ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(発酵乳、腸内細菌の科学:研究の最前線) |
免疫生物学的乳酸桿菌はSARS-CoV-2感染に対する呼吸器上皮細胞の耐性を改善します |
Md. Aminul Islam et al., |
以前、経鼻投与された免疫調節性乳酸菌が、トール様受容体3(TLR3)の活性化によって誘導される肺抗ウイルス自然免疫応答を有益に調節し、マウスの呼吸器病原体、インフルエンザウイルス、呼吸器合胞体ウイルスに対する防御を改善したことを報告しました。 ここでは、人間の呼吸器上皮細胞(Calu-3細胞)における生存および生存不能のラクトバチルスプランタルム株の免疫調節効果、そして、これらの免疫生物学的乳酸桿菌が急性呼吸器症候群コロナウイルス2(SARS-CoV-2)感染に対する感受性を低下させる能力を評価しました。 |
|
遺伝的に多様なコロナウイルス科は、鳥類や哺乳類を含む多くの種で循環しています。 過去20年間に、3つの危険なヒトコロナウイルス(CoV)が出現しました。 2002年の重症急性呼吸器症候群CoV(SARS-CoV)、2012年の中東呼吸器症候群CoV(MERS-CoV)、2019年のSARS-CoV-2は、2020年と2021年の間に世界の健康に影響を及ぼしましたコロナウイルス病2019(COVID-19)の原因物質です[1]。 SARS-CoV-2感染者のかなりの割合が無症候性ですが、COVID-19の臨床症状は、咳、発熱、筋肉痛、疲労などの比較的軽度の一般的な風邪のような病気から、重度の呼吸困難、胸部痛、多発性肺炎、性呼吸窮迫症候群(ARDS)、および多臓器機能障害症候群までさまざまです[2]。 世界中の免疫未経験集団におけるSARS-CoV-2の効率的な感染により、このウイルスは世界中で2億1,000万人以上に感染し、2019年12月から2021年8月の間に約450万人が死亡しました。 |
| 生化学的、遺伝的およびゲノム研究により、SARS-CoV-2のスパイクタンパク質がコロナウイルスの宿主細胞への侵入を媒介することが実証されています。 SARS-CoV-2のSタンパク質の受容体結合ドメイン(RBD)は、ヒトアンジオテンシン変換酵素2(ACE2)との相互作用を通じて、ウイルスの認識と宿主細胞への付着を仲介します。 さらに、SARS-CoV-2エントリーには、セリンプロテアーゼTMPRSS2によるSタンパク質の効率的なプライミングが必要です[3]。 したがって、ACE2およびTMPRSS2の発現が高いヒト組織の細胞集団は、SARS-CoV-2感染に対してより脆弱です。 この点で、SARS-CoV-2は、ACE2 + TMPRSS2 +呼吸上皮細胞に感染する能力が高いため、ヒトの上咽頭、気管支[4]および肺[5]で効率的に複製することが示されました。 |
| 呼吸上皮におけるSARS-CoV-2の複製は見過ごされませんが、むしろ免疫系によって媒介される一連の反応を引き起こします。ウイルスの分子パターンがToll様受容体(TLR)やレチノイン酸誘導性遺伝子(RIG)-I様受容体などの宿主細胞パターン認識受容体(PRR)によって認識されると、宿主の生来の抗ウイルス免疫応答が開始されます。 この応答は、シグナル伝達経路の活性化を開始し、最終的にI型インターフェロン(IFN)、ならびに炎症性サイトカインおよびケモカインの産生をもたらします。 これらは一緒になって、病原体を排除する効果的な抗ウイルス反応に不可欠です[6–8]。 I型IFNは、オートクリンまたはパラクリン方式で受容体(IFNAR)を介してシグナル伝達カスケードをトリガーし、シグナル伝達兼転写活性化因子1(STAT1)およびSTAT2のリン酸化を誘導します。 |
| IFNシグナル伝達は、何百もの抗ウイルスタンパク質とケモカインの発現を引き起こし、宿主細胞に抗ウイルス状態を誘導します。 実際、I型IFNと抗ウイルス因子の反応障害は、より重症のCOVID-19症例と関連しています[9–11]。 一方、ウイルス感染は、インターロイキン(IL)-6、ケモカイン(CCモチーフ)リガンド2(CCL2)、CCL5、CCL8、ケモカイン(CXCモチーフ)リガンド8( CXCL8)、CXCL9、CXCL16、およびCXCL2を含む呼吸管での炎症性サイトカインおよびケモカインの産生を引き起こします。 これらの炎症性因子は、免疫細胞を動員して活性化することによりウイルス複製を抑制する上で極めて重要な役割を果たしますが、炎症性因子の不十分な産生がウイルスの病因に寄与する可能性があります[12]。 したがって、COVID-19の重症度は、SARS-CoV-2によって誘発される細胞傷害だけでなく、宿主の免疫応答にも依存します[13]。 |
| 近年、私たちや他の人々は、免疫調節性乳酸桿菌(免疫生物)の経鼻投与によって呼吸器免疫を調節できることを実証しました[14–19]。 この点に関して、マウスモデルでのいくつかの研究は、免疫生物による鼻粘膜のプライミングが、呼吸器合胞体ウイルス(RSV)[17–19]、インフルエンザウイルス(IFV)[20] およびマウスの肺炎ウイルス(PVM)[14–16]によって生成されるものを含むウイルス感染に対する耐性を強化することを示しました。 |
| 動物モデルでの結果と一致して、前向き観察研究は、RSV急性呼吸器感染症の乳児と比較した場合、健康な乳児におけるラクトバチルスの鼻咽頭存在量が高いことを報告しました[21]。 さらに、RSV感染に関連する小児喘鳴疾患を発症するリスクが低下した健康な乳児の気道で、ラクトバチルス属菌の相対的な過剰が検出されました[22]。 これらの有益な免疫調節特性を有する乳酸桿菌株の中には、Lactiplantibacillus plantarum(Basonym Lactobacillus plantarum)種に属する菌株のグループがあり、抗ウイルス免疫を強化する大きな可能性を秘めています(Villena et al。[8]で概説)。 私たちの研究では、TLR3アゴニストであるポリ(I:C)またはRSVによるチャレンジの前に、L。plantarumCRL1506をマウスに経鼻投与すると、呼吸器の自然免疫応答が特異的に調節され、ウイルスクリアランスが改善され、炎症性肺損傷が減少しました[17 ]。 興味深いことに、我々はまた、熱殺菌したL. plantarum CRL1506による鼻のプライミングが呼吸器の抗ウイルス免疫応答を調節できることを示し、生存率が免疫生物学的乳酸桿菌がそれらの保護効果を発揮するための必須因子ではないことを示しています[17]。 |
| これらの以前の研究は、一般に免疫調節性乳酸桿菌、特にL. plantarumが、SARS-CoV-2に対する呼吸器の自然免疫応答を有益に調節できるだろうという推測を促しました。 したがって、この研究の目的は、生存可能または生存不可能な免疫調節性L. plantarum株がサイトカイン産生を特異的に調節できるかどうか、TLR3アゴニストポリ(I:C)で刺激された、またはSARS-CoV-2でチャレンジされた呼吸器上皮細胞における抗ウイルス因子の発現は、このようにしてウイルス複製を減少させるかどうかを評価することでした。 |
| 2.1 免疫調節性乳酸桿菌 | Lactiplantibacillus plantarum CRL1506およびL.plantarum CRL1905は、CERELAカルチャーコレクション(CERELA-CONICET、アルゼンチン、トゥクマン)に属しています。 L. plantarum MPL16およびL.plantarum MPL18は、Food and Feed Immunology Group Culture Collection(東北大学、仙台、日本)に属しています。 乳酸桿菌株の増殖には、Man–Rogosa–Sharpe(MRS)ブロスを37℃で使用しました。 インビトロ免疫調節アッセイのために、一晩の培養物を遠心分離によって収集し、滅菌リン酸緩衝生理食塩水(PBS)で3回洗浄した。 乳酸桿菌はペトロフハウザー計数チャンバーで計数され、使用するまでダルベッコの改良イーグル培地(DMEM)に再懸濁されました。 MPL16株とCRL1506株は免疫調節機能のために選択され、CRL681株とMPL18株はネガティブコントロールとして選択されました[23,24]。 生存不能なL.plantarum CRL1506およびMPL16は、80℃のウォーターバスで30分間のtyndallizationによって得られ、細菌の増殖の欠如はMRS寒天プレートを使用して確認されました[17–19]。 |
| 2.2 細胞培養 |
| ヒト肺上皮細胞株Calu-3(HTB-55、ATCC、マナッサス、バージニア州、米国)は、ヒト肺腺癌に由来します。 Calu-3細胞は、20%ウシ胎児血清(FBS)と1%非必須アミノ酸溶液(Gibco、Grand Island、NY、USA)を添加したDMEMで増殖させました。 FBSを10%に減らし、1%ペニシリン/ストレプトマイシン(Gibco)を抗ウイルスアッセイで培地に添加しました。 アフリカングリーンモンキー腎臓上皮細胞株Vero76(ATCC CRL-1587)は、10%FBSを添加した最小必須培地(MEM)で培養しました。 FBSは抗ウイルスアッセイで2%に減少しました。 細胞は、以前に記載されたように[25]、5%CO2を含む雰囲気中の加湿インキュベーター内で37℃でインキュベートされた。 |
| 2.3 培養上清中のサイトカイン濃度 |
| TLR3の活性化によって誘導される自然免疫応答における乳酸桿菌の影響を評価するために、異なるL. plantarum株(5×107細胞/ mL)を含む1mLのDMEMを2×106細胞/ウェルの密度で6ウェルプレートで培養したCalu-3細胞単層に添加しました。 細胞をさらに37℃、5%CO2で24時間インキュベートした。 PBSで洗浄することにより乳酸桿菌を除去し、合成二本鎖RNA(dsRNA)とTLR3アゴニストを使用し、Calu-3細胞を15 g / mLのTLR3アゴニストポリ(I:C)(Sigma-Aldrich、サンルイス、ミズーリ州、米国)で刺激しました[25]。 TLR3アゴニストポリ(I:C)(基礎レベル)でチャレンジする前とTLR3活性化の48時間後に、前述のように酵素結合免疫吸着測定法(ELISA)技術キットによるサイトカインの評価のために培養上清を収集しました[25]。 IFN- β(感度:0.7 pg / mL)、IL-6(感度:0.7 pg / mL)、CCL5(またはRANTES)(感度:6.6 pg / mL)、CXCL8(またはIL-8)(感度: 7.5 pg / mL)およびCXCL10(またはIP-10)(感度:4.46 pg / mL)は、製造元(R&D Systems、ミネソタ州ミネアポリス、米国)の推奨に従って市販のELISAキットで測定しました。 |
| 2.4 SARS-CoV-2感染 |
| SARS-CoV-2(臨床分離株hCoV-19 / USA / VA / 2020)のワーキングストックがVero76細胞で得られました。 ACE2発現の高いCalu-3細胞の亜集団であるCalu-32B4は、SARS-CoV-2感染実験で使用されました。 これらの細胞は、以前に記載されているように[25–27] ACE2抗体で選別することによって得られました。 Calu-3 2B4細胞は、異なるL. plantarum株(5107細胞/ mL)を含む1 mLのDMEMで、37℃、5%CO2で24時間刺激されました。 PBS洗浄を使用して乳酸桿菌を除去し、次にCalu-32B4細胞を37℃でSARS-CoV-2(MOI0.1)に感染させた。 30分後、結合していないウイルスをPBS洗浄により除去し、細胞を37℃、5%CO2で48時間または72時間インキュベートしました。 Calu-3 2B4細胞におけるSARS-CoV-2の複製は、感染後48時間および72時間で評価されました。これは、これらの細胞におけるこのウイルスの複製速度がVero76細胞で観察されたものよりも遅いという以前の報告を考慮したものです [28]。 |
| 感染性SARS-CoV-2のプラーク形成単位(PFU)は、他の場所で説明されているように[29–31]、わずかな変更を加えて、Vero76細胞でのプラーク滴定によって定量化されました。 Vero 76を24ウェルプレートに播種し、PBSで洗浄し、SARSCoV-2を含む細胞培養上清の段階希釈液と2重にインキュベートし、DMEM中の1.2%Avicelで覆いました。 72時間後に細胞を6%ホルマリンで固定し、クリスタルバイオレットで染色して可視化しました[25]。 |
| 原形質膜の損傷時に放出される細胞質ゾル酵素である乳酸デヒドロゲナーゼ(LDH)は、細胞毒性の指標としてCalu-32B4細胞の上清で測定されました。 LDHアッセイキット(Weiner Lab、ボストン、マサチューセッツ州、米国)を使用して、Calu-32B4細胞の上清中のLDHレベルを測定しました。 IFN-、IL-6、CCL5、CXCL8およびCXCL10の濃度は、上記のようにELISAキットによって測定された。 すべての実験は、COVID-19に関連するバイオセーフティガイダンス[32,33]に従って、バイオセーフティレベル3(BSL-3)施設で実施されました。 |
| 2.5 2段階リアルタイム定量PCRによる定量的発現解析 |
| Calu-3細胞における免疫因子の発現を特徴づけるために、2段階のリアルタイム定量PCR(qPCR)が実施されました。 各Calu-3細胞サンプルからのトータルRNA分離は、TRIzol試薬(Invitrogen、米国マサチューセッツ州ウォルサム)を使用して実施しました。 Quantitect RTキット(Qiagen、東京、日本)を、製造元の推奨に従ってcDNAの合成に使用しました。 リアルタイムqPCRは、7300リアルタイムPCRシステム(Applied Biosystems、ウォリントン、英国)およびプラチナSYBRグリーンqPCRスーパーミックスウラシル-DNAグリコシラーゼと6-カルボキシル-X-ローダミン(ROX)(Invitrogen)を使用して実施しました。 TMPRSS2のプライマー(F:CAAGTGCTCCAACTCTGGGAT、 R:AACACACCGATTCTCGTCCTC)、ACE2(F:ACAGTCCACACTTGCCCAAAT、 R:GAGAGCACTGAAGACCCATT)、DDX58(F:TGCGAATCAGATCCCAGTGTA、 R:TGCCTGTAACTCTATACCCATGT)、 およびIFNAR2 (F:TCATGGTGTATATCAGCCTCGT、 R:AGTTGGTACAATGGAGTGGT TTT)は、Sun らに従って使用されました。 [34]。 2'5'-OAS1のプライマー(F:AGGAAAGGTG CTTCCGAGGTAG、 R:GGACTGAGGAAGACAACCAGGT)およびMx1(F:TTCAGCACCTGATGGCCTATC、 R:TGGATGATCAAAGGGATGTGG)は、Shuai らに従って使用されました [35]。 |
| PCRサイクリング条件は、50℃で2分、続いて95℃で2分、そして95℃で15秒、60℃で30秒、72℃で30秒の40サイクルでした。 反応混合物には、5mLのサンプルcDNAと15mLのマスターミックスが含まれ、これにはセンスプライマーとアンチセンスプライマーが含まれていました。 ハウスキーピング遺伝子ATF4の発現(F:CTCCGGGACAGATTGGATGTT、 R:Calu-3細胞におけるSARS-CoV-2感染中に差次的発現がないGGCTGCTTATTAGTCTCCTGGAC)[34]を参照遺伝子として使用した。 |
| 2.6 統計分析 |
| 結果は、3回行った実験からの平均標準偏差(SD)として表されました。 正規分布データは、2元配置分散分析によってテストされました。 テューキーの検定(平均のペアワイズ比較用)またはフィッシャーの最小有意差(LSD)検定(多重比較用)を使用して、グループ間の差を評価しました。 差は、p <0.05で有意であると見なされました。 |
| 3. 結果 |
| 3.1 免疫生物学的L. plantarum株は、TLR3アゴニストポリ(I:C)刺激に対する呼吸器上皮細胞の応答を特異的に調節します |
| 私たちの最初の目的は、TLR3アゴニストポリ(I:C)チャレンジに対する呼吸器上皮細胞の応答に対する免疫調節性L. plantarum株の影響を研究することでした。 Calu-3細胞をL.plantarum MPL16またはCRL1506で処理しても、有害な影響を引き起こしたり、培養上清のLDHレベルを変更したりすることはありませんでした(データは示していません)。 ポリ(I:C)(基礎レベル)で刺激する前に、L.plantarum MPL16とCRL1506の両方でCalu-3細胞を処理すると、培養上清中のIFN-βとIL-6の濃度が大幅に増加しましたが、 CXCL8が観察されました(図1)。 MPL16株は、L.plantarum CRL1506と同じくらい効率的にIFN-βおよびIL-6の基礎生産を増加させました。 以前に記載されているように[25]、ELISAキットを使用しても、培養されたCalu-3細胞の上清ではCXCL10またはCCL5の基礎レベル(図1)は検出されませんでした。 |
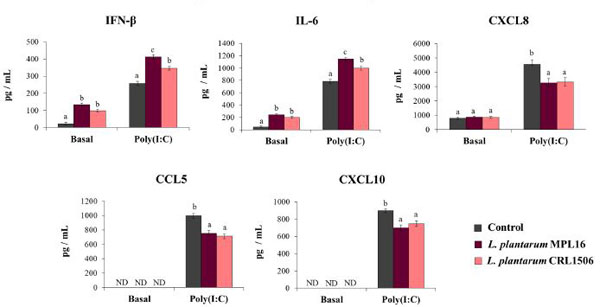 |
| 図1 トール様受容体3(TLR3)シグナル伝達活性化によって誘導される呼吸器上皮細胞の抗ウイルス免疫応答に対するL. plantarum株の影響 |
Calu-3細胞をL.plantarumMPL16またはCRL1506で処理しました (5107細胞/ mL)24時間、次にポリ(I:C)で48時間刺激。 乳酸桿菌処理を行わない呼吸器上皮細胞を対照として使用した。 TLR3活性化の前(基本グループ)と後(ポリ(I:C)グループ)に、ELISAを使用したサイトカインの測定のために培養上清を収集しました。 ND:検出されません。 結果は、3つの独立した実験からのデータを表しています。 文字は有意差(p <0.05)、a <b <cを示します。 |
| ポリ(I:C)による刺激に対するCalu-3細胞の応答に対する免疫生物学的乳酸桿菌の影響も評価されました。 最近説明したように、ポリ(I:C)でCalu-3細胞に挑戦すると、IFN-βレベルと、コントロール細胞のIL-6およびCXCL8濃度が向上しました[25]。 さらに、検出可能な濃度のCXCL10およびCCL5が、ポリ(I:C)チャレンジ細胞の上清に見られました(図1)。 L. plantarum MPL16またはCRL1506による呼吸器上皮細胞の前処理は、対照細胞と比較した場合、IFN-βおよびIL-6レベルを有意に増強し、3つのケモカインの濃度を低下させました。 注目すべきことに、MPL16株はIFN-βとIL-6の増強においてL. plantarum CRL1506よりも効率的でしたが、2つの株はCXCL8、CCL5、CXCL10の減少に同等に効果的でした(図1)。 |
| 3.2 免疫生物学的L.plantarum株はSARS-CoV-2感染に対する呼吸上皮細胞の耐性を増加させる |
| コロナウイルスは、複製およびmRNA転写中にdsRNA分子を合成することができ[36]、SARS-CoV-2は気道の上皮細胞でdsRNAを感知する抗ウイルスシステムによって検出されることがわかりました[28]。 さらに、最近、Calu-3 2B4細胞にSARS-CoV-2を感染させると、ポリ(I:C)による刺激に似たIFN-βおよび炎症性サイトカインとケモカインのプロファイルが誘導されることを報告しました[25]。 次に、免疫生物学的乳酸桿菌が、invivo [17,18,19]およびCalu-3細胞(図1)でポリ(I:C)刺激によって引き起こされるI型IFNおよびサイトカインプロファイルを差別的に調節する能力を考慮して、L.plantarum MPL16およびCRL1506は、Calu-32B4細胞におけるSARS-CoV-2複製に影響を与える可能性があります。 したがって、我々は、Calu-32B4細胞を生存可能なL.plantarum MPL16またはCRL1506で刺激し、続いて呼吸器上皮細胞にSARS-CoV-2でチャレンジしました。 ウイルス力価とLDHは、感染後48時間と72時間で評価されました(図2)。 |
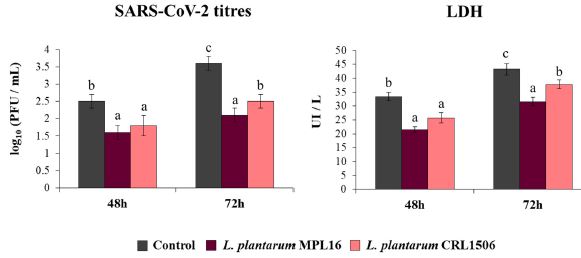 |
| 図2 重症急性呼吸器症候群ウイルス2(SARS-CoV-2)の感染に対する呼吸器上皮細胞の耐性に対するL.plantarum株の影響 |
Calu-3 2B4細胞をL. plantarum MPL16またはCRL1506(5×107細胞/ mL)で24時間処理した後、コロナウイルスに感染させました。 乳酸桿菌処理を行わない呼吸器上皮細胞を対照として使用した。 培養上清中のSARS-CoV-2力価とLDHレベルは、感染後48時間と72時間で評価されました。 |
| SARS-CoV-2の複製は、評価された2つの時点で観察され、72時間で力価が高くなりました。 48時間と比較した場合、72時間目に感染細胞の上清にも高いLDHレベルが見られました。 Calu-3 2B4細胞を免疫生物学的乳酸桿菌で前処理すると、対照群と比較して、コロナウイルスの複製と48時間および72時間でのLDHの値が大幅に減少しました。 両方の乳酸桿菌は感染後48時間でコロナウイルス力価とLDHレベルを低下させるのに同等に効果的でしたが、L. plantarum MPL16はCRL1506株よりも72時間でこれらのパラメーターを低下させる効果がありました(図2) |
| SARS-CoV-2の感染に対する免疫生物処理されたCalu-32B4細胞の耐性の改善が、抗ウイルス免疫応答の増強に関連していることを示すために、チャレンジした細胞の上清培養物中のIFN-β、IL-6、およびケモカインの濃度も測定しました(図3)。 SARS-CoV-2チャレンジは、評価された2つの時点ですべてのサイトカインの産生を誘発しました。 注目すべきことに、CXCL8、CCL5、CXCL10のレベルは、72時間よりも48時間の方が大きかった。 Calu-3 2B4細胞を免疫生物学的乳酸桿菌で前処理すると、SARS-CoV-2感染によって誘導されるIFN-βおよびIL-6レベルが大幅に増強されました。 感染後48時間でMPL16およびCRL1506で処理した細胞間でIFN-βおよびIL-6のレベルに差は観察されませんでしたが、L.plantarum MPL16はCRL1506株よりも効率的で、2つの免疫因子を72感染後で増加させました(図3)。 両方の乳酸桿菌は、評価された2時間でCXCL8濃度を低下させるのに同等に効果的でしたが、感染後48時間でCCL5とCXCL10のレベルを分析した場合、乳酸桿菌処理の効果は検出されませんでした(図3)。 ただし、MPL16またはCRL1506株で処理されたCalu-3 2B4細胞は、SARS-CoV-2感染後72時間で有意に低いCCL5およびCXCL10レベルを生成しました。 注目すべきことに、L. plantarum MPL16は、この効果の誘導においてCRL1506株よりも効率的でした(図3)。 |
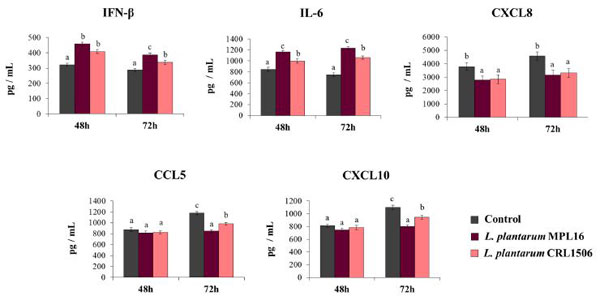 |
| 図3 重症急性呼吸器症候群ウイルス2(SARS-CoV-2)の感染によって誘発される呼吸器上皮細胞の抗ウイルス免疫応答に対するL. plantarum株の影響 |
| Calu-3 2B4細胞をL. plantarum MPL16またはCRL1506(5×107細胞/ mL)で24時間処理した後、コロナウイルスに感染させました。 乳酸桿菌処理を行わない呼吸器上皮細胞を対照として使用した。 感染後48時間および72時間にELISAを使用してサイトカインを測定するために、培養上清を収集しました。 結果は、3つの独立した実験からのデータを表しています。 文字は有意差(p <0.05)、a <b <cを示します。 |
| また、SARS-CoV-2が感染後6時間目と12時間目に呼吸上皮細胞に侵入するために使用する受容体の発現を評価しました。これは、コロナウイルスがCalu-3細胞の攻撃後早期に発現を調節できることが示唆されているためです。 [34]。 ACE2の発現レベルは評価された2つの時点で類似していたが、TMPRSS2の発現は12時間よりも6時間で有意に高かった(図4)。 乳酸桿菌処理細胞におけるACE2およびTMPRSS2の発現を対照と比較した場合、違いは見られませんでした。 PRR TLR3とDDX58、および受容体IFNAR2の発現の増加は、SARS-CoV-2に感染したCalu-3細胞で見られ、アップレギュレーションは6時間よりも12時間で顕著でした(図4)。 L. plantarum MPL16とCRL1506はどちらも、研究した2つの時点で、対照細胞と比較した場合、TLR3、DDX58、およびIFNAR2の発現を有意に増強することができました。 ただし、MPL16株は、感染後12時間でDDX58とIFNAR2を増加させる点で、L.plantarumCRL1506よりも効率的でした(図4)。 さらに、Calu-3細胞は、感染後48時間および72時間でMx1およびOAS1の発現を増加させました。 MPL16およびCRL1506処理は、評価された2つの時点で、対照細胞と比較した場合、両方の抗ウイルス因子の発現を有意に増強することができました。 しかし、L.plantarumMPL16は、SARS-CoV-2感染後72時間でMx1とOAS1の増加において、CRL1506株よりも効率的でした(図4)。 |
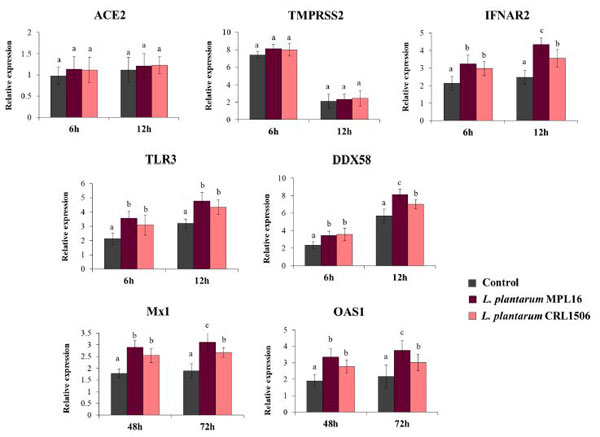 |
| 図4 重症急性呼吸器症候群ウイルス2(SARS-CoV-2)の感染によって誘発される呼吸器上皮細胞の抗ウイルス免疫応答に対するL. plantarum株の影響 |
| Calu-3細胞をL.plantarum MPL16またはCRL1506(5107細胞/ mL)で24時間処理した後、コロナウイルスに感染させました。 呼吸器上皮細胞の乳酸桿菌処理を対照として使用した。 ACE2、TMPRSS2および抗ウイルス因子は、感染後の示された時点でリアルタイムqPCRによって決定されました。 ND:検出されません。 結果は、3つの独立した実験からのデータを表しています。 文字は有意差(p <0.05)、a <b <cを示します。 |
| 3.3 生きているL.プランタルム株は呼吸上皮細胞における抗ウイルス免疫応答の調節に必要である |
| 我々は以前に、マウスに鼻経路で投与された生存不能な免疫生物学的乳酸桿菌が、ポリ(I:C)投与またはRSV感染によって引き起こされる呼吸器の自然免疫応答を特異的に調節できることを報告しました[17、18、19]。 次に、私たちの次の目的は、生存不能なL. plantarum MPL16またはCRL1506がTLR3の活性化に対するCalu-3細胞の応答を調節できるかどうかを評価することでした。 この目的のために、呼吸上皮細胞を熱殺菌(HK)乳酸桿菌で処理し、次にポリ(I:C)でチャレンジしました。 サイトカインは、TLR3活性化の前後にCalu-3細胞の上清で評価されました。 図5に示すように、HK乳酸桿菌による呼吸器上皮細胞の処理は、IFN-β、IL-6、またはケモカインCXCL8、CCL5、CXCL10の基礎レベルに変化を引き起こしませんでした。 さらに、ポリ(I:C)で刺激した後、対照呼吸上皮細胞をHK乳酸菌処理細胞と比較した場合、IFN-β、IL-6、またはケモカインの濃度に差は見られませんでした(図5)。 |
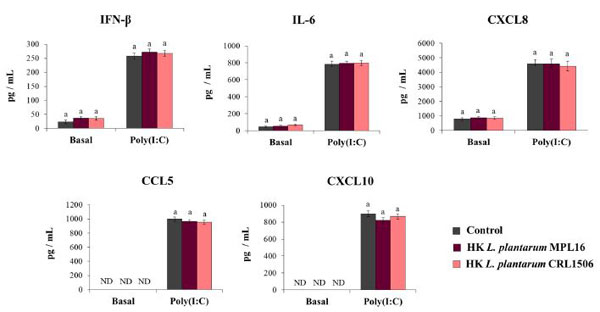 |
| 図5.トール様受容体3(TLR3)の活性化によって誘導される呼吸器上皮細胞の抗ウイルス免疫応答に対する生存不能なL.plantarum株の影響 |
Calu-3細胞を熱殺菌(HK)L.plantarum MPL16またはCRL1506(5×107細胞/ mL)で24時間処理した後、ポリ(I:C)で48時間刺激しました。 ND:検出されません。 結果は、3つの独立した実験からのデータを表しています。 同様の文字は、有意差がないことを示します(p <0.05)。 |
| また、ポリ(I:C)刺激前の免疫調節活性がないため、HK乳酸菌がコロナウイルス感染に対する呼吸器上皮細胞の応答に影響を与えることができないことを実証することも目的としました。 この仮説を検証するために、Calu-32B4細胞をHKL。plantarum MPL16またはCRL1506で前処理し、コロナウイルスに感染させました。 予想通り、HKMPL16もHKCRL1506も、SARS-CoV-2力価またはLDHレベルを低下させることができませんでした(図6)。 |
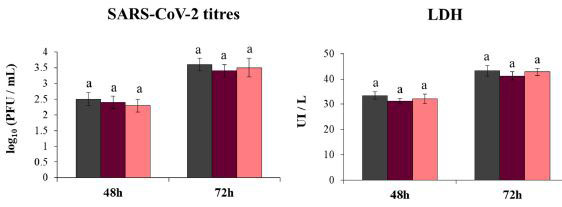 |
| 図6. 重症急性呼吸器症候群ウイルス2(SARS-CoV-2)の感染に対する呼吸器上皮細胞の耐性に対する生存不能なL.plantarum株の影響 |
Calu-3 2B4細胞を熱殺菌(HK)L.plantarum(5107細胞/ mL)で24時間処理した後、コロナウイルスに感染させました。 結果は、3つの独立した実験からのデータを表しています。 同様の文字は、有意差がないことを示します(p <0.05)。 |
| 3.4 L.plantarumが呼吸器上皮細胞の抗ウイルス免疫を調節する能力は菌株依存性である |
| 粘膜免疫応答に対するプロバイオティクス乳酸菌の効果は、菌株特異的特性であることが示されています。 実際、ある乳酸桿菌菌株の免疫調節能力は、同じ種であっても他の菌株に外挿することはできません[24,37]。 したがって、TLR3の活性化との関連で呼吸器上皮細胞に対する非免疫調節性L.plantarum株の影響を評価することを目的とした。 この目的のために、非免疫調節株CRL1905およびMPL18 [23,24,38]で呼吸器上皮細胞を刺激し、poly(I:C)でチャレンジしました。 図7に示すように、L.plantarumCRL1905およびMPL18は、poly(I:C)でチャレンジする前に、Calu-3細胞の上清中のIFN-β、IL-6、CXCL8、CCL5、またはCXCL10のレベルを変更できませんでした。 )。 CRL1905およびMPL18株は、TLR3の活性化後、IFN-β、IL-6、およびケモカインの濃度の変化を誘発することもできませんでした(図7)。 |
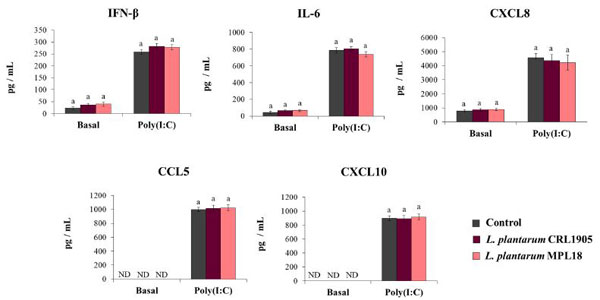 |
| 図7.Toll様受容体3(TLR3)の活性化によって誘導される呼吸器上皮細胞の抗ウイルス免疫応答に対する非免疫調節性L. plantarum株の影響 |
Calu-3細胞をL.plantarum CRL1905またはMPL16(5×107細胞/ mL)で24時間処理した後、ポリ(I:C)で48時間刺激しました。 乳酸桿菌処理を行わない呼吸器上皮細胞を対照として使用した。 TLR3活性化の前(基本グループ)と後(ポリ(I:C)グループ)に、ELISAを使用したサイトカインの測定またはリアルタイムqPCRを使用した抗ウイルス因子のために培養上清を収集しました。 ND:検出されません。 結果は、3つの独立した実験からのデータを表しています。 同様の文字は、有意差がないことを示します。 |
| 最後に、L.plantarum CRL1905およびMPL18が、コロナウイルスチャレンジに対するCalu-3 2B4細胞の耐性を改変する能力を評価しました(図8)。 CRL1905株もMPL18株も、評価された2つの時点で、感染した呼吸器上皮細胞のコロナウイルス力価を低下させることができませんでした。 これらの結果と一致して、CRL1905およびMPL18で処理されたCalu-3 2B4細胞のIFN-βのレベルは、SARS-CoV-2感染後48時間および72時間の両方で対照細胞と比較して異ならなかった(図8)。 |
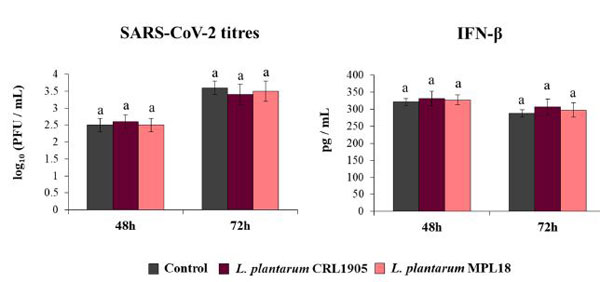 |
| 図8. 重症急性呼吸器症候群ウイルス2(SARS-CoV-2)の感染に対する呼吸器上皮細胞の耐性に対する非免疫調節性L.plantarum株の影響 |
| Calu-3 2B4細胞をL.plantarum CRL1905またはMPL18(5×107細胞/ mL)で24時間処理した後、コロナウイルスに感染させました。 乳酸桿菌処理を行わない呼吸器上皮細胞を対照として使用した。 培養上清中のSARS-CoV-2力価およびインターフェロン(IFN)-βレベルは、感染後48時間および72時間で測定されました。 結果は、3つの独立した実験からのデータを表しています。 同様の文字は、有意差がないことを示します(p <0.05)。 |
| 4. 議論 |
| IFVやRSVのようないくつかの呼吸器ウイルスについて説明したように、免疫応答はSARS-CoV-2誘発性肺炎の結果に重要な役割を果たします。 急性感染症は、免疫性炎症性肺損傷により悪化した疾患を誘発する可能性があり、重症のCOVID-19患者に重篤な罹患率と死亡率をもたらします[12、13]。 したがって、ウイルス誘発性免疫病理学を調節するための新しいアプローチを特定することは、SARS-CoV-2によって引き起こされるような急性および致死性の呼吸器ウイルス感染症を予防または治療するのに有益であろう。 この点に関して、私たちの以前の研究は、経鼻投与された免疫生物学的乳酸桿菌が、ウイルス感染の状況において呼吸器免疫応答に有益に影響を与える可能性があることを明らかにしました[17、18、19]。 私たちの手によって、経鼻投与されたL.plantarum CRL1506は、ポリ(I:C)による鼻腔チャレンジまたはRSV感染後のマウスの気管支肺胞洗浄液中のIFN-α、IFN-β、IL-6、およびIFN-γのレベルを増加させることができました。 これらの影響は、ウイルス複製と肺損傷の有意な減少と関連していた[17]。 さらに、CRL1506株が気道のMHC-II + CD11c + CD11blowCD103 +およびMHC-II + CD11c + CD11bhighCD103-樹状細胞とCD3 + CD4 +IFN-γ+ T細胞を増加させることができることを示しました。 免疫細胞の数と活動を調節するその能力を示しています。 この研究で示された実験は、L.plantarum CRL1506の免疫調節活性における呼吸器上皮細胞の重要な寄与に取り組んだ以前の研究の直接の拡張を表しています[17]。 ここで、Calu-3細胞を免疫生物学的L. plantarum株で前処理すると、IFN-βとIL-6の産生が大幅に増加し、CCL5、CXCL8、CXCL-10が減少することが観察されました。 したがって、L.plantarum CRL1506は、経鼻投与されたときに呼吸器免疫細胞と上皮細胞の両方と相互作用することができると推測するのは魅力的です。 したがって、抗ウイルス防御を改善する変化を誘発します。 |
| また、Calu-3細胞を免疫生物学的L. plantarum株で前処理すると、SARS-CoV-2感染に対する耐性が高まることも示しました。 私たちの知る限り、SARS-CoV-2感染に対する免疫調節性プロバイオティクス乳酸菌の効果を報告した研究はありません。 ある報告では、SARS-CoV-2でチャレンジする前に乳酸菌で2時間刺激したVero E6細胞を使用し、感染後3日後にMTTテストを使用して感染細胞の生存率を評価しました[39]。 この研究では、VeroE6細胞をLimosilactobacillus fermentum 90TC4で処理すると、感染したコントロールと比較して、SARS-CoV-2でチャレンジした細胞の生存率が大幅に向上したことが説明されています。 しかし、その研究は、乳酸桿菌の有益な効果を説明するための可能なメカニズムを提供しませんでした。 したがって、呼吸器上皮細胞を使用し、自然免疫応答を評価することにより、SARS-CoV-2感染の状況で乳酸桿菌の潜在的な保護効果のより正確な評価を行うことに興味がありました。 ここでは、免疫生物学的L. plantarum株が、SARS-CoV-2感染によって引き起こされる呼吸器上皮細胞の自然免疫応答に影響を及ぼし、ウイルス複製に対する細胞の耐性を改善できることを示しました。 |
| Calu-3細胞でのハイスループットシーケンシングトランスクリプトミクス研究により、SARS-CoV-2の急速な増殖が明らかになり、これには宿主遺伝子の初期の集中的な応答が伴いました[34]。 SARS-CoV-2は、感染の最初の24時間はACE2の発現に変化を引き起こさなかったが、Calu-3細胞ではTMPRSS2を有意に増加させた[34]。 これらの発見と一致して、感染後6時間目と12時間目にCalu-3細胞でのACE2の発現の変化は観察されませんでした。 一方、TMPRSS2は6時間目に大幅にアップレギュレーションされました。 ACE2およびTMPRSS2は感受性細胞へのSARS-CoV-2侵入に必須であると報告されているため[4]、免疫生物学的L.plantarumがウイルスの複製を減少させる能力に関与する1つの考えられるメカニズムは侵入受容体の発現の減少です。 私たちの研究では、Calu-3細胞をL. plantarum MPL16またはCRL1506で処理しても、対照細胞と比較してSARS-CoV-2によるチャレンジの前(データは示していない)または後(図4)にACE2またはTMPRSS2の発現レベルの変化は誘発されませんでした。 我々の発見を支持して、Calu-3およびCaco-2細胞で行われた比較研究およびSARS-CoV-2の侵入受容体のプロファイルの評価は、Calu-3細胞におけるACE2およびTMPRSS2の発現が1〜4倍であり、Caco-2細胞よりもそれぞれ4〜64倍高いことを発見した [35]。 興味深いことに、この研究では、SARS-CoV-2の感染力が両方の細胞株で同様に効率的であることがわかりました。 これらの結果は、このメカニズムによるウイルス複製の阻害を達成するために、受容体の発現におけるかなり大きな変動が必要であることを示している。 したがって、侵入受容体発現の調節は、免疫生物性乳酸桿菌によって誘発されるSARS-CoV-2に対する保護効果には関与しないであろう。 |
| 感染すると、SARS-CoV-2は、サイトカインと抗ウイルス因子の有意なアップレギュレーションを特徴とする呼吸器上皮細胞で数百の遺伝子の発現も誘導します[34]。 Calu-3細胞での転写研究により、コロナウイルスがTLR3およびRNAウイルス認識DDX58の標準的なRIG-I様受容体の発現を増加させることがわかりました。 さらに、SARS-CoV-2に24時間感染させた後、Calu-3細胞でIRF7、IRF9、STAT1、およびSTAT2遺伝子の段階的なアップレギュレーションが観察されました[34]。 |
| これらの発見と一致して、SARS-CoV-2でCalu-3細胞に挑戦することでJAK / STATシグナル伝達経路を刺激できることが実証されました。 これは、哺乳類の自然免疫応答の重要な腕を構成するI型(IFN-α/β)およびIII型(IFN-λ)IFNの産生を促進します[6]。 注目すべきことに、Calu-3細胞を使用すると、SARS-CoV-2ウイルス複製がI型IFN治療によって用量依存的に阻害されることが報告されました[6]。 さらに、呼吸器上皮細胞における抗ウイルスJAK / STATシグナル伝達経路の阻害は、SARS-CoV-2増殖を有意に増強することが実証されました[6]。 SARS-CoVと比較したSARS-CoV-2のI型IFN感受性を評価することを目的とした比較研究は、SARS-CoV-2はCalu-3細胞でSARS-CoVと同様のウイルス複製を維持しますが、新規コロナウイルスはI型IFNに対してはるかに感受性が高くなります[40]。 次に、I型IFNの早期産生(または投与)は、呼吸器上皮細胞におけるSARS-CoV-2複製を効率的に打ち消します[41]。 これらの発見は、MPL16やCRL1506のような免疫生物学的L.plantarum株が、IFN-βの産生の増加とそれに続くウイルス感知受容体TLR3や DDX58の発現の増強を通じてSARS-CoV-2感染に対する耐性を改善できることを示しています。 ウイルス複製を制限するのに役立つ抗ウイルス因子Mx1とOAS1も同様です。 したがって、Calu-3細胞を生存不能な乳酸桿菌で処理しても、以下に説明するように、IFN-β産生を調節したり、ウイルス力価を低下させたりすることはできませんでした。 |
| 我々の結果はまた、免疫生物学的L. plantarum MPL16およびCRL1506によって誘導されるサイトカインプロファイルの変化が、ウイルス複製を制限するのに役立つだけでなく、調節不全の免疫応答によって引き起こされる炎症性損傷の軽減にも協力できることを示唆しています。 遅延したIFN応答は強い炎症反応の誘発と関連しており、免疫病理学と重度のCOVID-19をもたらすことが示されました[42]。 さらに、SARS-CoV-2感染は、呼吸上皮の核因子カッパB(NF-kB)シグナル伝達経路を強力に活性化し、炎症性サイトカイン/ケモカインのアップレギュレーションを引き起こします[34]。 転写研究により、SARS-CoV-2に感染したCalu-3細胞におけるCCL2、CXCL8、CSF3、CSF2、およびCXCL10などのCOVID-19の重度の臨床症状を持つ患者の血清で強く上昇することが示された炎症性因子[43,44]の発現が著しく増強されていることが明らかになりました[34]。 これらの以前の研究と一致して、SARS-CoV-2感染がCalu-3細胞におけるCXCL8、CCL5、およびCXCL10の産生を増加させることをここで観察しました。 注目すべきことに、L.plantarum MPL16およびCRL1506は、好中球およびT細胞に対するこれらの走化性物質を有意に減少させました。 上皮細胞が気道内の免疫細胞の動員と活性化に影響を与える能力を持っていることを考慮すると[45]、IFN-βの改善とCXCL8、CCL5、CXCL10の減少が免疫生物学的L.plantarumは、炎症性損傷を制御するためにinvivoで寄与するであろう。 さらに、L. plantarumCRL1506によるマウスの鼻プライミングは、ポリ(I:C)投与またはRSV感染後に肺CD3 + CD4 + IL-10 + T細胞を増加させ、炎症性肺組織の損傷の制御に寄与することを以前に示しました [17]。 したがって、呼吸上皮および/または制御性T細胞応答の調節を介した、SARS-CoV-2感染の状況におけるMPL16およびCRL1506株の潜在的な抗炎症保護効果の評価は、さらなる調査を必要とします。 |
| 私たちの以前の研究は、腸管[24,46]および呼吸器[17,18,19]管における抗ウイルス免疫応答をinvivoで有益に調節する乳酸桿菌の菌株依存性能力を実証しました。 さらに、腸上皮細胞培養で実施された転写研究により、TLR3を介した抗ウイルス免疫応答およびロタウイルス感染に対する耐性を調節する乳酸桿菌の菌株依存性能力が明らかになりました[23、24、47]。 ここで呼吸上皮細胞培養で得られた結果は、これらの以前の発見と一致しています。 L.plantarum MPL16およびCRL1506は、TLR3の活性化によって引き起こされる呼吸上皮細胞の応答を特異的に調節し、SARS-CoV-2感染に対する耐性を高めましたが、L.plantarum MPL18およびCRL1905株はこれらの利点を誘導できませんでした。 さらに、MPL16株とCRL1506株の両方が、Calu-3細胞の抗ウイルス免疫応答を有益に調節することができましたが、L.plantarum MPL16はこの効果をより効率的に達成しました。 したがって、ここに提示された結果は、SARS-CoV-2によって生成されたものを含む、特定のウイルス感染から保護するために使用できる特定の株の徹底的な研究の必要性を強調しています。 |
| 生存不能なL.plantarum MPL16またはCRL1506は、Calu-3細胞におけるポリ(I:C)誘発免疫応答またはSARS-CoV-2に対する耐性を調節することができなかったという事実に興味を持って注目します。 以前のinvivo研究では、TLR3アゴニストpoly(I:C)で鼻腔内チャレンジされた、またはRSVに感染したマウスの気道で、HK L. plantarum CRL1506がI型IFN、IL-6の産生、およびCD11c + CD11blowCD103 +樹状細胞の数を増加させることができることが示されたため、これは予想外でした[17]。 これらの対照的な結果は、Rosenbergと同僚によっても報告されました。 研究者らは、HK免疫生物学的L. plantarum NCIMB 8826によるマウスの鼻腔プライミングが、PVM [14,15,16]およびIFV [48]感染の致死的後遺症に対する保護を提供することを実証しました。 NCIMB8826株のinvivo保護効果は、ウイルスの複製とクリアランスの減少に関連していませんでしたが、肺の損傷を最小限に抑える炎症反応の効率的な調節に関連していました[15、16、48]。 注目すべきは、MLE-12細胞(マウス肺II型肺胞上皮様細胞)とmTECs細胞(分極したマウス気管上皮細胞)の2つの細胞培養モデルでの研究で、HK L. plantarum NCIMB 8826が、in vivoで有効な実験条件を再現したプロトコルを使用して、IFV感染によって誘発されるCCL2、CCL5、CXCL1、およびCXCL10の産生を減少させることができないことを明らかにしました[48]。 これら、およびここに提示された我々の結果は、経鼻投与されたHK L. plantarumがインビボで呼吸器抗ウイルス免疫応答の修飾を誘導する能力は、乳酸桿菌と気道の上皮細胞ではなく免疫細胞との相互作用に依存することを示している。 マクロファージおよび/または樹状細胞は、以前の研究[17、18、19]で示唆されているように、HK乳酸桿菌によって調節される細胞です。 これらの結果は、SARS-CoV-2に対する耐性を高めるために生存不能な乳酸桿菌を使用する可能性を排除すべきではないことを示しています。 invivoの結果は、上皮細胞培養で得られた結果とは異なる可能性があるためです。 したがって、HK L. plantarum MPL16またはCRL1506がSARS-CoV-2に対する免疫応答を調節する能力をinvivoで調査する必要があります。 生存不能な乳酸菌の使用は、生存可能な微生物の投与がリスクを表す可能性がある免疫不全患者の免疫力を向上させるための興味深い代替手段であるためです。 |
| 5. 結論 |
| 現在まで、SARS-CoV-2感染およびCOVID-19の結果に対する免疫調節プロバイオティクスの潜在的な有益な効果は、IFV、RSV、PVMなどの他のウイルス性病原体との関連で実施された研究から推定されています[7,8]。 私たちの知る限りでは、これは免疫調節性乳酸桿菌がSARS-CoV-2感染に対する耐性にプラスの影響を与える能力の最初の実証です。 この研究で提示されたinvitroの結果は、免疫調節性L. plantarumが呼吸上皮細胞の自然免疫応答を特異的に調節し、SARS-CoV-2感染に対する耐性に好影響を与えるという証拠を提供します。 COVID-19の予防または治療のためのL.plantarum MPL16またはCRL1506などの有益な免疫生物性乳酸桿菌の特定の菌株を特定するには、動物モデルにおけるSARS-CoV-2感染のさらなる機構研究およびinvivo実験が必要です。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
この文献は、Pathogens 2021, 10, 1197.に掲載されたImmunobiotic Lactobacilli Improve Resistance of Respiratory Epithelial Cells to SARS-CoV-2 Infectionを日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |