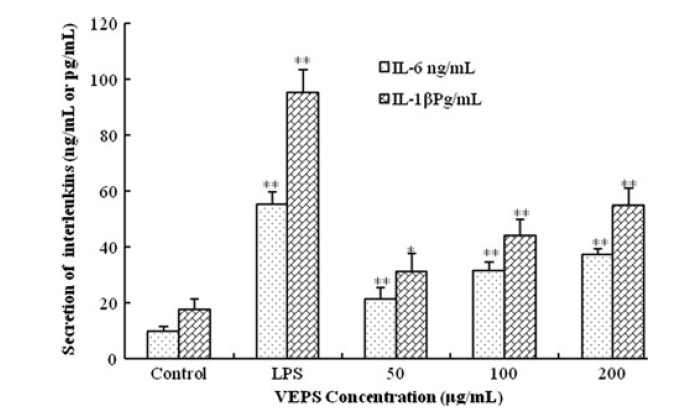 |
図6。50、100、および200μg/ mLのVEPSで刺激した後のRAW264.7細胞のIL-6およびIL-1βの分泌、LPSと対照の比較、
|
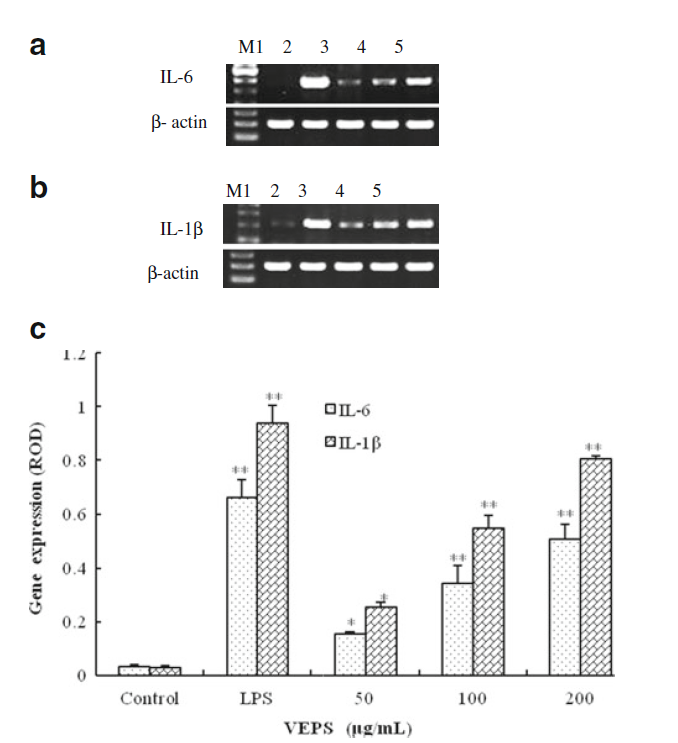 |
図7.マクロファージRAW264.7のインターロイキン遺伝子発現は、RT-PCRによりVEPSおよびLPSに影響を受けました。
|
VEPSによるRAW264.7の形態変化 |
マクロファージは通常、SEM観察下のコントロールと比較して、LPSおよびVEPSでの刺激後に平坦化されました(図8)。
|
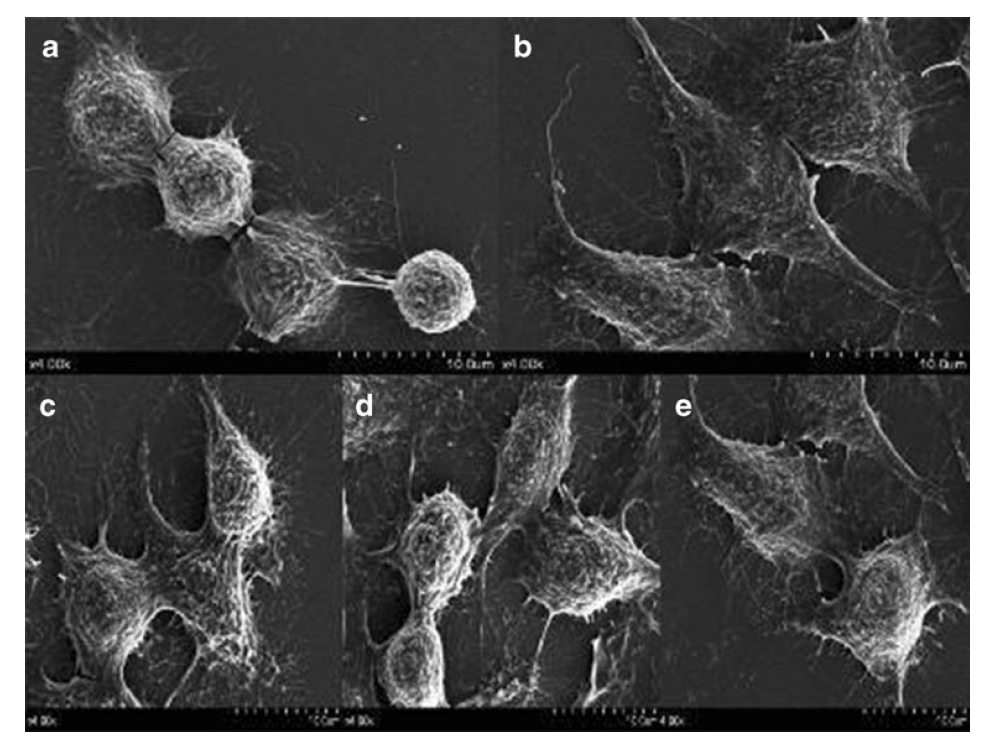 |
図8。SEMによるVEPSおよびLPSの刺激下でのRAW264.7の細胞形態。
|
討論 |
Lactococcus lactis subsp cremorisのVEPSは、2つのブタ腸管受容体(poRP105およびpoMD-1 [20、21])を介してNF-κBおよびさまざまなサイトカインを強く誘導することができました。これはヒトに非常に近いモデルであるマウスマクロファージRAW264.7細胞の調査に影響を与えました。マクロファージは通常、小腸に蓄積され、さまざまな化合物によって引き起こされる感染に対する宿主防御として、自然免疫および適応免疫において重要な役割を果たします。 非常に頻繁に、多糖類またはLPSが関与します。 私たちのアッセイでは、VEPSは50、100、および200μg/ mLの濃度でRAW264.7の細胞増殖を増加させる能力、また、食細胞NO分泌、iNOS遺伝子およびタンパク質の発現、分泌の著しい増加、私たちのアッセイでは、VEPSは50、100、および200μg/ mLの濃度でRAW264.7の細胞増殖、また、食細胞の著しい増加、NO分泌、iNOS遺伝子およびタンパク質の発現、分泌、IL-6およびIL-1βの遺伝子発現を増加させる能力を示しました。そして平坦化形態を示しました。陽性対照では、LPSはVEPSグループと比較してはるかに強力であることを常に示しています。 LPSの存在下でVEPSを使用した場合にも相乗効果が観察されました(データは示していません)。NOは炎症反応のメディエーターおよびレギュレーターとして認識されています。亜酸化窒素シンターゼには3つのアイソフォームがあるが、iNOSはNO合成で最も重要な重要な酵素であり、多くの生理学的および病理学的機能に直接影響します。たとえば、T細胞と抗原提示細胞の結合体による抗原認識の瞬間、または生体内の特定のホルモンの存在下で、炎症のイベントでサイトカインの放出を促進する[22–25]心血管を弛緩させ、血流を増加ささせます。 私たちのアッセイでは、VEPSは50、100、および200μg/ mLの濃度でRAW264.7の細胞増殖を増加させる能力、また、食細胞NO分泌、iNOS遺伝子およびタンパク質の発現、分泌の著しい増加、私たちのアッセイでは、VEPSは50、100、および200μg/ mLの濃度でRAW264.7の細胞増殖、また、食細胞の著しい増加、NO分泌、iNOS遺伝子およびタンパク質の発現、分泌、IL-6およびIL-1βの遺伝子発現を増加させる能力を示しました。そして平坦化形態を示しました。陽性対照では、LPSはVEPSグループと比較してはるかに強力であることを常に示しています。 LPSの存在下でVEPSを使用した場合にも相乗効果が観察されました(データは示していません)。NOは炎症反応のメディエーターおよびレギュレーターとして認識されています。亜酸化窒素シンターゼには3つのアイソフォームがあるが、iNOSはNO合成で最も重要な重要な酵素であり、多くの生理学的および病理学的機能に直接影響します。たとえば、T細胞と抗原提示細胞の結合体による抗原認識の瞬間、または生体内の特定のホルモンの存在下で、炎症のイベントでサイトカインの放出を促進する[22–25]心血管を弛緩させ、血流を増加ささせます。 マクロファージの細胞形態は、LPSによって活性化されると、丸い形状から平らな形状または広がった形状に変更できることが報告されています[28]。 ただし、RAW264.7の同様の形態がVEPSによって平坦化されたのは今のことです。 平坦化と広がりの形状の程度は、増殖、食作用、NO放出およびiNOS発現と一致しています。形態学的変化のプロセスは、細胞骨格アクチンの複雑で動的な再編成に関与しています[29]。しかし、VEPSによるこれらの活性化は、さらに研究する必要があるにもかかわらず、マクロファージの細胞増殖を有利に増加させ、免疫を促進する可逆的な生理学的プロセスであると考えています。 |
|
この文献は、Inflammation, Vol. 36, No. 4, August 2013 に掲載されたNitric Oxide and Interleukins are Involved in Cell Proliferation of RAW264.7 Macrophages Activated by Viili Exopolysaccharidesを日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |