ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(発酵乳、腸内細菌の科学:研究の最前線) |
効率的にGABA(γ-アミノ酪酸)を産生するLactiplantibacillus plantarumで製造された豆乳ヨーグルトは、ストレプトゾトシン誘発糖尿病マウスの高血糖を改善し、腸内微生物叢を再構築します |
Brian Bor-Chun Weng et al., |
|
|
| 概要 |
豆乳ヨーグルトは、微生物を適切に操作して豆乳を発酵させるだけで作られるヴィーガン食品として人気を集めています。 イソフラボンを豊富に含む大豆が高血糖症の改善に効果があることはよく知られています。 大豆発酵は、これらの貴重な栄養素の生物学的利用能を向上させることができます。 Lactiplantibacillus plantarum は、豆乳製造において最も豊富で頻繁に分離される種の 1 つです。効率的に GABA (γ-アミノ酪酸) を産生する L. plantarum を使用して製造した豆乳ヨーグルトと L. plantarum の脱グリコシル化活性を、ストレプトゾトシン 誘発高血糖マウス モデルで機能的に評価しました。 高血糖マウスをグループに割り当てて、dH2O、豆乳、高GABA産生L. plantarum GA30(LPGA30)、低GABA産生L. plantarum PV30(LPPV30)で製造された豆乳ヨーグルト、または追加のGABA 対応物 (GA + GABA および PV + GABA グループ)30 mg g-1で強化された豆乳ヨーグルトのいずれかを毎日強制経口摂取させて処理します。 dH2O グループを除くすべての豆乳ヨーグルトグループは、グルコース恒常性、耐糖能試験結果、腎臓組織の完全性が改善され、体重が維持されました。 一方、豆乳グループには部分的な利点が示されています。毎日豆乳ヨーグルトを補給したグループ (LPGA30 および LPPV30) の血漿 GABA 濃度は、dH2O グループと豆乳グループの平均 0.5 μM の 5 倍高く安定しました。 また、GABA 強化豆乳ヨーグルトに相当するグループ (GA + GABA および PV + GABA) グループは、血漿インスリン レベルの回復に関与していました。 腸内マイクロバイオーム分析により、dH2O 群のストレプトゾトシン誘発高血糖マウスでは正常なFirmicutes門と Bacteroidetes門を超えて通性嫌気性Proteobacteriaが侵入した腸内毒素症(訳者注:腸内細菌叢異常)が明らかになりました。 過渡的に生息するActinobacteriaによる腸内微生物叢の回復は、LPGA30 グループでは実証されましたが、LPPV30 グループでは実証されませんでした。 効率的にGABAを生成するL. plantarum GA30で製造された豆乳ヨーグルトは、Enterococcus, Lactobacillusおよび Bifidobacterium,の優勢な属による腸内細菌叢の調節と、Akkermansia muciniphila, Butyricicoccus pullicaecorum, Corynebacterium spp. および Adlercreutzia spp.などのいくつかのマイナーな有益な微生物群集の存在において並外れた利点を示しました。 効率的にGABAを産生するL. plantarum GA30豆乳を発酵させて豆乳ヨーグルトを製造すると、豊富な大豆イソフラボンに対して深い相乗効果を示し、ストレプトゾトシン誘発性高血糖マウスのインスリン産生のための膵臓β細胞の機能を回復します。さらに、高血糖マウスの健康な腸内細菌叢の再構築におけるGABA産生L. plantarumのプロバイオティックな役割は、共生関係の可能性を示唆しており、さらなる調査が待たれている。 |
| 目次(クリックして記事にアクセスできます) |
| 1.序章 |
| 2.材料および方法 |
| 2.1.Lactiplantibacillus plantarum 株と GABA 合成の供給源 |
| 2.3.豆乳ヨーグルト中のイソフラボンの分析 |
| 2.4.動物実験 |
| 2.5.ストレプトゾトシン誘発性高血糖マウスモデルと治療群 |
| 2.6.血漿血糖値測定 |
| 2.7.γ-アミノ酪酸 (GABA) の測定 |
| 2.8.耐糖能検査 |
| 2.9.インスリン測定 |
| 2.10.腸内マイクロバイオームのメタゲノミクス解析 |
| 2.11.腎病理組織検査 |
| 2.12.統計分析 |
| 3.結果と考察 |
| 3.1.厳選されたLactiplantibacillus plantarum株で豆乳を発酵させ、イソフラボンアグリコンとGABAを濃縮 |
| 3.2.GABA強化豆乳ヨーグルトはストレプトゾトシン誘発性高血糖マウスの障害を改善した |
| 3.3.プラトー血漿 GABA は、効率的な GABA 生産L. plantarum または GABA 強化を無視した豆乳ヨーグルトの補給によって達成されます |
| 3.4.GABA強化豆乳ヨーグルトは、インスリン産生における膵臓β細胞の機能を最もよく保存します |
| 3.5.LPGA30 グループと LPPV30 グループにおける腸内微生物叢の多様性と豊富さの再確立の意味 |
| 4.結論 |
本文 |
| 1.序章 |
| 大豆の摂取は、I 型と II 型の両方の糖尿病のリスク低下と関連しており、糖尿病の症状を改善します(1-3)。 浸漬して粉砕した大豆を煮てろ過して作られた豆乳は、伝統的な中国の朝食のスープであり、豆腐やさまざまな大豆ベースの製品に開発できます。 豆乳は、皮膚保護、低コレステロール、抗高血圧、抗糖尿病、抗がん、老化防止などの栄養補助食品としての特性を示します(4,5)。 抗糖尿病の健康増進機能は、次の 4 つの化学カテゴリを含む植物エストロゲン性大豆イソフラボンすなわちアグリコン、グルコシド、アセチルグルコシド、およびマロニルグルコシドによって寄与されます(6)。 これらの中でも、アグリコン ゲニステインは、cAMP/PKA シグナル伝達経路を介して膵臓 β 細胞の代謝回転を促進することがよく実証されています(7)。 ダイゼインのより高い治療可能性についても検討されています(8)。それにもかかわらず、豊富な大豆イソフラボンはグリコシド結合体として天然に存在し、大豆残渣に保持されることが多いため、直接利用することはできません。また、適切に操作された微生物発酵により、その生物学的利用能が劇的に向上します(9)。 さらに、大豆オリゴ糖のグリコシド部分はプレバイオティクスの構成に似ています。 大豆オリゴ糖は腸内微生物叢を改善し、免疫力を高めることが実証されています(10)。 |
| 豆乳ヨーグルトは、最近人気を集めている新しい大豆ベースのヴィーガン製品です。 厳選された乳酸菌による豆乳発酵のみで製造されています。 微生物の嫌気性代謝による酸性化は、豆乳の豆乳ヨーグルトへの固化を助けます。 さらに、大豆発酵には、抗栄養素因子の減少、抗酸化能力の増加など、複数の栄養上の利点があり、腸内細菌叢による腸内消化のための微生物発酵プロセスと組み合わせることで、グリコシド結合を酵素的に加水分解してイソフラボンなどの栄養素の生物学的利用能を最適化するのに役立つ可能性があります。 豆乳ヨーグルトを使用した限定的な研究では、豆乳ヨーグルトの摂取によりラットの肝臓への脂質の蓄積を防ぐことができると報告されています(11)。さらに、L. plantarum発酵大豆製品は、高コレステロール食を与えたラットの脂質恒常性を改善することが実証されています(12)。プロバイオティクスとして、Lactobacillus spp.は、バクテリオシン生成を介して病原性E. coli, Salmonella typhimurium、その他のグラム陰性菌を阻害することで腸内細菌に影響を与えることができます(13,14)。 これらの研究は、腸の健康における潜在的な役割を示唆しています。 |
| 通常の食事からの GABA (γ-アミノ酪酸) 摂取不足が注意されています(15)。 それにもかかわらず、GABA は腸管内の乳酸菌種から固有に取得できます(16)。 実際、乳酸菌発酵食品は、GABA の主要な外因性供給源です。 神経化学物質としてのよく知られた役割に加えて、最近の研究結果は、抗糖尿病における GABA の重要な役割を裏付けています。 GABA は、膵臓の β 細胞塊に対して保護および再生効果を発揮します(17)。 さらに、Ben-Othman et al. (2017)は、ex vivo 培養物における持続的な 5 μM ~ 1 mM GABA 処理による GABA 受容体媒介膵臓 β 細胞増殖を実証しました(18)。 さらに、GABA には、血糖恒常性を維持するためのインスリンまたはグルカゴン分泌を介した膵臓の α 細胞から β 細胞への変換を積極的に制御する自己分泌機能もあります。 我々は最近、Rhizopus oligosporus と GABA を産生する L. plantarum を共接種して発酵させた大豆残渣基質において、発酵時間に依存して GABA 濃度が増加することを実証し、この発酵生成物は細胞内活性酸素種の減少に利点を示します。 高グルコース培養条件下でイヌ腎臓細胞を活性化し、毎日補給すると STZ 誘発高血糖マウスのグルコース恒常性を改善します(9)。 したがって、外因性の発酵食品源または固有の腸内微生物源の強化を仲介するプレバイオティクスを介した安定かつ十分な GABA 摂取は、健康に重大な影響を与える可能性があります。 それにもかかわらず、発酵大豆製品を介した GABA 摂取の重要性は調査が待たれており、微生物の GABA を介して発酵した大豆が血糖恒常性および健康な腸内細菌叢の調節に及ぼす影響はまだ十分に理解されていません。 本研究では、選択されたLactiplantibacillus plantarum属菌を用いた豆乳発酵によって製造された機能性豆乳ヨーグルトの健康増進への影響が明らかになった。 したがって、グルタミン酸を GABA に変換する高い効率を示し、脱グリコシル化活性をストレプトゾトシン誘発高血糖マウス モデルで調査しました。 |
| 2.材料および方法 |
| 2.1.Lactiplantibacillus plantarum 株と GABA 合成の供給源 |
| 効率的な GABA 生産株である L. plantarum GA30 株と低生産株である L. plantarum PV30 株が、伝統的な中国の豆腐製造から分離されました。 両方の分離株は、実験室での特性評価試験のために大豆ペプトンベースの培地(CAS 番号 91079-38-8、米国ミズーリ州シグマ)で日常的に培養され、生物資源収集研究センター(BCRC、台湾、新竹、ROC)に正式に寄託されています。 L. plantarum 株 BCRC 10069 は、2 つの分離株から GABA を生産するための最適な発酵条件を比較および設定するための参照標準として購入されました(19)。 基礎培地(BCRC の推奨に従って指定、ラクトバチルス MRS ブロス、DIFCO 0881)は、GABA 生成効果を評価するために、初期 500 mM の L-グルタミン酸ナトリウムで構成されました。 GABA の分析と透析精製については、後のセクションで説明します。 精製および凍結乾燥した細菌産生 GABA を、後続の食事性 GABA 強化豆乳ヨーグルト グループの添加物として使用しました。 |
| 2.2.豆乳ヨーグルト発酵 |
| 豆乳は商業的な豆腐生産者 (台湾の嘉義市) から購入し、滅菌した後、衛生的な条件下で室温まで冷却しました。 L. plantarum 株の対数期に近い細菌培養物 (109 CFU mL-1) を、5 L 嫌気性発酵槽内で 1% (v/v) の比率で豆乳に添加し、37 °C で 48 時間処理しました。 その後の一晩の冷却プロセス中に、最終的な発酵豆乳は自然に酸性化し、凝固した豆乳ヨーグルトになりました。 その後の動物実験のために、ヨーグルトのアリコートを凍結乾燥し、動物実験の準備が整うまで -80 °C の冷凍庫に保管しました。 |
| 2.3.豆乳ヨーグルト中のイソフラボンの分析 |
| イソフラボン含有量は、当社の以前の方法に従って高速液体クロマトグラフィー (HPLC) によって検出されました(9)。 簡単に説明すると、サンプルのアリコートの重量を量り、メタノール抽出のために Falcon チューブに入れました。遠心分離後、上清を回収した。 HPLC は、Li Chrospher 100RP-18e カラム (4 mm × 250 mm、5 μm)、移動相 0.05% TFA:CH3CN = 95:5_gradiant_60:66mm、および 280 nm の UV 検出器を使用して実行され、温度は 40℃に設定します。 流速 1.0 mL min-1 で実行時間 60 分の逆相カラムの特定の設定により、現在のイソフラボンのピークを検出できました。 保証チェックには内部標準が使用されました。 曲線の下の領域の特定のピークの定量化は、以前に確立された個別の化学検量線に対して実行されました。 6 つのイソフラボン、すなわちダイジン、グリシチン、ゲニスチン、ダイゼイン、グリシテイン、およびゲニステインを Sigma、(セントルイス、ミズーリ州、米国) から購入しました。 検出限界 (LOD) と定量限界 (LOQ) は、6 つの標準溶液の段階希釈によって評価され、その後 HPLC によって分析されました。 LOD と LOQ はそれぞれ 0.0305 μg mL-1 と 0.1221 μg mL-1 でした。 |
| 2.4.動物実験 |
| 現在の研究で実施された動物実験プロトコルは、中華民国台湾嘉義市の国立嘉義大学動物管理使用委員会によって承認番号103015で承認されました。 8 週齢の雄 C57/BL6 マウスを中華民国台湾国立実験動物センターから購入し、環境制御された特定の病原体のない施設で飼育しました。 動物に標識を付けて耳タグを付け、基本食(LabDiet 5001、米国)および蒸留水を自由に与えた。研究全体を通じて、体重を週に 2 回モニタリングしました。 |
| 2.5.ストレプトゾトシン誘発性高血糖マウスモデルと治療群 |
| 高血糖は、Hariyanto et al. (2022)の説明に従って誘発され、測定されました(9)。 簡単に説明すると、連続 5 日間の低用量 (体重 kg あたり 40 mg) ストレプトゾトシン (STZ; 10 mM クエン酸ナトリウム、pH = 4.5) の腹腔内注射を実行しました。 誘導に成功した高血糖マウスの空腹時血糖値は、250 mg dL-1 以上であると測定されました。 次いで、30 匹の高血糖マウスを 6 つの実験グループにランダムに割り当てました。 処理とサンプリングのスケジュールをスキーム 1 に示します。 全ての動物には、実験期間全体にわたって、朝に100μLの用量を毎日強制経口摂取させた。 モックコントロールグループには100μLのdH2Oのみを強制経口投与したことを除いて、 治療群には体重1kg当たり200mgの豆乳100μLを、豆乳ヨーグルトには体重1kg当たり200mgの豆乳100μLを強制栄養で与え、GABAを効率的に生産するL. plantarum GA30(LPGA30グループ)を使用した豆乳ヨーグルト、低GABA産L. plantarumPV30(LPPV30グループ)は、毎日強制経口摂取させた。 それぞれの豆乳ヨーグルトは、追加の 30 mg g-1 (乾燥ベースで w/w) GABA、GA + GABA グループ、および PV + GABA グループで強化されました。 効率的にGABAを生産するL. plantarum GA30(LPGA30グループ)で生産され、 低GABA産生L. plantarum PV30(LPPV30グループ)、GA + GABAグループとPV+GABAグループのそれぞれの豆乳ヨーグルトに、追加の 30 mg g-1 (乾燥ベースで w/w) GABA を強化した。 |
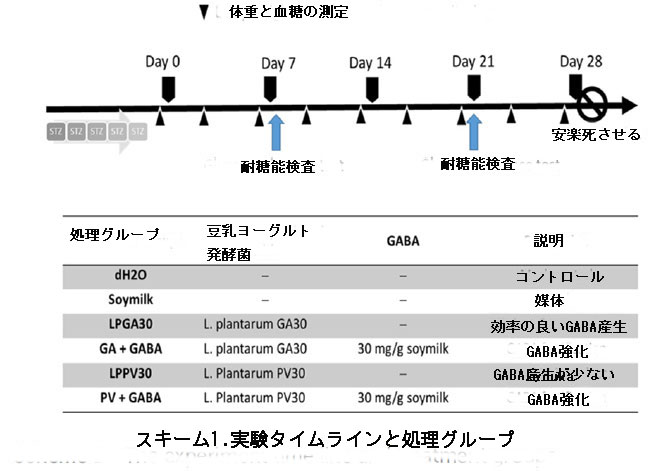 |
| 2.6.血漿血糖値測定 |
| 約 50 ~ 70 μL の血液サンプルを、週に 2 回、ヘパリン添加毛細管 (Chase、no. 2501、USA) を使用して伏在静脈の回転側から静脈穿刺で採取しました。 遠心分離後(Hettich Mikro 20 Centrifuge、Hettich、ドイツ)に血漿を得た。 血糖モニタリングシステム(Ascensia Elite® XL、Bayer、No. 1502、Basel、スイス)を使用して、血漿グルコースレベルを直ちに測定した。 |
| 2.7.γ-アミノ酪酸 (GABA) の測定 |
| GABA 濃度の測定は以前の方法から変更されました(9,20)。 血漿サンプルのアリコートを PBS で 10 倍に事前希釈しました。 続いて、メーカーの推奨に従って、高性能透析膜 (MWCO3500、Servapor®、Kelowna International Scientific Inc.、台湾) を使用して、希釈溶液の透析を 5 時間実行しました。 透析液をO-フタルアルデヒド(OPA、Sigma)と反応させてGABAを測定した。 50 μL のサンプルを HPLC 分析に使用しました。 LOD および LOQ は、標準の段階希釈によって評価されました。 LOD と LOQ はそれぞれ 33.7 nM と 112.6 nM でした。 |
| 2.8.耐糖能検査 |
| 耐糖能試験は、以前に報告された方法に従って 1 週目と 3 週目に実施されました(9,21)。 マウスを一晩絶食させ、グルコース溶液(体重1kg当たり0.5g)100μLを単回腹膜注射した。 直ちに、前述のように血液を 1 滴採取し、グルコース投与後 0、30、60、90、および 120 分後にグルコースレベルを測定しました。 血糖値は血糖モニタリングシステム(Ascensia Elite® XL、Bayer、No. 1502、Basel、スイス)を使用して直ちに測定されました。 |
| 2.9.インスリン測定 |
| 7日目に伏在静脈から採取し、28日目に安楽死エンドポイントの心臓穿刺から採取した血液サンプルを、ELISAキット(Mercodiaマウスインスリンカタログ番号10-1247-01、米国)を使用して血漿インスリンレベルについて測定した。 簡単に言うと、10μLの血漿を抗体でコーティングされた96ウェルプレートに加えた。 ELISA 手順はメーカーの推奨に従って実施されました。 HRP-TMB 反応の比色分析は、ELISA リーダー (Biotek Instruments, USA) を使用して OD450 nm で実行されました。 |
| 2.10.腸内マイクロバイオームのメタゲノミクス解析 |
| 約 2 cm の腸を無菌的に採取し、すぐにアルミホイルで包み、その後、プレスキーム 1で処理します。 実験タイムラインと治療グループは、さらなる分析まで-80 °C の冷凍庫で保管しました。 微生物叢分析では、解凍サンプルを 2 mL の PBS で内腔にフラッシュして洗浄液を収集しました。 同じグループのマウスから 3 つのサンプルをプールしました。 これらは、dH2O、LPGA30、およびLPPV30グループのSTZ誘発糖尿病マウスでした。 腸内マイクロバイオーム分析は、中華民国台湾の Qiagen Co. によって実施されました。 簡単に説明すると、プールした洗浄液サンプルのゲノム DNA をゲノム DNA 抽出キットを使用して抽出し、腸内微生物のプロファイリングを次世代シーケンス技術 (QIAseq および GeneRead Library Prep Kits、Qiagen、米国) を使用して実行しました。 正常なマウスの腸内微生物叢データは、同じ条件下で行われた兄弟実験から採用されました。 |
| 2.11.腎病理組織検査 |
| 腎臓を収集し、すぐに 10% ホルマリンに 3 日間浸し、その後、パラフィン包埋の前に 5 時間、4% ホルムアルデヒド溶液を含む 10% ホルマリンに変更しました。 次に、以前の方法に従って、パラフィン包埋腎臓組織の垂直断面を脱蝋し、ヘマトキシリンとエオシンで染色しました。 腎糸球体単位の完全性および慢性尿細管間質損傷の特徴の顕微鏡評価は、CCD 搭載顕微鏡を使用して実行されました。 |
| 2.12.統計分析 |
| データは表に示され、グラフ数値は平均±標準偏差として表されます。 データは、米国カリフォルニア州サンディエゴの Prism、GraphPad ソフトウェアを使用した二元配置分散分析によって分析されました。 それ以外の場合、治療比較は MS Excel Student の t 検定によって実行されました。 治療間の有意性は、有意水準 α = 0.05 でアスタリスクラベルとして表されました。 |
| 3.結果と考察 |
| 3.1.厳選されたLactiplantibacillus plantarum株で豆乳を発酵させ、イソフラボンアグリコンとGABAを濃縮 |
| 乳酸菌は、さまざまな発酵食品に含まれるプロバイオティクスの一般的な供給源です。 それらの中には、天然に GABA を生成するものもあれば、グルコシドイソフラボンをアグリコンに遊離させるなど、グリコシル化フェノール化合物を効率的に触媒するものもあります。 表 1 に、さまざまな L. plantarum 株によって生産された豆乳および豆乳ヨーグルトのグリコシル化および脱グリコシル化イソフラボン、総イソフラボン、および GABA 含有量を示します。 イソフラボンの大部分は天然では多糖類と結合しているため、その生物学的利用能が制限されています。 豆乳では、グリコシル化イソフラボンの小計 (3985 ± 7.6 μg g-1) はアグリコン (609 ± 8.1 μg g-1) の 6 倍を超えました。 LPGA30、LPPV30、またはBCRC10069のいずれかのL. plantarum株で発酵すると、イソフラボンアグリコン(ダイゼイン、グリシテイン、ゲニステイン)の濃度が明らかに増加し、グリコシル化イソフラボン(ダイジン、グリシチン、ゲニステイン)は減少しました。 それにもかかわらず、効率的にGABAを産生する株LPGA30は、他の株と比較して相対的に非効率的である。 さらに、総イソフラボンのわずかな減少は、48 時間の発酵中の生分解に起因する可能性があります。オリゴ糖結合大豆イソフラボンの利用(吸収)は主に、アグリコンに遊離するβ-グルクロニダーゼなどの微生物の加水分解性酵素に依存しています。豆乳摂取後のヒト血漿中のダイジンまたはゲニスチンは高感度質量分析法では検出されなかったことが報告されています(23)。対照的に、ダイゼインとゲニステインは血漿中で 1 時間以内にピークに達し、12 時間高濃度のままでした。 同様に、発酵豆乳をマウスに与えた場合、現在のマウスモデルではダイゼインとゲニステインの優先的な血漿取り込みが明らかでしたが、脱グリコシル化型グリシテインは検出されませんでした(HPLCクロマトグラフィーによるESIデータ†)。 |
| GABA は主に発酵食品から摂取できる食物であり、腸内の GABA 生成微生物によって容易に合成されます。 関連する健康への影響において、プレバイオティクスおよびプロバイオティクス GABA 源の生物学的利用能に注目が集まっています。 48 時間の豆乳発酵により、乾燥基準レベル 460 ±2.8 μg g-1、530 ± 4.4 μg g-1、および 67 ± 2.1 μg g-1 で GABA 含有量が異なって上昇しました。それぞれ L. plantarum 株 BCRC10069、GA30、および PV30 によるものです (表 1)。 また、豆乳からはGABAは検出されませんでした。 最近の研究 (24,25 )では、L. plantarum spp. を使用して、操作された培養条件と伝統的な発酵食品の実践の下でGABAが豊富な製品を生産することも実証されています。 乳製品ヨーグルトでは、最適化された条件下 (L-グルタミン酸一ナトリウムおよびピリドキサール-5-リン酸を含む)、L. plantarumNDC75017 は、牛乳 100 g あたり 315 mg という高い GABA 含有量をもたらしました(26)。 現在、効率的な GABA 生産または脱グリコシル化を示す L. plantarum 株 GA30 および PV30 が分岐して選択されました。我々の以前の結果は、選択された L. plantarum GA30 による 48 時間のグルタミン酸の GABA への変換が最大化され、最終生産量 8 log CFUmL-1 細菌と 5000 mM 以上の GABA 生産に達したことを実証しました(19)。 現在の培養条件下では、L. plantarum GA30による豆乳発酵により、L. plantarum PV30よりも8倍近いGABAが生成されます(新鮮な豆乳ヨーグルト100 gあたり53 mg)。ただし、乳製品ヨーグルトはヴィーガン豆乳ヨーグルトよりも優れています。 それにもかかわらず、L. plantarum PV30 は、発酵に 3 倍多くのイソフラボン アグリコンを寄与することにより、L. plantarum GA30 よりもイソフラボン脱グリコシル化の効率が高いことを示しています。 Tamang et al (2016) は、豆乳とその発酵製品の保護機能は、栄養素の利用を促進し、食欲を促進し、抗酸化活性を高め、腸内微生物叢を強化することにより、発酵食品の複数の実際的な特性と関連している可能性があると報告しています(27)。 以前、我々はまた、豆乳製造由来の大豆残渣に Rhizopus oligosporus と GABA を産生する Lactiplantibacillus plantarum を同時接種することで、残留イソフラボンの利用を最大限に高め、リサイクルにより付加される健康機能の価値を実証しました(9)。今回の結果は、選択された L. plantarum 株で発酵させた豆乳が GABA の効率的な生産、またはイソフラボン アグリコン含量の増加を示したことを示しています。効率的な GABA 生成または脱グリコシル化を示す、選択された L. plantarum 種の分岐を使用して製造された 2 つの豆乳ヨーグルトの抗高血糖特性は、さらなる研究に値します。 |
 |
| 3.2.GABA強化豆乳ヨーグルトはストレプトゾトシン誘発性高血糖マウスの障害を改善した |
| グルコース利用の調節不全の影響は、酸化損傷、慢性炎症、長期にわたる高血糖状態による体重減少を引き起こす可能性があります。 図 1A に示すように、dH2O グループは、長期にわたる高血糖状態下で最後の実験期間 (20 ~ 28 日) に体重減少が見られました。 一般に、豆乳または豆乳ヨーグルトを毎日強制栄養補給すると、LPPV30 グループの研究終了時に体重が減少したことを除いて、体重減少が防止されました。 さらに、ストレプトゾトシン誘発性高血糖症は、すべてのグループの血漿グルコース レベルの上昇で証明されました。(図1B)。 毎日の豆乳補給は、dH2O グループと比較して血糖恒常性において中程度の利点を示しましたが、豆乳ヨーグルトグループとその GABA 強化対応物はすべて、豆乳および dH2O グループと比較して上昇する血漿グルコースを有意に (P < 0.05) 低下させました。 興味深いことに、GA + GABA および PV + GABA グループの追加の GABA 強化は、LPGA30 および LPPV30 グループに比べて無視できる程度の改善を示し、統計的有意性が得られました。それにもかかわらず、この結果は、血糖恒常性における大豆イソフラボンの重要な役割を実証しています。 さらに、微生物の発酵プロセスは、グリコシドフェノール成分を解放するだけでなく、イソフラボンの生物学的利用能を高めることができ、抗酸化状態の上昇により、高血糖状態に関連する酸化負担や損傷を引き起こすストレプトゾトシンの第一選択の保護が促進される可能性があります(9)。 L. plantarum PV30 で製造された豆乳ヨーグルトは GA30 よりも 3 倍高いアグリコン含量を持っていたため、LPPV30 グループのより高い抗酸化状態が予想されました (表 1)。 さらに、1週目と3週目に行った耐糖能試験では、単回ブドウ糖注射によりdH2O群では血糖値が30分でピークに達したが、他の群では血糖恒常性の制御不良が示されなかったことが実証された(図1C)。 驚くべきことに、豆乳の補給は、豆乳ヨーグルトおよびその追加の GABA 添加グループで見られたのと同じくらい効果的でした。 それにもかかわらず、豆乳グループの1週目では60分でピーク血糖値が遅延し、3週目では30~90分でピーク血糖値が延長したことは、L. plantarum株による豆乳発酵による追加の利益を暗示しました。 最後に、図 1D に示すように、腎糸球体組織切片の保護機能も組織病理学的に評価されました。 慢性高血糖は腎障害を引き起こすことがよくあります。 腎皮質糸球体の焦点領域の組織病理学的検査では、dH2O グループで被膜基底膜の肥厚と不規則なエッジ (矢印で示す) が見つかりましたが、他の治療グループと比較して豆乳グループでは顕著ではありませんでした。 さらに、糸球体毛細血管は、GA + GABA および PV + GABA グループではコンパクトであり、あまり拡張していませんでした。 一般に、LPGA30 および LPPV30 グループでは、経時的腎損傷はあまり目立たず、GABA 強化対応グループである GA + GABA および PV + GABA グループは、拡大した被膜基底腔や不規則なエッジの明らかな兆候がなく、さらに優れた保護機能を備えているようです。 これは、GABA が豊富な発酵大豆残渣とイソフラボンで保護された腎臓の完全性に関する以前の発見と一致しています。 選択された L. plantarum 株で製造された大豆ヨーグルトは、高血糖合併症の予防において全体的に期待される結果を示し、大豆イソフラボンの利点に加えて、GABA の相乗的保護の存在が認められています。 |
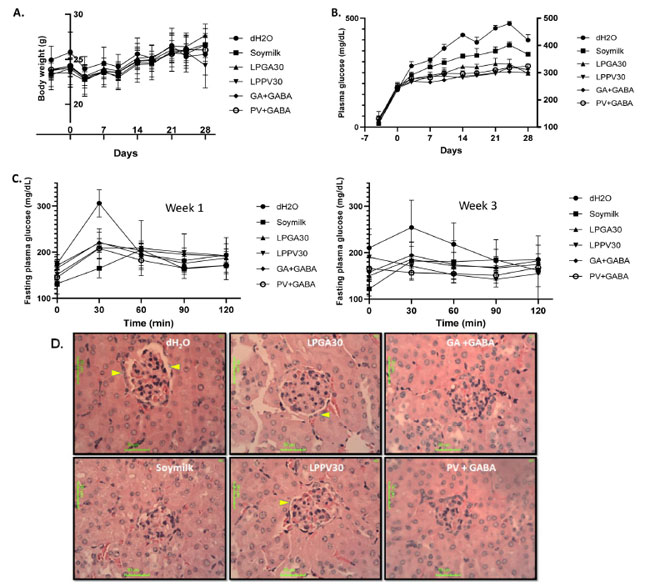 |
| 図1 体重変化の評価 (A)、全実験期間にわたる血漿グルコースレベル(B)、1週目と3週目の耐糖能試験、およびdH2O、豆乳、豆乳ヨーグルトを毎日補給したストレプトゾトシン誘発高血糖マウス群の糸球体の腎組織病理学(D) LPGA30 および LPPV30、それらの GABA 強化対応物である GA + GABA および PV + GABA グループ。 |
| 3.3.プラトー血漿 GABA は、効率的な GABA 生産L. plantarum または GABA 強化を無視した豆乳ヨーグルトの補給によって達成されます。 |
| 乳酸菌発酵食品からの食事性 GABA は、通常の食物不足を補うために重要です(15)。 抑制性神経伝達物質として、血漿 GABA は血液脳関門を通過できません。 血漿 GABA はマイクロモル以下の範囲で安定したレベルに維持されており、ヒトの炎症への反応においては年齢に依存しません(28)。 重要なことに、GABA 投与は、非肥満糖尿病マウス モデルの膵島における膵臓 β 細胞の増殖と下方制御された自己免疫性 T 細胞媒介細胞毒性を促進することが実証されています(29)。 さらに、GABA はさまざまな末梢組織、特に膵臓内分泌機能の仲介に存在します。 実際、それは血漿グルコースと相関して膵臓内で固有に合成され、膵臓のα細胞およびβ細胞の変換を調節します(18)。 図 2 に示すように、毎日 100 μL dH2O または豆乳を強制経口摂取したストレプトゾトシン誘発高血糖マウスの平均血漿 GABA レベルはそれぞれ 0.46 および 0.66 μM でした。 一方、豆乳ヨーグルトまたは GABA 強化豆乳ヨーグルトのサプリメントでは、血漿 GABA レベルが 5 倍 (約 2 ~ 3 μM) 高い濃度に大幅に上昇しました。 効率的な GABA 生産者である L. plantarum GA30 によって生産された豆乳ヨーグルトの血漿 GABA レベルは、低 GABA 生産者である L. plantarum PV30 よりわずかに高いだけです (P >0.05)。 さらに、追加の GABA (30 mg g-1 豆乳ヨーグルト) を供給された GA + GABA および PV + GABA グループは、対応する豆乳ヨーグルトである LPGA30 および LPPV30 グループと比較して、血漿 GABA レベルに有意な差を示さなかった。 これらの結果は、通常の食事(マウスの餌)には GABA が不足しており、豆乳の補給は血漿 GABA レベルに影響を及ぼさないことを示しています。 4 週間の実験期間にわたって毎日サプリメントを摂取した後、豆乳ヨーグルトグループとその GABA 強化対応グループは、血漿 GABA 濃度がプラトー濃度に達しているようです。 Gerber と Hare (1980)30 は、カニューレを挿入した健康なげっ歯類の平均血漿 GABA が約 0.58 μM であると報告しています。 これは、マウスにおけるストレプトゾトシン誘発膵臓 β 細胞破壊の現在の所見と類似しています。 正常マウスおよび高血糖マウスでは、血漿 GABA が低レベルのままであると推測できます。 したがって、現在の GABA 強化豆乳ヨーグルトを長期的に摂取すると、おそらく生理学的限界に達する最適な GABA 摂取が得られる可能性があります。 Li et al (2015) は、薬物動態および薬力学の研究において、健康なヒト被験者の循環インスリンレベルの増加と相関して、血漿 GABA が厳密に制御された恒常性を示したと報告しました(31)。末梢のGABA取り込みと安定したインスリンレベルに関連するその恒常性をさらに解明するには、より詳細な研究が必要です。 |
| それにもかかわらず、低GABA産生L. plantarum種による豆乳発酵が期待されなかった。 特に、L. plantarum PV30 の GABA 合成効率は、インビトロでの L. plantarum GA30 のほぼ 8 分の 1 です (表 1)。 血漿 GABA の上昇が L. plantarum spp. による豆乳発酵と深く関連していると仮定するのは容易に合理的かもしれません。 サイレントGABA合成遺伝子が微生物のコミュニケーションを通じて再活性化されるのか、プラトー血漿GABAに起因する高血糖状態での代償性GABA吸収によって再活性化されるのかについては、さらなる解明が必要である。 さらに、多様に選択された L. plantarum 株を使用して製造された豆乳ヨーグルトのシンバイオティクス (プレバイオティクス + プロバイオティクス) 特性も、さらに調査される興味深いものです。 それにもかかわらず、GABA は膵臓 β 細胞塊および膵臓 β 細胞変換のシグナル伝達において再生効果を発揮することがわかっています(17)。 現在のストレプトゾトシン誘発高血糖マウス研究では、追加の GABA (30 mg g-1 ヨーグルト) 強化を無視して、豆乳ヨーグルト 1 kg あたり 200 mg を毎日強制経口投与すると、GABA は容易に血漿中に取り込まれます。 GABA 強化により、血漿 GABA レベルのさらなる増加は限定的となりました。 プラトー血漿 GABA レベルが、インスリン産生における膵臓 β 細胞機能の回復を予測する上で重要であるかどうかは、以下で説明されます。 |
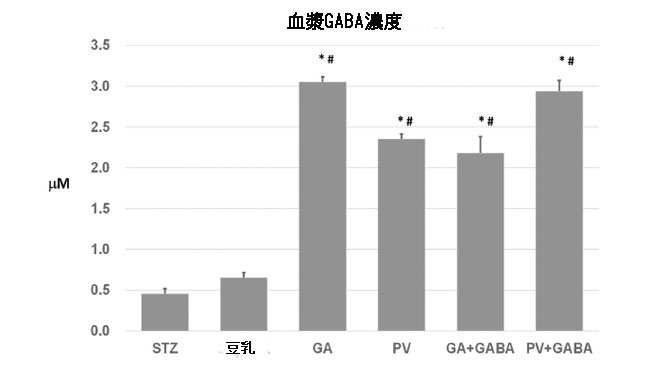 |
| 図2 dH2O、豆乳、豆乳ヨーグルト、GABA 強化豆ヨーグルトを補給したストレプトゾトシン誘発高血糖マウス(STZ)の血漿GABA濃度 P = 0.05 での有意性。* と # はそれぞれ dH2O グループと豆乳グループとの比較で示されます。 |
| 3.4.GABA強化豆乳ヨーグルトは、インスリン産生における膵臓β細胞の機能を最もよく保存します |
| 1週目と4週目の終わりに測定された血漿インスリンレベルを図3に示します。すべてのグループにおけるインスリンレベルの一般的な低下は、破壊された膵臓β細胞によるストレプトゾトシン誘導性高血糖に起因する血糖調節異常を裏付けた。dH2O グループでは、血中インスリン レベルの劇的な低下が明らかでした。最初の週では、LPPV30 および PV + GABA グループは、dH2O グループよりもインスリン産生の傾向が高く (P < 0.1)、β 細胞機能がより良く保存されているようです。L. plantarum PV30 は、GABA 合成における有効性は低いですが、L. plantarum GA30 と比較して、イソフラボン アグリコンの生成において効率的な脱グリコシル化剤です。 グルコシド大豆イソフラボンは、そのグルコース部分が発酵プロセス中または腸内微生物叢によって容易に除去されて遊離イソフラボン アグリコンになるまで、ほとんど吸収されません。したがって、L. plantarum PV30によるグリコシル化フェノール化合物の遊離は、大豆イソフラボンのより高い生物学的利用能と、ストレプトゾトシンによって引き起こされる酸化損傷の非常に初期段階でβ細胞を保護するのに十分な抗酸化物質を意味します。実際、グリコシル化イソフラボンは健康な成人の腸細胞を通って吸収されず、腸内マイクロバイオームがイソフラボンの生物学的利用能に重要な役割を果たしているため、アグリコンイソフラボンの腸吸収にはβ-グルコシダーゼ活性が必要です(23,33)。 また、豆乳由来のイソフラボン配糖体のβ-グルコシド結合は、Bifidobacteriumが産生するβ-グルコシダーゼおよびβ-ガラクトシダーゼにより加水分解されることが報告されています(34)。さらに、4 週目の終わりには、すべての治療グループの血漿インスリン レベルが dH2O グループの血漿インスリン レベルよりも著しく高かった。 驚くべきことに、豆乳グループは、LPGA30 グループおよび LPPV30 グループと同様の血漿インスリンレベルを示しました。血漿 GABA は豆乳グループでは影響を受けなかったので (図 2)、腸内細菌叢に対する豆乳単独のプレバイオティクス特性と、豊富な大豆イソフラボンが膵臓 β 細胞インスリン産生の保護を仲介することが証明されています。これは、豆乳グループにおける血漿グルコースレベルの中間制御に関する以前の結果を裏付けています(図1A)。さらに、GA + GABA グループと PV + GABA グループはどちらも平均して血漿インスリン濃度が高く、dH2O グループよりも有意に (P < 0.05) 高い値でした。 豆乳ヨーグルト由来の GABA は、膵臓 β 細胞に保護効果と再生効果を与える上で重要な役割を果たしている可能性があります。Soltani et al., (2011) は、GABA がα細胞の GABA 受容体と反応し、グルカゴン分泌の抑制において膜過分極を引き起こす可能性があることを明らかにしました(17)。さらに、毎日の腹腔内(マウスあたり20μmol)GABA注射による80日間連続の治療効果も実証し、ストレプトゾトシン誘発糖尿病マウスにおいてβ細胞量の回復、血糖恒常性の改善、および糖尿病の回復を示した。 したがって、GABA 状態を長期にわたって安定して維持することは、グルコース恒常性を維持し、さらには糖尿病を治療するためにも重要です。GABAを強化した豆乳ヨーグルトは、糖尿病の高血糖症状の改善に期待できる機能性食品です。 結論として、脱グリコシル化とGABA合成に優れた選択されたL. plantarum株を用いて豆乳発酵によって製造された豆乳ヨーグルトは、膵臓β細胞のインスリン産生に相乗効果をもたらし、糖尿病を改善する。 |
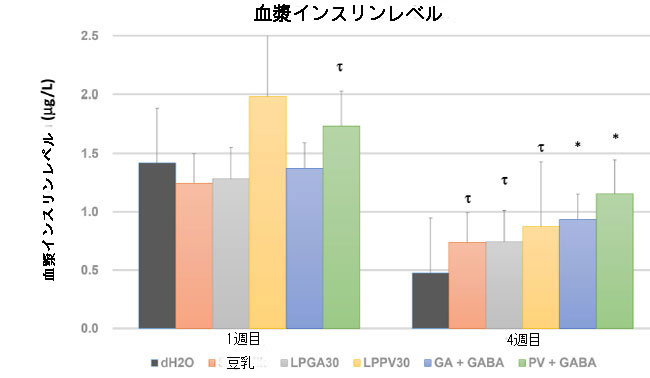 |
| 図 3 1 週目と 4 週目に測定したインスリンレベル。*、P < 0.05、および τ、P < 0.1 で dH2O グループと比較して有意性が得られました。 |
| 3.5. LPGA30 グループと LPPV30 グループにおける腸内微生物叢の多様性と豊富さの再確立の意味 |
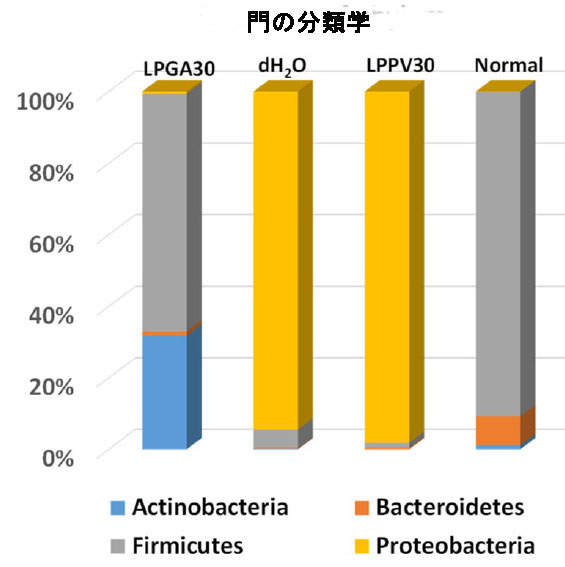
| 腸内マイクロバイオーム解析により、健康で正常なC57/BL6マウス(ストレプトゾトシン誘発性高血糖マウス)の腸内で優勢な細菌集団の系統学的プロファイルが明らかになった LPGA30 群と LPPV30 群のマウスには、それぞれ L. plantarum GA30 または L. plantarum PV30 で生成された異なる豆乳ヨーグルトが補給されました。 腸内微生物叢の門レベルで異なるグループを図 4 に示します。 |
 |
 |
| 図 4 異なる治療群およびハウスキーピング対照正常マウスの腸内マイクロバイオーム分析における門の分類。 |
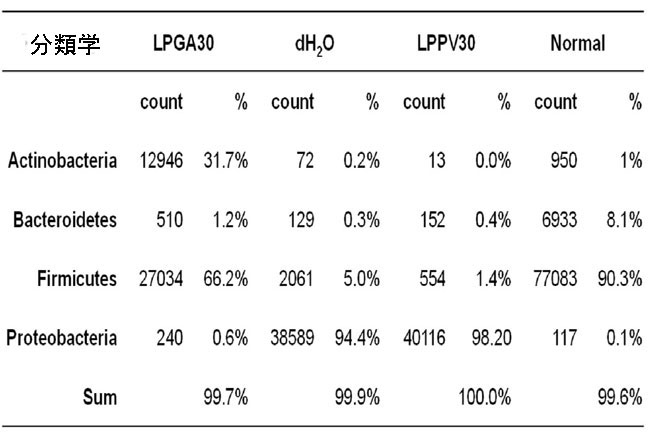
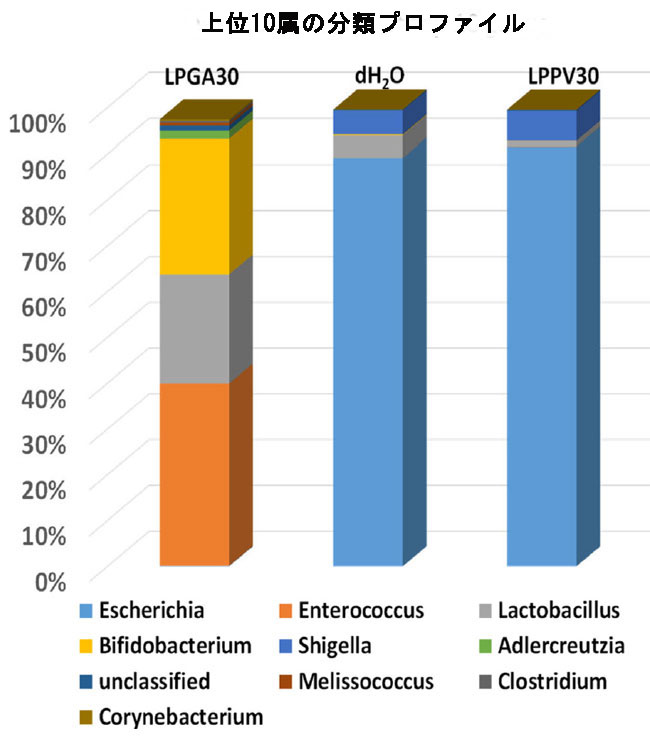
| 一般に、Actinobacteria, Bacteroidetes, Firmicutes およびProteobacteriaが、標準的な LabDiet 飼料で飼育されている現在の動物の細菌の 99% 以上を占めています。 ハウスキーピング対照コロニーの正常な健康マウスではFirmicutes門 (90.3%) とBacteroidetes門 (8.1%) が優勢でしたが、ストレプトゾトシン誘発高血糖マウスにdH2Oを毎日強制摂取した場合(dH2O群)、またはL. plantarum PV30で製造した豆乳ヨーグルト(LPPV30群)にはProteobacteriaが過剰に存在していた(それぞれ94.4%と98.2%)。 さらに、Firmicutes属とBacteroidetes属は、dH2O グループではわずか 5% と 0.3%、LPPV30 グループでは 1.4% と 0.4% に劇的に減少しました。 Proteobacteriaの増殖は腸内毒素症の特徴であり、腸細胞の完全性の破壊によって媒介される腸上部の酸素利用可能性と pH レベルの変化は、通性嫌気性Proteobacteriaが偏性嫌気性Firmicutesに置き換わる環境を促進します。 しかし、効率的なGABA生産者であるL. plantarum GA30(LPGA30グループ)が生産する豆乳ヨーグルトを毎日強制経口摂取させると、減少したFirmicutes属とBacteroidetes属がそれぞれ66.2%と1.2%に回復することが実証されています。 さらに、Proteobacteriaの個体数はわずか 0.6% であるのに対し、Actinobacteriaの量は 31.7% と顕著でした (図 4)。 腸内微生物叢の腸内毒素症により、宿主の脂質とグルコースの代謝が変化する可能性があります(36)。 実際、腸内細菌叢は宿主のエネルギー配分と利用において重要な役割を果たしており、腸内細菌叢の動的な変化は宿主のグルコース恒常性を反映している可能性があります。 それにもかかわらず、Patterson et al., (2015) は、単回高用量ストレプトゾトシン誘発糖尿病ラットが、Bacteroidetes/Firmicutes比の変化により腸内微生物叢に劇的な影響を 5 週間示し、全体的な腸内微生物の多様性を減少させ、Lactobacillus および Bifidobacterium属の割合が増加したと報告しました(37)。 現在の研究は、低用量のストレプトゾトシンを 5 日間連続して毎日投与した高血糖誘発マウスは、4 週間後に安楽死させ、腸内微生物叢の多様性を有し、優勢な属はEscherichia および Shigellaであることを実証しました (dH2O グループ)。 一方、LPGA30 グループの優勢な属は、Firmicutes門とActinobacteria門のEnterococcus, Lactobacillus および Bifidobacteriumの移行集団でした (図 5)。 |
 |
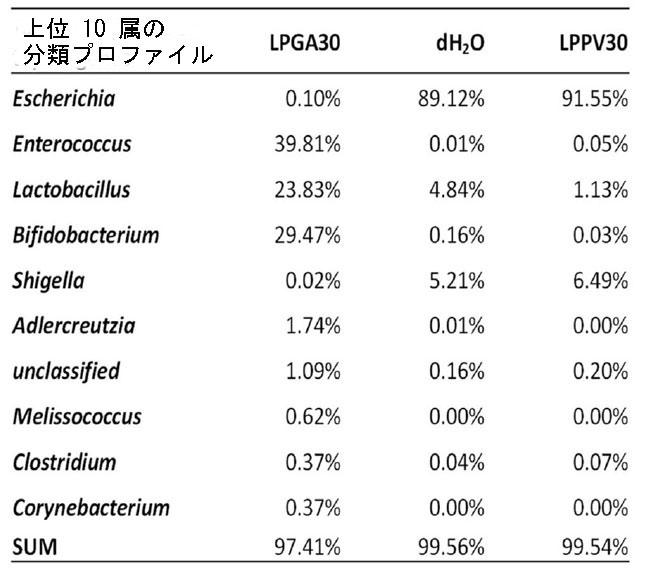 |
| 図 5 さまざまな治療グループの腸内マイクロバイオーム分析における上位 10 属の分類プロファイル |
| 一部の L. plantarum 株は、糖の輸送と利用のための遺伝子発現を調節することにより、その居住地でいくつかの宿主因子と相互作用できることが報告されています。 さらに、高脂肪食環境における最近の研究では、GABA を産生する L. brevisを 8 ~ 10 週間経口投与すると、メタボリック シンドロームとうつ病が改善されました(39)。 彼らの GABA 産生 L. brevis は、Firmicutes門とBacteroidetes門 (盲腸含有量) の存在量や比率を変化させなかったということは、言及する価値があります(39)。 現在の研究とのこの不一致は、現在のマイクロバイオーム分析が上部腸管 (十二指腸) で行われたためである可能性があります。 腸内細菌叢の組成は、ストレプトゾトシン誘発糖尿病ラットの十二指腸、回腸、および結腸における細菌叢の変化のパターンを伴って、腸領域特異的な様式で発生します(40)。 さらに、LPGA30 群と LPPV30 群の両方でプラトー血漿 GABA レベルが見られたが、ストレプトゾトシン誘発高血糖マウスに投与された効率的な GABA 生成 L. plantarum GA30 生成豆乳ヨーグルトのみが、腸上部のFirmicutesに次いで2番目に主要な集団であるActinobacteria門 (一次Bifidobacterium属) を含む微生物叢のバランスが一時的に再調整されたことは言及する価値がある。 実際、上部腸の収縮能の異なる抑制性または興奮性の調子は、異なる GABA 受容体アイソフォームの分布に依存します。 GABA の存在は消化管の運動性に影響を及ぼし、腸内細菌叢の多様性と豊富さによる微生物の定着を引き起こす可能性があります。 Auteri et al., (2015)41 は、腸内 GABA 受容体媒介シグナル伝達を介した GABA 作用が胃腸管の運動性と抗炎症活性に影響を与えると記載しました。 最後に、Akkermansia muciniphila, Butyricicoccus pullicaecorum,Corynebacterium spp. および Adlercreutzia spp.を含むいくつかの少数かつ有益な腸内微生物集団は、L. plantarum GA30生成豆乳ヨーグルトを与えたマウスでは観察されたが、L. plantarum PV30 または dH2O グループではほとんど観察されなかった (データは示されていません)。 これらの微量細菌は腸の pH 値、炭素と窒素の供給源、微生物叢のメンバー間の共生関係に影響を与える他の成長因子を変化させる可能性があります。 GABA との調整を十分に考慮しながら、宿主の消化生理学と相互扶助する宿主のエネルギー利用や腸内細菌叢の動的な変化をさらに評価することは興味深いでしょう。 |
| 4.結論 |
| 豆乳ヨーグルトは、GABA 合成と脱グリコシル化する潜在能力を持つ厳選されたL. plantarum属菌を操作することにより、豆乳発酵という簡単なプロセスで健康機能を促進する大豆発酵食品として人気を集めています。 現在、L. plantarum GA30 または L. plantarum PV30 で製造された豆乳ヨーグルトを与えられた高血糖マウスで実証されているように、最適化された血漿 GABA レベルと大豆イソフラボンの生物学的利用能の向上により、複数の利点が証明されています。 GABA とイソフラボンを豊富に含む豆乳ヨーグルトは、相乗的に、ストレプトゾトシン誘発糖尿病マウスの高血糖の改善において膵臓 β 細胞機能の回復を介して血糖恒常性を改善することが期待されています。 さらに、腸内毒素症における高血糖の影響が明らかになりました。 効率的にGABAを生成するL. plantarum GA30を補給して製造された豆乳ヨーグルトは、Actinobacteria門が一時的に存在する腸内微生物叢を再構築することができ、豆乳ヨーグルトのさらなる健康上の利点をもたらします。 宿主と微生物の共生オーケストレーションに向けたプレバイオティクスとプロバイオティクスの相互作用をさらに解明するには、より詳細な研究が必要です。 |
参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
この文献は、Food Funct., 2023, 14, 1699–1709に掲載されたSoy yoghurts produced with efficient GABA (γ-aminobutyric acid)-producing Lactiplantibacillus plantarum ameliorate hyperglycaemia and re-establish gut microbiota in streptozotocin (STZ)-induced diabetic mice. を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |