ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(発酵乳、腸内細菌の科学:研究の最前線) |
脳の発達と神経変性疾患におけるマイクロバイオームの役割 |
Varsha Nandwana et al., |
|
|
概要 |
| 何千億もの共生微生物が私たちの体の内外に住んでおり、そのほとんどは生後すぐに腸に定着し、残りの人生の間そこにとどまります。 動物モデルでは、中枢神経系と腸内微生物叢 (腸-脳軸) との間の双方向通信が広く研究されています。 また、微生物叢(マイクロバイオーム)の組成の変化が、自閉症スペクトラム障害、アルツハイマー病、パーキンソン病、多発性硬化症、筋萎縮性側索硬化症、不安、ストレスなど、さまざまな神経発達障害および神経変性障害の病因に重要な役割を果たしていることは明らかです。 微生物叢の構成は、遺伝学、健康状態、分娩方法、環境、栄養、運動、および脳機能の維持における腸内微生物叢とその代謝物の役割に関する現在の理解など、さまざまな要因の影響を受けます。 前述の神経疾患の発症は、この総説記事にまとめられています。 さらに、神経疾患に対処するためのプロバイオティクス、プレバイオティクス、およびシンバイオティクスの使用における現在の飛躍的な進歩についても説明します。 さらに、記憶におけるホウ素ベースの食事の役割、ホウ素と微生物叢の関係、抗炎症剤としてのホウ素、神経変性疾患におけるホウ素についても議論しました。 さらに、今後数年間で、ホウ素試薬は 腸内毒素症(dysbiosis) を改善するために重要な役割を果たし、研究者に新しい領域を開くでしょう。 |
| 目次(クリックして記事にアクセスできます) |
| 1.はじめに |
| 2.腸内細菌叢 |
| 2.1. 分類と特徴付け |
| 2.2.腸内細菌叢の構成の変化 |
| 2.3. 私たちの体における腸内細菌叢の重要な機能と有益な効果 |
| 2.4.腸内細菌叢とその代謝物 |
| 3.腸-脳軸 |
| 4. 脳の発達における腸内細菌叢の役割 |
| 5. 神経発達疾患における腸内細菌叢 |
| 5.1 自閉症スペクトラム障害(ASD) |
| 6. 神経変性疾患における腸内細菌叢 |
| 6.1. アルツハイマー病 |
| 6.1.1. 腸内毒素症とアルツハイマー病 |
| 6.1.2.アルツハイマー病に関与する代謝物 リポ多糖 (LPS) |
| 6.1.3.漏れやすい腸と漏れやすい脳 |
| 6.2.パーキンソン病 |
| 6.3.多発性硬化症(MS) |
| 6.4.筋萎縮性側索硬化症(ALS) |
| 7. プレバイオティクス、プロバイオティクス、シンバイオティクス、サイコバイオティクスのサプリメントが腸内細菌叢と関連疾患に及ぼす影響 |
| 8. 神経保護剤としてのホウ素 |
| 8.1 食事中のホウ素 |
| 8.2 ホウ素と腸内細菌叢 |
| 8.3 抗炎症剤としてのホウ素 |
| 8.4 神経変性疾患におけるホウ素 |
| 8.5 その他の神経保護剤 |
| 9. まとめと今後の展望 |
| (注1)略語 |
| (注2)菌名の読み方 |
1.はじめに |
|
人間の体には、ひとまとめにしてヒト微生物叢として知られる何十億もの小さな生き物が生息しており、それらのゲノムは微生物叢(マイクロバイオーム)と呼ばれています。 「忘れられた臓器」として知られる腸内微生物叢には、およそ 300 万個の遺伝子があり、これはヒトゲノムの最大 150 倍です [1]。微生物は私たちの皮膚だけでなく、泌尿生殖器、胃腸、呼吸器系でも繁栄しますが、胃腸管は最も密集しています。胃腸 (GI) 管の末端に位置する結腸と直腸は、人体で最も多くの細菌を収容していると考えられています [2]。驚くべきことに、私たちの腸内微生物叢の 3 分の 1 だけがほとんどの個人に共有されていますが、残りの 3 分の 2 は、個人の ID カードと同様に各人に固有のものです [3]。この微生物集団の構成は時間の経過とともに変化し、外部および内因性の両方の変動を受けます [4]。食事、代謝、年齢、場所、ストレス、および抗生物質療法はすべて、有用な共生生物と潜在的に病原性の微生物との間のバランスを崩す可能性があります。「腸内毒素症」[5] は、この崩壊した均衡を表す言葉です。腸内微生物叢は、免疫機能と代謝バランスの維持、ビタミン生成、病原体防御、血管新生の刺激、および腸バリアの維持において重要な役割を果たしていることが示されています。腸内微生物叢が恒常性を維持し、中枢神経系 (CNS) を含む実質的にすべての主要な身体系を調節する役割を果たしているという認識は、過去 20 年間に生物医学に反乱を引き起こしました [6]。 「腸-脳軸」(GBA)は、腸内微生物と中枢神経系の間の双方向通信経路の存在を意味し、現在広く受け入れられています [7]。 この軸の調節不全は、自閉症スペクトラム障害、アルツハイマー病、多発性硬化症、パーキンソン病などの神経障害の病態生理学に関与していることがますます疑われています [6]。 現在、プレバイオティクス、プロバイオティクス、シンバイオティクスなどの微生物叢に基づく治療法や微生物叢の糞便移植は、代謝とメンタルヘルスを改善するために真正生物を促進することを目的としています [8]。さらに、活性生物微量元素の一種であるホウ素 (B) は、神経保護効果を与える必須栄養素であることが示唆されています。ホウ素の摂取は、骨、ミネラル、脂質の代謝、および免疫機能に関連しています。ビタミン B が人間の健康にとって重要であるという証拠が増えているため、ビタミン B の栄養素摂取と脳および心理機能との間の関連性を調査することが重要です [9]。このレビューの主な目標は、さまざまな神経発達疾患および神経変性疾患における腸内微生物叢の機能、およびその構成の変化について現在知られていることを要約することです。さらに、これらの病気を改善するために採用されてきた数多くの治療法についても説明します。 |
2.腸内細菌叢 |
| 2.1 分類と特徴付け |
| 腸内細菌叢は、属、科、目、および門によって分類された細菌です。 Firmicutesと Bacteroidetes は最も一般的な腸内微生物門であり、総微生物叢の 90% から 95% を占めており [10]、腸内のすべての細菌の遺伝物質としても説明されています。 Clostridium (Firmicutes門の 95%)、Blautia,Faecalibacterium, Enterococcus, Lactobacillus Eubacterium, Roseburium, および Ruminococcusは、Firmicutes門を構成する 200 を超える属の 1 つです。 Bacteroidetes は、Bacteroides や Prevotella などの優勢な属で構成されています。 腸内微生物叢の他の門には、Proteobacteria, Actinobacteria, Fusobacteria, Spirochaetes, および Verrucomicrobiaが含まれます[11]。 |
| 研究者は、16S rRNA、18S、および ITS 遺伝子の DNA 抽出と配列決定に基づく新しい方法 [13] を使用して、腸内微生物叢の構成要素を系統学的に同定および定量化することができました [12]。 16S rRNA 遺伝子 (1500 bp) は、情報科学の目的には十分な大きさであり、ほとんどすべての細菌に見られ、その機能は時間の経過とともに変化していないため、最も一般的に使用されている系統発生ツールでした [14,15]。メタゲノミクスは、細菌叢の現在の分類法に関する豊富な情報を提供しますが、重要な機能についてはほとんど提供しません。メタトランスクリプトミクスは、細菌叢によって発現される遺伝子の機能分析を研究し、微生物集団の分類学的構成を評価する可能性があるRNA ベースの手法です。メタプロテオミクスは、多生物システムの集団タンパク質組成の研究であり、微生物群集からのタンパク質の大規模な同定と定量化は、分子レベルでの微生物の表現型への直接的な洞察を提供します [16]。 メタボロミクスは、生体試料中の代謝産物の体系的な分析であり、疾患の根底にある代謝障害の特定、バイオマーカーの開発など、さまざまなレベルでの代謝表現型の詳細な表現型と精密医療を可能にします。 これは、疾患の診断や治療活動の監視に使用できるほか、新しい潜在的な治療法の発見にも使用できます [17]。 |
| 2.2.腸内細菌叢の構成の変化 |
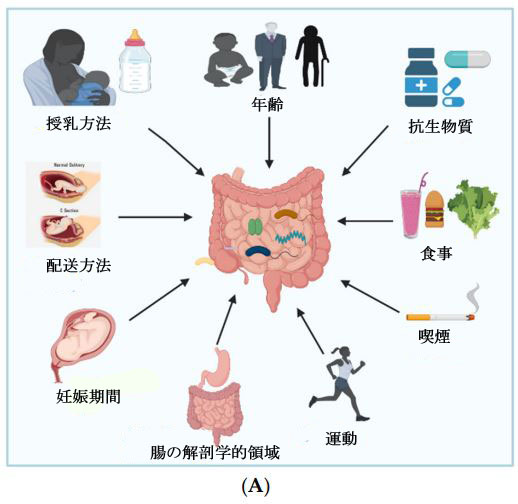
| 腸内細菌叢の構成は動的であり、腸の解剖学的領域、妊娠期間、分娩の種類、母乳育児の技術、離乳期間、年齢、抗生物質の使用、民族性、食習慣、文化的慣行など、さまざまなパラメーターの影響を受けます (図. 1A) [18]。 細菌の複雑さと量は、消化管を下るにつれて成長する傾向があり、胃では数が少ないが、結腸では非常に高濃度である[19]. Lactobacilli, Veillonella, および Helicobacterは胃で最も一般的な細菌であり、Bacilli, Streptococcaceae, Actinomycinaeae, およびCorynebacteriaceaeは十二指腸、空腸、回腸で典型的な細菌です。 結腸内のLachnospiraceae と Bacteroidetesの濃度は、回腸を下るにつれて増加します[19]。 出生時の妊娠期間は、腸内微生物のコロニー形成における重要な要因です。 早産児 (37 週以前に生まれた) の腸内微生物叢は、正期産児とは異なります。 早産児における微生物のコロニー形成は、限られた微生物多様性、Enterobacter, Enterococcus, Escherichia, および Klebsiellaなどの潜在的に病原性および通性嫌気性細菌の数の増加、およびBifidobacterium, Bacteroides, および Atopobium などの偏性嫌気性菌のレベルが低いことによって特徴付けられます[ 20]。 経膣分娩で生まれた乳児の微生物叢は、Bifidobacterium longum と Bifidobacterium catenulatum および Escherichia coli、Staphylococcus、Bacteroides fragilis、Streptococcus などの他の通性嫌気性種で構成されています。 一方、帝王切開で生まれた乳児は、母体の皮膚微生物叢に似た優勢な種としてStreptococciを持ち、防御的な Bifidobacterium longum subspecies infantis (B. infantis)細菌のレベルが低い[21]。 母乳で育てられた新生児は、粉ミルクで育てられた乳児よりも一貫した均一な微生物叢の集団を持ち、2 倍の Bifidobacterium spp.を保有し、Clostridium およびStreptococcus 菌種の数が少ない[22]。 |
 |
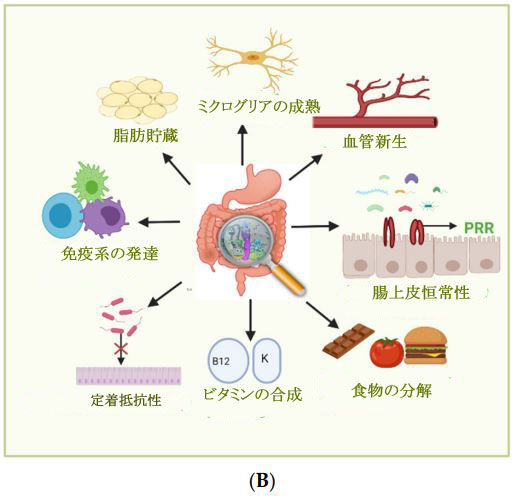 |
| 図 1 (A) 腸内細菌叢の変化を引き起こす要因。 (B) 腸内細菌叢の重要な機能 |
| 人間の腸は、出生時に微生物学的に無菌ではありません。 実際には、細菌のコロニー形成は子宮内で始まる多次元的な手順であり、分娩方法や摂食方法などのさまざまな要因の影響を受けます。1歳までに、子供の微生物叢の構成には、Akkermansia muciniphila, Bacteroides, Veillonella, および Clostridium botulinum spp.が特徴的に豊富に含まれています。 成人期には、Firmicutes (Lachnospiraceae and Ruminococcaceae), Bacteroidetes (Bacteroidaceae, Prevotellaceae, および Rikenellaceae), および Actinobacteria (Bacteroidaceae, Prevotellaceae, および Rikenellaceae)という 3 つの細菌門が優勢であり、人が年をとるにつれて、Bifidobacterium およびFirmicutesのレベル 減少する傾向があり、Bacteriodetes および Proteobacterium のレベルが上昇します[12]。 食事内容は、腸内微生物群に大きな影響を与えます [23]。高脂肪食は、Bifidobacteriaを含む、腸内のグラム陰性菌およびグラム陽性菌の数の減少に関連しています [24]。 Zimmer らの研究によると、完全菜食主義者は対照患者よりもBacteroides, Bifidobacterium, およびEnterobacteriaceae spp.のレベルが低いことを示しました。 [25]。全粒穀物およびプレバイオティクスが豊富な食事は、Enterobacteriaceae および Desulfovibrionaceae,を含む日和見感染症を減少させる一方で、腸バリア保護菌として機能するBifidobacteriaceaeを増加させることが実証されています [26]。抗生物質は感染症との闘いにおいて不可欠なツールですが、抗生物質のクラス、投与量、曝露時間によっては、その使用によって細菌の多様性と数が減少する可能性があります.分娩中の抗生物質は、Bacteroidetesを減少させながら、新生児の腸内のBacteroides と Enterobacteriaの数を増やすことができます[27]。さらに、運動不足や肥満、ストレス、喫煙などの食事以外のライフスタイル変数も、有益な腸内細菌叢を低下させます [28]。 |
| 2.3 私たちの体における腸内細菌叢の重要な機能と有益な効果 |
| 宿主の免疫系の発達、活性化、機能はすべて、微生物叢の影響を受けます。 次に、宿主の免疫系は、微生物叢との共生相互作用を維持するためのいくつかのメカニズムを進化させてきました [29]。 |
| 自然免疫系は、物理的および化学的障壁、免疫細胞、および血液タンパク質 (サイトカインなど) で構成されており、私たちの防御の最前線です。 共生細菌と病原性細菌のこの識別検出は、小腸の上皮細胞とリンパ系細胞の膜に見られる Toll 様受容体 (TLR) によって媒介されます。 Toll 様受容体は、ペプチドグリカン成分 (ムラミン酸、フラジェリン、莢膜多糖、リポ多糖) などの多様な細菌抗原を含む複数の微生物関連分子パターン (MAMP) を識別し、自然腸免疫を活性化します。 NOD 様受容体 (NLR) は、さまざまな微生物固有の化学物質を識別し、ダメージ関連パターン センサーとして機能するインフラマソームの形成を引き起こします。 小腸粘膜の特殊な分泌細胞であるパネート細胞も、抗菌ペプチド (AMP) を生成することにより、腸内微生物叢の構造を定義する上で重要な役割を果たします。 免疫受容体は、パターン認識受容体 (PRR) として知られており、病原体と共生生物を検出し、微生物と宿主に対して異なる結果をもたらす二重の役割を果たします [30]。 |
| 自然免疫とは異なり、獲得免疫は特定の病原体に対して高度に特異的であり、B 細胞と T 細胞として知られる 2 つの主要なリンパ球集団によって媒介されます。 獲得免疫システムは、CD4+ T 細胞に大きく依存しています。 ナイーブ CD4+ T 細胞は、刺激を受けた後、T ヘルパー 1 (Th1)、Th2、Th17、または制御性 T 細胞 (Treg) の 4 つのサブタイプのいずれかに発生する可能性があります。 各系統は、分化後に固有のサイトカインを分泌します [31]。 Bacteroides fragilis は、その多糖分子を介して、細胞内感染を根絶するために重要な全身の Th1 応答を誘導します。 一方、Segmented Filamentous Bacteria(セグメント細菌)は、Th17 細胞の効果的なインデューサーであることが示され、Clostridiaは、機能不全が自己免疫疾患につながる可能性のある免疫寛容の重要なメディエーターである結腸御性 T 細胞 (Treg)の産生を増加させることがわかっています。 腸内マイクロバイオームは、CD8+ T 細胞が他の末梢免疫細胞 (限界帯 B 細胞、形質細胞様樹状細胞、不変のナチュラル キラー T 細胞など) を訓練することによって影響を与えるのを助けます。 パイエル板には腸関連 B 細胞が存在しますが、腸内微生物叢は粘膜 IgA 合成の重要な推進力です。 共生細菌と可溶性抗原は、分泌型 IgA (SIgA) によってコーティングされており、宿主の上皮に付着して固有層に侵入するのを防ぎます。 その結果、分泌型 IgA (SIgA)は腸のバリアとして機能し、宿主と微生物叢の間の相利共生関係の維持に役立ちます [32]。 |
| それらはコロニー形成抵抗性、すなわち、共有資源と生息地をめぐって競合すること、および宿主防御プロセスを強化することによって、外因性病原体感染からの保護を提供します[33]。 腸上皮には微生物成分を特定するさまざまなパターン認識受容体 (PRR)が備わっており、その結果、細胞増殖を促進する細胞シグナル伝達経路を活性化するため、それらは上皮の恒常性をサポートします[34]。 腸内微生物叢は、局所の微小血管細胞を活性化することにより、血管新生 [35] と脂肪蓄積を刺激することもできます。 それらは神経系にも影響を与え、ナイーブミクログリアの発達に寄与し、微生物叢が存在しないと成熟ミクログリアの数が減少します. また、食物分子を分解し、ビタミンやその他の栄養素を合成し、骨量密度を変更し、医薬品を活性物質に代謝します [36]。 |
| 2.4 腸内細菌叢とその代謝物 |
| さまざまな細菌の属および種が、宿主と協力して、さまざまな代謝産物の産生に関与しています。 大腸では、小腸での消化と吸収を逃れる難消化性オリゴ糖が腸内微生物叢によって発酵され、酢酸、プロピオン酸、酪酸 (>95%) などの短鎖脂肪酸 (SCFA) が生成されます。 ギ酸、吉草酸、カプロン酸、またはその他の成分が残りを占めています [37]。 ほとんどのプロピオン酸は肝臓で代謝されるため (Koh et al., 2016)、酢酸は循環器系で最も一般的な 短鎖脂肪酸であるのに対し、酪酸は結腸の健康を維持するために上皮粘膜で主に代謝されます (van der Beek et al.,2017)。 Firmicutes は酪酸の主な生産者であるのに対し、Bacteroidetes は酢酸とプロピオン酸の主要な生産者です [38]。 短鎖脂肪酸は宿主細胞に運ばれ、G タンパク質共役受容体 GPR41、GPR43、および GPR109A と相互作用します。これらは代謝物を感知し、腸上皮および免疫細胞で発現します。 その結果、腸や他の臓器の恒常性を維持するために重要なプロセスが誘導されます [39]。 2 番目のメカニズムは、HDAC1 や HDAC3 などの核クラス I ヒストン脱アセチル化酵素 (HDAC) の細胞内直接抑制に関連しています。 核クラス I ヒストン脱アセチル化酵素の阻害は主に、炎症誘発性サイトカイン (IL-6、IL-8、TNF-α など) の減少や NF-kB 作用の減少など、抗炎症性免疫表現型に関連しています。 短鎖脂肪酸は、クローディン 1、オクルディン、小帯オクルデンス 1 などの密着結合タンパク質を調節することによって、上皮バリアの完全性を維持することもできます (Wang et al., 2012)。 したがって、これらのタンパク質のレベルが低下すると、細菌とリポ多糖類(LPS)の移行が促進され、炎症反応が促進されます。 短鎖脂肪酸は、結腸細胞の分化と増殖、粘膜細胞の移動、ムチン2 発現の増強、酸化ストレスの調節、および免疫応答においても役割を果たします (van der Beek et al., 2017)。 これらはすべて、パーキンソン病やクローン病などの病気に対する人体の闘いに不可欠です [38]。 さらに、短鎖脂肪酸は代謝された後、いくつかの臓器に送達することができます。たとえば、プロピオン酸は主に糖新生に関与していますが、酢酸と酪酸は主に脂質生合成に関与しています (図 2) [38]。 |
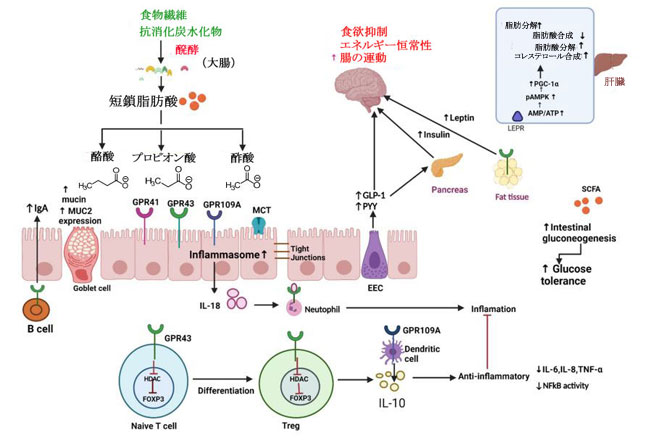 |
| 図 2 腸内細菌叢とその代謝産物の概略図 |
| 大腸では、食物繊維と未消化の炭水化物が腸内細菌叢によって 短鎖脂肪酸 (酪酸、プロピオン酸、酢酸) に発酵されます。これらの代謝産物は、タイトジャンクションを調節し、ムチン産生を増加させ、MUC-2 発現をアップレギュレートし、B 細胞の活性化を通じて IgA の合成を増加させることができます。 これらの代謝産物は、HDAC (ナイーブ T 細胞から制御性 T 細胞への分化につながる) を阻害することにより、抗炎症効果も発揮します。 これにより、IL-6、IL-8、TNF-α、NFkB 活性などの炎症性サイトカインの産生がさらに減少します。SCFAはまた、腸の糖新生を刺激し、耐糖能を改善します。 さらに、SCFAは EEC を刺激して GLP-1 と PYY を放出させ、食欲を抑制することで食欲抑制剤として作用し、GLP-1 は膵臓を刺激してインスリンを放出させ、筋肉と脂肪組織へのグルコースの取り込みを増加させます。 SCFAは、脂肪組織のレプチン合成を増加させることによって食欲を抑制します。 肝臓のSCFAでは、AMP/ATP 比を増加させることによって直接的に、または LEPR を介して間接的に AMPK をリン酸化し、活性化します。 AMPK の活性化は、脂肪酸の酸化、脂肪分解、およびコレステロール合成を促進し、脂肪酸合成を減少させる PGC-1α 発現を引き起こします。 略語: SCFA、短鎖脂肪酸。 MUC2、ムチン2; HDAc、ヒストンのアセチル化。 EEC、腸内分泌細胞。 GLP-1、グルカゴン様ペプチド 1; PYY、ペプチドYY; pAMPK、リン酸化アデノシン一リン酸活性化プロテインキナーゼ。 AMP、アデノシン一リン酸。 ATP、アデノシン三リン酸。 LEPR、レプチン受容体。 PGC-1α ペルオキシソーム増殖因子活性化受容体ガンマ コアクチベーター 1-アルファ。 |
| トリプトファンは重要な代謝産物の前駆体であり、重要な芳香族アミノ酸です。 宿主細胞では、オートムギ、魚、牛乳、チーズなどの食物源からのトリプトファンは、キヌレニン [40,41] またはセロトニン [42] の 2 つの経路のいずれかを取ることができます。 3 番目の経路では、腸内微生物がトリプトファンからインドールやその誘導体を含むさまざまな化合物への直接変換に関与しています (図 3 および図 4) [43]。 Clostridium perfringens および Escherichia coli は、アリル炭化水素受容体に結合するインドールアセチルグリシン、インドキシル硫酸、インドール-3-プロピオン酸、6-硫酸、およびセロトニンなどのインドール誘導体を生成します。 したがって、腸-脳軸に関与する炎症性および抗炎症性遺伝子の産生を調節することにより、結腸の完全性を維持するのに役立ちます [44,45]。 トリメチルアミン-N-オキシド (TMAO)、トリメチルアミン、アグマチン、ヒスタミンなどの生体アミンは、ヒスタミン受容体に作用し、腸上皮恒常性、細胞増殖、および老化に関与しています。抗炎症および抗腫瘍効果を調節します。 それらはClostridium saccharolyticum、Campylobacter jejuni、Bifidobacteriumによって産生される[38]。 胆汁酸 (Bas) は、肝細胞によって産生されるコレステロール由来の小分子です。 腸内微生物叢は、一次胆汁酸 (ケノデオキシコール酸とコール酸) を二次胆汁酸に変換し、それらを脱抱合します [43]。 これらの胆汁酸は、主に G タンパク質共役胆汁酸受容体 1 を標的とし、グルコース、コレステロール、およびエネルギーの恒常性を調節し、腸のバリア機能を維持し、脂溶性ビタミンの吸収を促進し、炎症性遺伝子の NF-kB 依存性転写を阻害します。 ビタミンB9、チアミン、ビタミンB2、ナイアシン、ビタミンK、ビタミンB1、リボフラビンなどのビタミンは、Bifidobacterium bifidum, Bacillus Subtilis, Escherichia coli, Bacteroidetesなどの細菌によって産生され、免疫機能、細胞代謝、細胞増殖、および宿主にビタミン源を提供します。 Clostridium, E. coli, Salmonella, および Bacteroidesによって産生されるポリフェノールは、抗酸化物質として機能し、結腸がんや炎症のリスクを軽減します [38,46,47]。 |
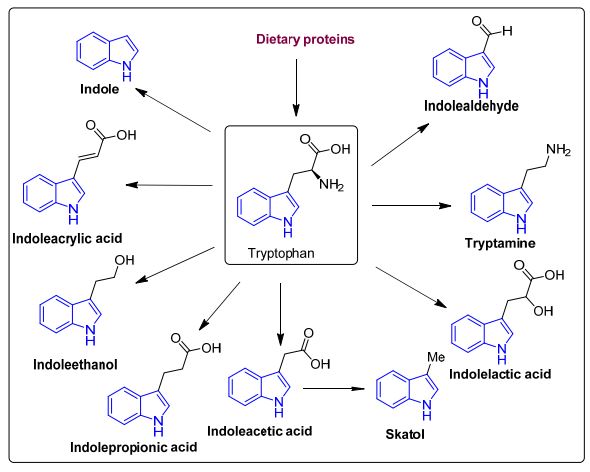 |
| 図 3. トリプトファン異化産物、インドール、およびその誘導体の構造 |
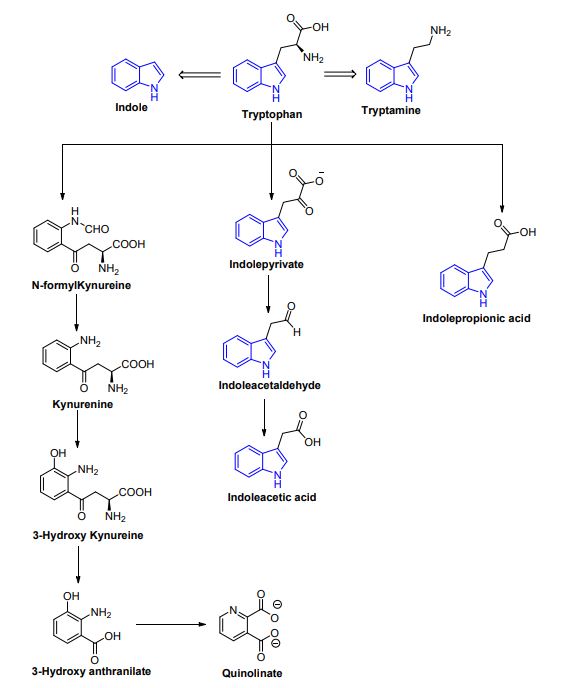 |
| 図 4. 神経活性化合物の産生に至るトリプトファン代謝経路の模式図 |
| 3.腸-脳軸 |
| 腸-脳軸 (GBA) は、双方向通信によって特徴付けられます。つまり、腸内微生物叢は脳にメッセージを送信し、脳は神経経路 (迷走神経および腸神経系)、内分泌経路 (コルチゾール)、免疫経路 (サイトカイン)、および体液経路を介して腸にシグナルを送信します [48]。 腸内分泌細胞と消化管腸クロム親和性細胞、腸粘膜関門、血液-脳関門を含む神経内分泌系と神経免疫系は、主にボトムアップのコミュニケーションに関与している[49,50]。 短鎖脂肪酸、トリプトファン代謝産物、および二次胆汁酸は、他の腸内微生物叢代謝産物の中でも、このボトムアップ コミュニケーションの重要なメディエーターです。 神経解剖学的経路、腸バリアの調節、および神経伝達物質 (5-HT やカテコールアミンなど) の放出はすべて、脳と腸内細菌間のトップダウンのコミュニケーションに関与しています [51,52]。 2 つの神経解剖学的経路が腸と脳を接続します。 1 つ目は、脊髄の迷走神経と自律神経系 (ANS) を介した腸と脳の間の直接的なコミュニケーションを反映しています。 2 つ目は、腸の腸神経系と脊髄の迷走神経および自律神経系との間の双方向の接続です [53]。 |
| 視床下部-下垂体-副腎 (HPA) 軸は、ストレッサーに対する適切な反応を確保するために、感染症などの心理的および身体的ストレスに反応して多くの生理学的機能を調節する重要な神経内分泌系です [54]。 室傍核からの副腎皮質刺激ホルモン放出ホルモン (CRH) の放出は、視床下部-下垂体-副腎軸の活性化を知らせ、続いて脳下垂体前葉からの副腎皮質刺激ホルモン (ACTH) の放出を誘導します。 副腎皮質刺激ホルモンは副腎皮質でグルココルチコイド (ヒトのコルチゾール) の生成を刺激し、これは腸の生理機能 (例えば、腸上皮バリアと免疫反応の変更) と腸内微生物叢の組成に大きな影響を与える可能性があります [55]。 免疫細胞 (樹状細胞、B 細胞、マスト細胞、および T 細胞) によるケモカインおよびサイトカインの産生などの免疫応答の活性化は、腸または他の場所で脳に影響を与えます。 腸内微生物叢は、ガンマアミノ酪酸、5-HT、ドーパミン、短鎖脂肪酸など、脳を含む人体に影響を与えるさまざまな重要な神経伝達物質を生成します [53]。 |
| 4. 脳の発達における腸内細菌叢の役割 |
| 腸内細菌叢は、他の神経発達過程とともに、ミクログリアの成熟と機能、血液-脳関門の形成と安定性、髄鞘形成、神経新生に影響を与えることがわかっている [56]。 無菌マウスのおかげで、初期の脳の発達における微生物叢の関連性が発見されました。 Diaz Heijtz と同僚 (2011) は、無菌マウスが、小脳との長期シナプス増強、ステロイド ホルモン代謝、および環状アデノシン 5 リン酸媒介シグナル伝達を含む、さまざまな可塑性および代謝経路に関連する遺伝子のアップレギュレーションを有することを発見しました。 ゲノムワイドなトランスクリプトミクスアプローチを使用して、海馬が最も影響を受けています[57]。 成体海馬の神経新生は、背側海馬の無菌マウスで促進されますが (Ogbonnaya et al., 2015)、抗生物質治療により神経新生が減少し、これはプロバイオティクスと運動で回復できます (Mohle et al., 2016)。 ドーパミン作動性および 5-HT 神経伝達物質の代謝回転の増加、ならびにシナプス形成マーカーの上昇が、無菌マウスの線条体で発見されており、運動および探索行動の変化に寄与している [58]。 Luczynski et al., 2016 は、正常な微生物叢を持つマウスと比較した場合、無菌マウスは海馬と扁桃体の体積が増加し、基底外側扁桃体に樹状突起肥大があることを示しました (社会的行動パターンと同様に不安と恐怖関連の反応を媒介します) [59]。 無菌マウスでは、脳の前頭前皮質部分で髄鞘形成が亢進し、髄鞘形成に関与する遺伝子の発現が上昇していた (Hoban et al., 2016) [57,60]。 Lu et al (2018) は、無菌マウストランスファウネーションパラダイムを利用して、出生後の脳の発達に対する高成長または低成長の表現型を引き起こすことが知られている早産新生児微生物叢の影響を調査しました。 高成長表現型に関連する微生物叢、神経細胞マーカーNeuN および血液バイオマーカー(ニューロフィラメントL)ならびに髄鞘形成マーカーMBP と比較すると、低成長表現型に関連する微生物叢は、これらのマーカーの低下を示しました。 さらに、Nos1の増加、循環および脳IGF-1の低下、循環IGFBP3の低下を含むIGF-1経路の変化によって示されるように、成長不良の表現型に関連する微生物叢は神経炎症の増加と関連していた[61]。 |
| 5. 神経発達疾患における腸内細菌叢 |
| 人間の脳は妊娠 3 週目から成長を始め、生まれる頃には 860億個の神経細胞と 100 兆個の接続があり、基本的な回路が作られています。 環境の影響下で、これらの初歩的な回路はますます複雑なリンク回路に進化します。 オキシトシンなどのホルモン、免疫系、セロトニンなどの神経伝達物質、微生物叢 - 腸 - 脳軸はすべて、社会的認知、感情、および行動を支える神経回路に寄与しています。 これらの社会的、認知的、行動的次元、およびそれらの神経発達は、自閉スペクトラム症(ASD) や統合失調症などの神経発達障害で損なわれます。 病気の素因がない人でも、「病原性」マイクロバイオームは病気を引き起こすのに十分な場合があります (Kim et al., 2017)。 たとえば、うつ病、過敏性腸症候群に関連する不安症、または統合失調症の患者から得た微生物叢を野生型マウスに移植すると、適応症に特有の行動異常が促進されました (De Palma et al., 2017; Zheng et al., 2016, 2019 ) [62,63]。 |
| 5.1 自閉症スペクトラム障害(ASD) |
| 自閉症は、社会的コミュニケーションの困難、興味の欠如、反復行動を特徴とする発達障害です [64]。 アメリカ疾病予防管理センター(CDC)の自閉症および発達障害モニタリング (ADDM) ネットワークからの推定によると、自閉症の割合は世界中で大幅に増加しており (有病率は 1%) [56]、54 人に 1 人の子供に影響を与えています [65]。 しかし、自閉スペクトラム症の特定の病因と病態生理を特定することは困難であり、効果的な治療法はほとんどありません。遺伝的および環境的変数、異常な免疫応答、および腸内細菌叢による腸の完全性の崩壊はすべて、自閉症の発症に関連しています[66]。 出生前の栄養、周産期のストレス、帝王切開、早産、母乳育児の制限、感染症、および抗生物質の使用は、自閉スペクトラム症に関連する環境リスク要因の1つです[67,68]。 食物拒絶、食物アレルギー、便秘-下痢、食物不耐症、胃の不快感、およびうるさい食習慣はすべて、一般集団よりも自閉スペクトラム症の子供に多く見られます[69]. 現在の証拠 [70,71] によると腸内細菌叢の変化は、自閉スペクトラム症を含む神経発達障害に関連しています。 便秘は、自閉スペクトラム症の子供でより一般的であることが示されており[72]、腸バリアの完全性を損なう可能性のある微生物の共生に関連している可能性があります[73]。 |
| Srikantha らによると、バリア形成タイトジャンクションの損失によって生じる腸透過性は、自閉スペクトラム症の病態生理学におけるバイオマーカーの可能性がある [74]。 いくつかの調査では、自閉スペクトラム症患者の血清、脳組織、および脊髄液中の IL-6、IL-1、TNF-α、および TGF-β のレベルが高いことがわかっている [75]。 Ashwood らは、自閉スペクトラム症患者には炎症誘発性化学物質循環が高く、調節循環が低いという仮説を支持し、これは粘膜免疫病理学の発生をさらに裏付けています。 コントロールと比較して、CD3+ T細胞は、自閉スペクトラム症の子供の十二指腸と結腸でより豊富に観察されました[76,77]。 自閉スペクトラム症の病態生理学における GMB 代謝産物、特に短鎖脂肪酸の重要性は、近年研究者の関心を集めています。 これらの短鎖脂肪酸は、クエン酸サイクルとカルニチン代謝の点でミトコンドリアの役割に影響を与え、自閉スペクトラム症関連遺伝子をエピジェネティックに変化させると考えられている [78]。 最も顕著な 短鎖脂肪酸の 1 つである酪酸 (BT) は、神経保護剤として推奨されており、ミトコンドリアの活動にプラスの影響を与えることが示されています。 核クラス I ヒストン脱アセチル化酵素 (HDAC)を阻害することにより、酪酸は血液脳関門 (BBB) を調節し、腸の炎症誘発性マクロファージの活動を抑制します。 その結果、酪酸を産生する細菌分類群は、自閉症の人では数が少ない [79]。 プロピオン酸は、自閉スペクトラム症を持つ人々に問題を引き起こす方法で消化管機能を混乱させます (Getachew et al.) [80]。 自閉スペクトラム症動物モデルで見られるものと同様の、可逆的な神経炎症、代謝、行動、およびエピジェネティックな変化を誘発する可能性があります。 Altieri らによると、大量の尿 p-クレゾールは自閉スペクトラム症のような反復行動に関連している [81]。 正常なマウスでは、尿毒症毒素であり GMB 代謝産物である 4-エチル フェニル サルフェート (4-EPS) は、自閉スペクトラム症様の行動と不安様の症状を引き起こします。 無菌マウスは 4-EPS を検出せず、これが GMB 代謝産物であることを示しています。 4-EPS レベルは、自閉スペクトラム症特性を持つマウスモデルで示されているように、母体免疫活性化 (MIA) の子孫でより高いことがわかりました。 自閉スペクトラム症の母体免疫活性化モデルでは、プロバイオティクス細菌である Bacteroides fragilis による治療により、4-EPS の量が減少した [82]。 プロバイオティクスまたはプレバイオティクス、ならびに糞便微生物叢移植(FMT)は、消化管異常および自閉スペクトラム症重症度を改善するために自閉スペクトラム症臨床試験で研究されています[83]。 19 週間の実験期間中、13 人の自閉スペクトラム症の子供にプロバイオティクス( LactobacillusとBifidobacteriumを含むビボソーム) を与えたところ、消化管の問題が大幅に改善されました [84]。 |
| 非盲検臨床実験では、健康な対照から自閉スペクトラム症と診断された子供への糞便微生物叢移植が行われ、自閉スペクトラム症の人々の胃腸関連症状の改善が見られます. この研究は、7~8週間の毎日の維持投与の後、自閉スペクトラム症の子供の80%が胃腸症状と自閉スペクトラム症の重症度を軽減したことを示した[85]。 このコホートの 2 年間の追跡調査では、治療を受けた患者が胃腸および自閉スペクトラム症の症状を改善したことが明らかになりました [86]。 プロバイオティクスおよびプレバイオティクス療法と比較して、糞便微生物叢移植療法は長期的な利益を示しました。 |
| 6. 神経変性疾患における腸内細菌叢 |
| 腸内細菌叢異常は、腸内細菌叢-脳軸 を通じて脳の免疫恒常性に影響を与え、パーキンソン病 (PD)、アルツハイマー病 (AD)、多発性硬化症、筋萎縮性側索硬化症などの神経変性疾患の病因に関与する可能性がある [87]。 |
| 6.1 アルツハイマー病 |
| 認知症は、記憶、行動、推論、日常活動を行う能力、判断力、および言語が低下する状態です。 アルツハイマー病やその他の認知症は、世界中で 5 番目に多い死亡原因であると主張されています [88]。 最も重要な危険因子は年齢であり、アルツハイマー型認知症患者の大多数は 65 歳以上です [89]。 細胞外アミロイド (A)、老人斑 (SP)、および細胞内神経原線維変化 (NFT) は、アルツハイマー病の重要な特徴です。 活性酸素種 (ROS) の産生の増加は、神経炎症と細胞死を引き起こします。 さらに、血管の異常とミトコンドリアの損傷は、アルツハイマー病の病因に関与しています [90,91]。 |
| 6.1.1 腸内毒素症とアルツハイマー病 |
| アルツハイマー病の発症に関連する代謝経路に影響を与えるシグナル伝達タンパク質の生成は、腸内微生物叢の変化の影響を受けます。 老化プロセスは、腸内細菌叢組成、すなわち抗炎症性細菌よりも豊富な炎症誘発性細菌を変更することにより、腸管透過性と血液脳関門機能を損なう局所的な全身性炎症を引き起こします(図5) [92]。 |
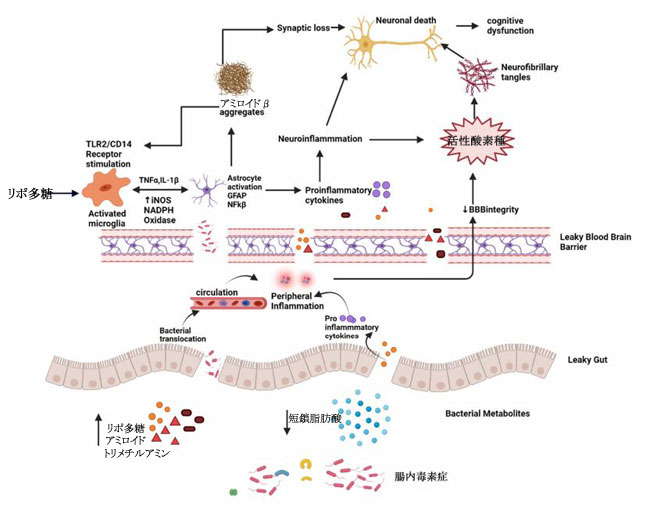 |
| 図5 腸内毒素症とアルツハイマー病 |
| 細菌のアミロイド、リポ多糖、トリメチルアミンを産生し、短鎖脂肪酸などの有益な細菌代謝産物を減少させる炎症誘発性微生物によって腸の平衡が乱されると、腸の透過性が損なわれます。 腸および血液脳関門の機能障害は、末梢および中枢神経系への微生物の侵入を増加させ、炎症誘発性サイトカインの産生を増加させ、末梢および中枢の炎症を引き起こします。 この神経炎症は、神経原線維変化の形成につながる活性酸素種を介して、直接神経細胞死につながります。 リポ多糖は、活性化ミクログリア上の TLR2/TLR4 CD14 受容体にも作用し、TNF-α、IL-1β、iNOS、NADPH オキシダーゼを増加させ、アストロ サイトの活性化と NF-kB 活性を増加させ、アミロイドβ 凝集をさらに促進します。 アミロイドβ は TLR4 受容体に対するアゴニストとしても作用し、アミロイド凝集の悪循環を促進し、最終的には アルツハイマー病 の神経細胞死を促進します。 |
| 6.1.2 アルツハイマー病に関与する代謝物 リポ多糖 (LPS) |
| リポ多糖 (LPS) は、グラム陰性菌の細胞壁の主要な成分である脂質糖化合物です [88] (正常な腸内微生物叢では 50 ~ 70%)。 リポ多糖は、神経変性疾患における神経炎症を調査するための貴重なツールです [93]。 腸上皮細胞間の緊密な結合により、リポ多糖が健康な人の血流に入るのを防ぎました. 緊密な接続が弱まると、リポ多糖が血流に入り、炎症を引き起こします。 その結果、血中リポ多糖レベルは、炎症だけでなくリーキーガットも示します。 多くの in vivo および in vitro 調査により、リポ多糖は、いくつかの炎症性メディエーターの発現を変化させる多くの細胞内分子を活性化し、神経変性に寄与または開始することが明らかになりました。 リポ多糖は、白血球およびミクログリアの TLR4-CD14/TLR2 受容体を活性化し、NF-kB を介したサイトカイン サージを引き起こし、アミロイドβレベルを上昇させ、オリゴデンドロ サイトを損傷し、アルツハイマー病の脳にミエリン損傷を引き起こします。 アミロイドβ1–42 は TLR4 アゴニストでもあるため、アルツハイマー病の持続的な進行の原因となる悪循環を引き起こす可能性があります [94]。 血液-脳関門も血清 リポ多糖によって破壊され、これが脳に入り、ミクログリア、星状細胞、および多数のアミロイド形成および炎症経路を再活性化する可能性があります。 炎症性サイトカインと NF-kB のレベルが上昇すると、アミロイド前駆体タンパク質 (APP) と アミロイドβタンパク質の切断と蓄積が促進され、神経細胞の喪失とアルツハイマー病の発症につながる [88]。 Zhao et al (2019) は、リポ多糖投与が C57BL/6J マウスの海馬で、病気の行動と認知障害、ミクログリアの活性化と神経細胞死を引き起こすことを示しました。 リポ多糖処理により、IL-4 および IL-10 のレベルが低下し、TNF、IL-1、PGE2、および一酸化窒素 (NO) のレベルが上昇しました。 ウェスタンブロット分析によると、NF-kB シグナル伝達経路は リポ多糖グループで活性化されていました。 さらに、TLR-4 特異的阻害ペプチドである VIPER は、リポ多糖誘発性神経炎症および認知障害を軽減した [95]。 Thingoreら(2020)によると、リポ多糖注射は神経炎症を上昇させ、記憶保持の低下を引き起こし、認知機能の低下を悪化させ、スーパー・オキサイド・ディスムターゼ(SOD) を低下させ、脂質過酸化を増加させることによって酸化ストレスを引き起こした [96]。 |
| アミロイド |
| アミロイドは、神経変性疾患患者の細胞機能障害を誘発する可能性がある自己凝集タンパク質です [97]。 アミロイドβ(Aβ)は、神経細胞の成長、シグナル伝達、および細胞内輸送に関与する膜貫通タンパク質であるAPPの切断産物である[98]。 GM 産生アミロイドは、いくつかの in vitro および in vivo 研究で アミロイドβ沈着を交配することが示されている [99]。 Curli は Escherichia coli によって作成され、TasA は |
| カルプロテクチン |
| カルプロテクチンは、S100A8/A9 (TLR4 リガンド) のヘテロ二量体である、好中球と単球によって生成される小さなカルシウム結合タンパク質です。 糞便カルプロテクチンの上昇は、腸の炎症の徴候として作用する可能性があります。 アルツハイマー病患者 22 名の糞便カルプロテクチンの濃度が、Leblhuber らによって芳香族アミノ酸の血清量と比較されました (2015)。 糞便中のカルプロテクチン濃度の上昇は、アルツハイマー病患者の腸バリア機能の障害に関連している[106]。 |
| 6.1.3 漏れやすい腸と漏れやすい脳 |
| 粘液層、腸上皮、固有層 [107] は腸のバリアを形成し、病原菌から体を保護し、有毒な粒子、化学物質、バクテリア、その他の健康を脅かす生物が血流に入るのを防ぎます。 微生物叢の構成は、粘液層の透過性に影響を与える [108]。 多数のムチン分解細菌 Akkermansia muciniphila は、腸のバリア機能と全身性炎症を改善します [109]。 タイトジャンクションの変化は、病原性E. coli , Salmonella, Shigella, Helicobacter pylori, Vibrio, またはClostridiumによって媒介される [110]。 リーキーガットとして知られる腸の透過性の増加は、タイトジャンクションの能力の問題によって引き起こされます。 Bacteroides fragilis 外毒素は、E-カドヘリン (細胞接着分子) を切断することにより、接着結合を破壊します [111]。 腸のホメオスタシスの乱れは、短鎖脂肪酸や H2 などの有益な物質を低下させ、リポ多糖、アミロイド、トリメチルアミン-N-オキシド(TMAO) などの有害な物質を増加させることにより、腸の透過性に悪影響を及ぼします。 腸粘膜バリアを透過性にし、末梢免疫応答を活性化し、末梢および中枢の酸化ストレスレベルを上昇させる [112]。 特殊な脳内皮細胞、星状細胞、周皮細胞で構成される 血液-脳関門 は、高度に選択的な半透過性境界である [113]。 血液-脳関門の整合性は、脳の成長と機能にとって重要です。 最近の研究によると、さまざまな化学物質が血液-脳関門を危険にさらし、タンパク質、ウイルス、さらには細菌などの分子が脳に入り、脳の健康を危険にさらす可能性があります (Welling et al., 2015) (表 1)。 血液-脳関門の構造および機能の崩壊は、アルツハイマー病の病因における初期の重要な段階である可能性がある [114]。 炎症誘発性および細胞傷害性イベントは、血管系への沈着に起因し、アルツハイマー病脳の血液-脳関門透過性の増加に寄与します (Roher et al., 2003, Carrano et al., 2011, Erickson and Banks, 2013)。 TJ は、マトリックス メタロプロテアーゼ (MMP)-2 および MMP-9 の産生を促進しながら、ZO-1、クローディン-5、およびオクルディンの発現を抑制する アミロイドβ1-42 オリゴマーによって破壊されます。 また、RAGE 受容体に結合し、活性酸素種の形成を引き起こします。活性酸素種は、TJ を破壊し、血液-脳関門の完全性を損ないます (Carrano et al., 2012)。 in vitro 研究およびトランスジェニック マウスのタウオパシー モデルによると、タウは血液-脳関門変性も誘発する可能性があります。 したがって、タウとアミロイドβ の両方が血液-脳関門の分解に寄与し、神経変性プロセスとそれに伴う炎症反応を悪化させる可能性がある [114]。 |
 |
 |
| 6.2 パーキンソン病 |
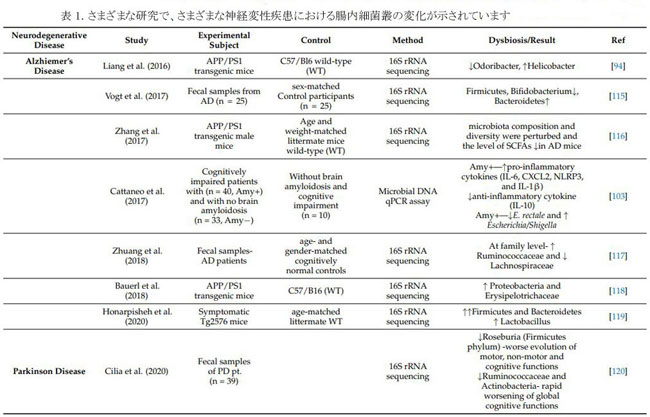
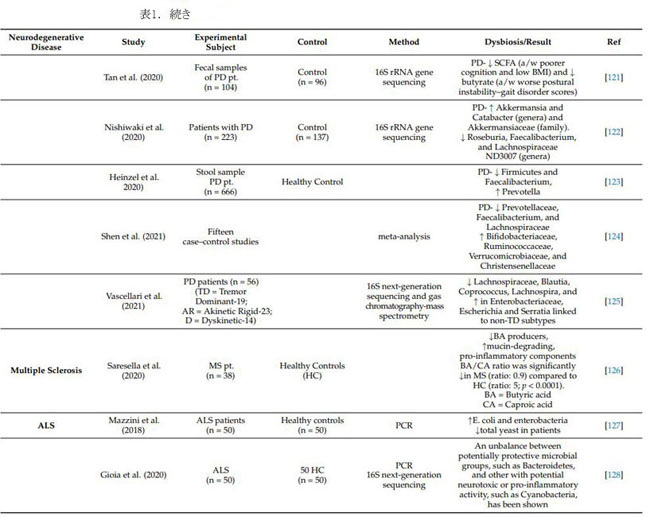
| パーキンソン病 (PD) は世界で 2 番目に多い神経変性疾患であり、黒質 (SN) のドーパミン作動性ニューロンにおけるレビー小体 (LB) として知られる α-シヌクレイン線維の異常な蓄積を特徴とする [129]。 年間 100,000 人あたり 10 ~ 50 人の世界的な発生率と、100,000 人あたり 100 ~ 300 人の有病率があり、世界的な人口の高齢化により、パーキンソン病を持つ人の数は 2030 年までに倍増すると予想されます [130]。 腸の透過性の増加と細菌エンドトキシンの全身暴露は、腸内微生物叢の変化によって引き起こされます。 これは過剰な α-syn 発現を引き起こし、ミスフォールディングをサポートして レビー小体を生成します。 腸のレビー小体は迷走神経を介してCNSに入り、最終的に黒質に移動して破壊し[131]、その結果、震え、こわばり、バランスの問題、自発運動の喪失などのパーキンソン病の臨床的徴候が形成されます(無動)。 便秘は、パーキンソン病で最も一般的な運動前徴候であり、70% 以上の個人が関与し、臨床症状が現れる 10 年以上前に病因が進行します。 その結果、便秘の症状は、前駆期パーキンソン病を特定するための臨床バイオマーカーと見なされます (Berg et al., 2015) [132]。 パーキンソン病患者では、多数の腸内微生物叢の代謝産物が大幅に減少し、便秘につながる可能性があります. 腸の感染症が存在する場合、パーキンソン病に対するより高い脆弱性が報告されており、パーキンソン病のような症状を引き起こす可能性があります。 マウスモデルでは、パーキンソン病由来の腸内微生物叢が α-シヌクレインを介した運動障害と脳疾患を悪化させる可能性があるのに対し、無菌マウスはより軽度の α-シヌクレイン病理を示した (Sampson et al., 2016) [133]。 パーキンソン病のマイクロバイオーム関連の変化については、表 1 で説明します。 |
| 6.3 多発性硬化症(MS) |
| 多発性硬化症は慢性自己免疫疾患であり、免疫細胞がミエリン鞘を標的とし、脱髄と軸索喪失を引き起こし、ミエリンが電気インパルスが神経細胞を流れることを可能にするため、麻痺につながります [134]。 自己免疫疾患の発症に関与する複数のリスク変数にもかかわらず、腸内微生物叢は多発性硬化症の最も重要な環境リスク要因であると考えられている [135]。 多発性硬化症患者は、健康な対照と比較して、Faecalibacterium, Eubacterium rectale, Corynebacterium, および Fusobacteriaの数が少なく、Escherichia, Shigella, Clostridium, および Firmicutes の割合が高かった[136,137]。 ヒトの多発性硬化症の特徴と一致する最も広く使用されている動物モデルは、実験的自己免疫性脳脊髄炎(EAE) です。実験的自己免疫性脳脊髄炎は 無菌マウスでは誘発されず、腸内微生物叢が実験的自己免疫性脳脊髄炎誘発に不可欠であることを示唆しています。 アンピシリン、バンコマイシン、ネオマイシン、硫酸塩、およびメトロニダゾールによる経口療法でも同様の反応が得られ、病気の発症が遅れ、重症度が軽減されました。 炎症誘発性サイトカインのレベルが低く、インターロイキンIL-10およびIL-13のレベルが高い[138]。 Farrokhi らによると、脂質 654 は、健康な人とアルツハイマー病の人の両方と比較して、多発性硬化症患者の血液中にかなり少ない量で発現しています。 (2013) [139]。 プロバイオティクス(Lactobacillus casei, Lactobacillus acidophilus, Lactobacillus reuteni, Bifidobacterium bifidum, およびStreptococcus thermophilusを含むIRT5)は、実験的自己免疫性脳脊髄炎の誘発前に投与されたため、病気の開始が遅くなり、病気の期間がより軽度になりました[140]。多発性硬化症患者における腸内毒素症については、表 1 で詳しく説明しています。 |
| 6.4 筋萎縮性側索硬化症(ALS) |
| 筋萎縮性側索硬化症は、脳と脊髄の神経細胞に影響を与える致命的な神経変性疾患であり、運動神経細胞の早期死をもたらす[141]。 呼吸麻痺のため、筋萎縮性側索硬化症患者の大部分は3〜5年以内に死亡する[142]。 多くの研究により、筋萎縮性側索硬化症の人の腸内細菌叢に異常があることがわかりました。Wu et al (2015) は、筋萎縮性側索硬化症動物モデルを使用して、タイトジャンクション構造が破壊され、腸の透過性が向上することを発見しました。 腸内細菌叢異常は筋萎縮性側索硬化症マウスでも見られ、Butyrivibrio fibrisolvens や E. coli などの酪酸産生菌の数が少ない [143]。 Fang et al (2016) は、 Firmicutes/Bacteroidetes属比の減少、Anaerostipes 属、Oscillibacter 属、および Lachnospiraceae 属 (有益な細菌) の大幅な減少、および筋萎縮性側索硬化症患者の Dorea を代謝するグルコースの有意な上昇を発見した [144]。 筋萎縮性側索硬化症マウス モデルと飲料水に 2% の酪酸を添加した食事を使用して、Zhang らは .対照マウスと比較して、腸内微生物の平衡が回復し、腸の完全性が向上し、寿命が延びたことを発見した [145]。 微生物叢と筋萎縮性側索硬化症に関連するさらなる研究については、表 1 で説明します。 |
| 7. プレバイオティクス、プロバイオティクス、シンバイオティクス、サイコバイオティクスのサプリメントが腸内細菌叢と関連疾患に及ぼす影響 |
| プレバイオティクスは、「宿主細菌によって選択的に使用され、健康上の利点を与える」基質です。 それらはオリゴ糖などの非消化性繊維で構成されており、消化管内のプロバイオティクスの特定の基質として機能し、成長を促進し、機能を改善します [146]。 プロバイオティクスは、「適切な量が供給されると、宿主に健康上の利益をもたらす生きた細菌」です [147]。 シンバイオティクスとは、プロバイオティクスとプレバイオティクスの組み合わせを指します。 プロバイオティクスとプレバイオティクスは、アルツハイマー病、パーキンソン病、うつ病、自閉症スペクトラム障害、その他の神経疾患および精神疾患の予防に有益な影響を与えることが示されています [148]。 「サイコバイオティクス」という用語は、免疫学的、体液性、神経細胞、および代謝経路を介したメンタルヘルスに対するプロバイオティクスおよび/またはプレバイオティクスの治療効果を説明するために発明されました。 [149]。 「酸(乳酸、酢酸)の排泄、栄養素と胃腸受容体部位の競合、免疫調節、および特殊な抗菌剤の作成」は、勝利のプロバイオティクスのメカニズムの1つです[150]. Tsilingiriらは、ポストバイオティクスを「直接的または間接的に、宿主に好影響を与える、微生物の代謝活動によって放出または生成される任意の分子」と定義している[151]。 動物モデルの神経障害に対するさまざまなプレバイオティクス、プロバイオティクス、シンバイオティクス製剤の効果を表 2 に示します。 この豊富な知識にもかかわらず、プロバイオティクスとプレバイオティクスの真の利点はほとんど知られていません。研究を比較すると、いくつかのギャップと不一致があります. プロバイオティクスとプレバイオティクスの有効性と安全性だけでなく、特定の患者グループのフローラの組成を改善するには、ヒトでの研究が必要である [148]。 |
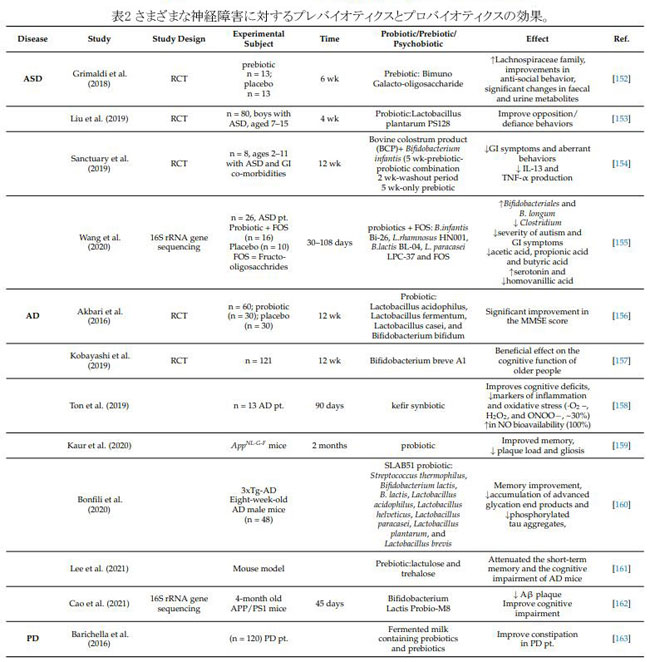 |
| 8. 神経保護剤としてのホウ素 |
| ホウ素 (原子番号 5) は、周期表の 13 族の非金属固体メンバーです。重要なミネラルは、食品と環境の両方に含まれている可能性があります。ホウ素は、いくつかの代謝酵素の活性に加えて、ステロイド ホルモンや、カルシウム、マグネシウム、ビタミン D などのさまざまな微量栄養素の代謝に不可欠である [164]。 さらに、骨の成長と維持、高感度C反応性タンパク質(hs-CRP)や腫瘍壊死因子α(TNF-α)などの炎症性バイオマーカーの減少に重要です。 スーパーオキシドジスムターゼ(SOD)、カタラーゼ、グルタチオンペルオキシダーゼなどの抗酸化酵素のレベルを高め、高齢者の脳の電気的活動、認知能力、短期記憶を強化し、前立腺がん、子宮頸がん、肺がんなどの悪性腫瘍の予防と治療に効果的です [165]。 2003年以来、さまざまな腫瘍に対してin vitroおよびin vivoで作用するプロテアソーム阻害剤であるボルテゾミブ(Velcade)は、悪性癌を治療するための臨床試験で利用されてきました(図6)[166]。 |
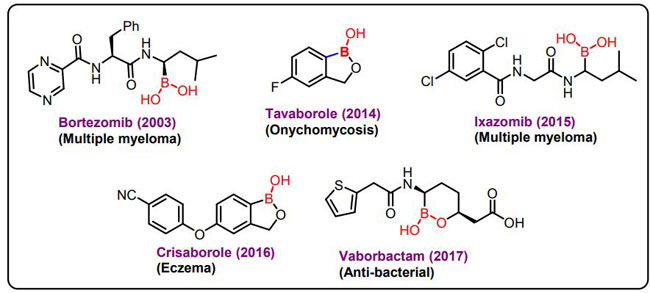 |
| 図 6. ホウ素ベースの市販薬 |
| プロテアソーム阻害剤としてのベルケイドの承認は、医薬品化学におけるボロン酸への関心を引き起こし、さらに 2 つの他の薬物イキサゾミブとバボルバクタムの発見につながります。 2015 年に FDA によって承認された Ixazomib は、多発性骨髄腫の治療に使用されます。 2017 年に FDA によって承認されたバボルバクタムは、β-ラクタマーゼ阻害剤であり、尿路感染症の治療のために抗生物質と組み合わせて使用されてきました。 ベンゾキサボロールを含む薬物であるタバボロールとクリサボロールは、2014 年と 2017 年に FDA によって承認され、それぞれ爪甲真菌症と湿疹の治療に使用されています。 [167,168,169]。 |
| いくつかのホウ素含有物質は、ホスホジエステラーゼ 4 酵素 (PDE4) および炎症関連サイトカインの放出を阻害することも報告されており、どちらも老化およびアルツハイマー病における認知の改善に関連している [170]。 |
| 8.1 食事中のホウ素 |
| 1904 年、Wiley は 500 mg/日 (1 日あたり 77 mg のホウ素) を超えるホウ酸を 50 日間摂取すると、食欲、消化、および健康に異常が生じることを発見し、4000 mg/日 (699 1 日当たりの mg ホウ素) は、通常の男性がそれを超えると怪我をする可能性がある最大値でした。 彼の研究に続いて、ホウ素が健康上の危険性を表すという考えが注目を集めました [171]。 これらの禁止は第二次世界大戦中に緩和されましたが、世界中の多くの国が1920年代半ばまでに食品へのホウ酸塩の添加を禁止し始めました. 戦後、制限は徐々に再課されました。 ホウ素は高等動物や人間にはほとんど栄養価がないという考えは、1980 年代初頭に変わり始めました。 これは、ホウ素欠乏症が低用量のビタミンDを与えられたヒナの肉眼的骨変形を増加させることが研究で発見された年でした[172]。 ホウ素は、ケールやほうれん草などの緑豊かな食品に含まれるミネラルです。 穀物、プルーン、レーズン、柑橘類以外の果物、ナッツにはすべて含まれています. 平均的な人の毎日の食事には、1.5 ~ 3 ミリグラム (mg) のホウ素が含まれています。 りんご、コーヒー、乾燥豆、牛乳、じゃがいもは、人の毎日の食事で最も一般的なホウ素の 5 つの供給源です。 1~3 mg/日のホウ素摂取は、0.25~0.50 mg/日の摂取と比較して、成人の骨と脳の健康を改善することが示されている [173]。 適切な量のホウ素は、消化管吸収を改善する腸の組織構造を作成するのに役立ちます [173]。 |
| ホウ素が欠乏した成体のカエルの雄は、精巣が萎縮し、精子数が減少し、精子の形態異常が見られました。 雌のカエルは卵巣が萎縮しており、卵母細胞の成熟が妨げられていました。 ホウ素欠乏症は、壊死卵の有意な増加、ならびに卵黄出血および外原腸形成によって特徴付けられる異常な原腸陥入の高発生率をもたらした [174]。 |
| 8.2 ホウ素と腸内細菌叢 |
| Evariste らは、実験で六方晶窒化ホウ素とホウ素を含む窒化ホウ素ナノチューブ (BNNT) を使用しました。 オタマジャクシの複数のエンドポイントと、宿主の腸に関連する細菌集団が、暴露後に測定されました。 窒化ホウ素ナノチューブへの暴露はオタマジャクシの成長を促進し、Bacteroidetes門の分類群が恩恵を受ける腸内マイクロバイオームのリモデリングにも関連していました. 調査結果は、研究されたナノ材料からの有害な影響がないことによって証明されるように、窒化ホウ素ナノチューブが生体適合性があるという結論を支持する [175]。 |
| Wang et al [176] の調査結果によると、ホウ素を含む超吸収性樹脂 (SARB) は、トウモロコシのわらの細菌群集の多様性を高めることができます。 Proteobacteriaは、10回の処理で泥炭基質およびトウモロコシわらグループの細菌集団の絶対的な利点の原因であることがわかりました。 一方、実験室で合成されたホウ素を含む超吸収性樹脂(SARB)は、細菌群集の元の構造を変えることができず、泥炭基質とトウモロコシわらの両方で細菌群集にわずかな毒性効果をもたらします。 そして、Proteobacteria と Actinobacteriaに対して増強効果があり、Acidobacteria と Firmicutesに対してある程度の効果を弱める[176]。 これらの例 [175,176] は、腸内細菌叢の生理機能の調節におけるホウ素の役割を明確に示しています。 |
| 8.3 抗炎症剤としてのホウ素 |
| いくつかの研究で、ホウ素は炎症性バイオマーカーのレベルを下げることが示されています。 健康な男性ボランティアを含む最近の人体実験では、11.6 mg のホウ素を補給してから 6 時間後に血漿ホウ素濃度が大幅に増加し、hs-CRP および TNF-α レベルが大幅に低下しました。 10 mg/d のホウ素を 1 週間補給すると、TNF-α が 12.32 から 9.97 pg/mL に 20% 減少しました。 hs-CRP の血漿濃度が 1460 から 795 ng/mL に、IL-6 が 1.55 から 0.87 pg/mL に大幅に減少した (ほぼ 50%) [177]。 ホウ素は、炎症反応に関与するセリンプロテアーゼ酵素の合成と活性を低下させます。 動物実験によると、関節炎を誘発したラットは、経口または腹腔内で供給されたホウ素から恩恵を受けました [178]。 天然に存在する植物ベースのホウ素-炭水化物複合体であるフルクトホウ酸カルシウムは、細胞培養に抗炎症効果があることが実証されており、オープンラベルのパイロット研究では、フルクトホウ酸カルシウムが変形性関節症 (OA) の症状に非常に有益な効果があることが示されています [179] . 次に、Scorei と同僚は、原発性変形性関節症の中年患者を対象に二重盲検試験を実施し、さまざまな用量のフルクトホウ酸カルシウムが全身性炎症と脂質異常症の指標にどのように影響するかを調べました。 彼らは、プラセボ群を除くすべての群で、フルクトホウ酸カルシウムを15日間補給した後、炎症性バイオマーカーであるCRP、フィブリノゲン(FBR)、および赤血球沈降速度(ESR)が減少したことを発見した[180]。 |
| 8.4 神経変性疾患におけるホウ素 |
| ホウ素の栄養摂取は、中枢神経機能に好ましい利益をもたらすことが示されていますが、証拠は骨の場合ほど決定的ではありません。 それにもかかわらず、彼らは、ホウ素が人間にとって健康的な生物活性元素であるという考えを最も受け入れやすい。 ホウ素化合物には強力な免疫調節特性がありますが、アルツハイマー病やパーキンソン病などの神経疾患を治療する可能性を調べた研究はほとんどありません。 |
| すでに説明したように、アルツハイマー病 (AD) は、アミロイド (Aβ) 凝集、過剰リン酸化タウ、神経炎症、および記憶障害によって特徴付けられます。 Maiti らは、アルツハイマー病の in vitro および in vivo モデルを使用して、トランス-2-フェニル-ビニル-ボロン酸-MIDA-エステル (TPVA) (1) とトランス-ベータ-スチリル-ボロン酸の治療効果を比較しました。 (TBSA) (2) (図 7)。 彼らは、TBSA が Aβ42 の凝集を防ぎ、TPVA よりも効率的に細胞の生存を促進することを発見しました。 TBSA の利点は、Aβ42 を発現する C. elegans およびアルツハイマー病の 5xFAD 動物モデルにまで拡大されました また、TBSA は認識および空間記憶の欠損を防ぎ、濃縮および変性細胞の数、GFAP レベル、および Aβ プラークを減少させることがわかった [181]。 |
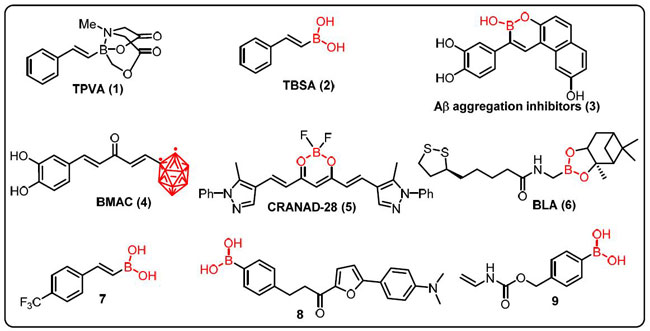 |
| 図 7 神経疾患における活性化合物を含むホウ素 |
| Maiti et al (2020) の調査結果は、ホウ素の食事摂取が人間の脳機能と認知機能を大幅に改善することを発見した Penland の調査結果を裏付けています。 同様に、脳波は、ホウ素欠乏症後のホウ素薬理学的介入が年配の男性と女性の機能を改善したことを示しました。 眠気や精神的敏捷性の低下、精神運動能力の向上 (運動速度や器用さなど)、認知処理の向上 (注意力や短期記憶など) などです。 |
| 別の研究で、Nielsen と Penland は、ラットのホウ素欠乏が、ホウ素補給を与えられたラットと比較した場合、自発活動評価における水平方向の動き、正面からのエントリー、マージン距離、および垂直方向の休憩とジャンプの数、距離、および時間を減少させることを発見した[182 ]。 これらの発見は以前の研究を裏付けており、ホウ素化合物が認識障害と空間記憶の問題の両方に役立つことを示しています。 |
| Lu と同僚 (2012) は、アルツハイマー病の治療において アミロイドβ 凝集阻害剤 (3)、抗酸化剤、および金属キレート剤として作用する一連のホウ素含有化合物を特定した [183]。 クルクミンは現在、アルツハイマー病における神経細胞の損傷を防ぐだけでなく、さまざまな癌を治療する可能性について研究されています. しかし、水溶液中での安定性と溶解度が低いため、その臨床的有用性は限られています。 したがって、Azziらは、クルクミンのβ-ジケトン活性がカルボニル基に置き換わり、2つのフェノール環の1つがオルトカルボランに置き換えられた、まったく新しいクラスのクルクミンのホウ素化モノカルボニル類似体(4)(BMAC)を提案しました。 (正二十面体ホウ素クラスター)。 さらに、アミロイド凝集体の発生を阻害する BMAC (4) の有効性がテストされ、2 つの OH 部分を含む化合物がクルクミンよりも優れていることが発見されました。 2 番目の -OH 基の存在は、β-アミロイド凝集体との化学物質の結合効率を改善することができる [182]。 |
| 研究では、ジフルオロホウ素クルクミン誘導体である CRANAD-28 (5) が、ex vivo と in vivo の両方で画像化するためのアミロイド ベータ プラークの識別に成功する可能性があることも示されています。 CRANAD-28 のイメージングの明るさ (5)、および血液脳関門を通過する能力と低毒性により、アルツハイマー病の研究において潜在的に有用なイメージング ツールとなる [183]。 アルツハイマー病の複雑な性質と病態生理学を考慮して、各単一標的療法の利点を単一分子に統合するための治療技術が開発されています。 Ritacca らは、密度汎関数理論を使用して、ラジカルスカベンジャーおよび金属キレート剤としてのホウ素の抗酸化特性を調査しました。 水性および脂質の設定では、最も実現可能なラジカルスカベンジャーメカニズムである水素移動、ラジカル付加物形成、および単一電子移動が徹底的に調査されました。 金属キレート化の能力は、過剰なアミロイド凝集を触媒することさえできる金属の 1 つである Cu(II) イオンの錯体形成を調べることによって調査された [184]。 |
| Sorout と同僚は、さまざまな曲率の BN ナノ粒子が、アミロイドの凝集とその後の線維化で重要な役割を果たしている β シート型へのペプチドのコンフォメーション遷移を妨げていることを発見しました。 この β シート形成の阻害は、ペプチドのらせん構造 (表面の曲率が最も高い BNNT) を安定化し、ペプチドがランダムなコイルとターン (平面 BNNS) のコンフォメーションに遷移するためのより好ましい経路を利用できるようにすることによるものです。 BN ナノ粒子が アミロイドβ ペプチドからのアミロイド形成を防ぐ効果的なツールとして機能する可能性があることを示している [185]。 新規のホウ素ベースの化合物 (BBC) は、新しいアルツハイマー病治療 (AD) の開発の有望な候補として製造およびテストされています。 Cacciatoreらは、抗酸化部分、すなわちBLAを含む新しいホウ素ベースのハイブリッドを実証しました(6)。 アルツハイマー病患者で酵素活性が増加する酵素であるアセチルコリンエステラーゼ (AChE) (50 M で 22.96%) を阻害しました。 これらの発見は、ホウ素ベースのハイブリッドを使用して、アルツハイマー病の治療のための新しい薬を製造できることを示唆しています [186]。 |
| Jiménez-Aligaga らは、アルツハイマー病の病因に関与する経路を標的とする一連のボロン酸およびボロン酸エステルについて説明しました。 化合物 7 は、神経障害における神経細胞の機能不全に対してより活性であり、その原因であることが判明しました。 さらに、化合物 7 はグルタチオン/グルタチオンジスルフィド (GSH/GSSG) 比を高め、ROS 産生に対する神経保護に重要な GSH レベル (32 nmol/mg タンパク質) が増加し、IC50 は 2.85 μm であった [187]。 |
| Jung とチームは、アルツハイマー病におけるβ-アミロイド斑の検出のためのカルコン由来のボロン酸蛍光プローブを実証しました。 化合物 8 は、合成β-アミロイド凝集体の蛍光応答に対して、蛍光のかなりの増強を示しました。 化合物 8 は、ボロン酸の存在によりβ-アミロイド凝集体 (KD = 0.79 ± 0.05 μm) に効率的に結合し、実験マウスのβ-アミロイドを強く染色しました。 したがって、この研究では、ホウ素ベースの化合物がアルツハイマー病の神経機能を研究するための潜在的なセンサーになる可能性があると説明されています[188]。 | 筋萎縮性側索硬化症は、分泌型 RNase アンギオゲニン (ANG) をコードするヒト遺伝子の変異によって引き起こされる神経疾患です。 合成 BA マスクは、Hoangと同僚によって報告されました。これは、ANG のリボ核酸分解活性を阻害し、神経保護効果を付与します。 通常の生理学的設定では、化合物 9 は不活性ですが、H2O2 の存在下では、PBA のホウ素-炭素結合が酸化的に切断され、活性酸素種を介した損傷を受けた細胞で活性な ANG が選択的に放出されます [189]。 |
| 実験的なパーキンソン病モデルで、Kucukdogru らは、窒化ホウ素ナノ粒子がニューロンを MPP+ (1-メチル-4-フェニルピリジニウム) 誘発アポトーシスから保護することを発見した。 hBN、または六方晶窒化ホウ素ナノ粒子の使用は、MPP+ 治療と比較して、PD モデルでの細胞生存を強化しました。 さらに、hBNを使用した後、酸化剤レベルが減少する一方で、抗酸化能力が増加しました。 最後に、調査結果は、hBN が MPP+ 毒性に対して大きな可能性を秘めており、パーキンソン病の治療における新しい神経保護剤および薬物送達方法として使用できることを示した [190]。 これらの発見は、ホウ素が神経保護効果を発揮し、記憶機能を維持するために重要であるという理論を裏付けています. |
| 8.5 その他の神経保護剤 |
| ホウ素とは別に、他の神経保護剤が文献で入手可能です。いくつかはここで簡単に説明されています。 クルクミンは、抗菌、抗酸化、抗炎症、抗腫瘍ポリフェノールです。 クルクミンは、有益な細菌株を促進し、腸のバリア機能を改善し、炎症性メディエーターの発生を減らします. クルクミン療法は、アルツハイマー病のモデルである APP/PS1 マウスの空間学習と記憶を改善し、その神経保護特性を示唆している [191,192]。 グルタミン酸とその受容体は、学習と記憶を支えるメカニズムであるシナプス可塑性において重要である [193]。 したがって、それらの正常なシグナル伝達の混乱は、アルツハイマー病、パーキンソン病、統合失調症などのさまざまな神経病理学的障害で重要な役割を果たしており、それらを有望な治療標的にしている [194]。 Corynebacterium glutamicum、Brevibacterium lactofermentum、Brevibacterium avium などのグルタミン酸ラセマーゼを持つ腸内細菌は、l-グルタミン酸を d-グルタミン酸に変換し、グルタミン酸 NMDAR に影響を与え、おそらくアルツハイマー病やパーキンソン病患者の認知を改善します。 その結果、腸内微生物叢とグルタミン酸を使用して革新的な認知症治療を開発できる可能性がある [195]。 |
| 最近の研究では、スタチン薬が腸内細菌叢の組成を変化させることが示されています。 HFD を与えられた C57BL/6 マウスでは、ロスバスタチンは腸内微生物叢を変化させ、Lachnospiraceae科の数だけでなく、Rikenella属とCoprococcus 属の数を大幅に増加させた [196]。 アトルバスタチンは、ミクログリアを介した神経炎症を軽減し、オクルディンとムコプロテイン 2 のタンパク質レベルを増加させることで腸のバリア機能を促進し、MCP-1、TNF-α を減少させ、IL-10 を増加させることで腸の免疫機能を調節します。 さらに、アトルバスタチンは、FirmicutesとLactobacillusを上昇させ、Bacteroidetesを減少させることにより、微生物組成を変化させます。 また、リーキーガットのバイオマーカーであるリポ多糖結合タンパク質などの循環エンドトキシンの量も減少させる[197]。 |
| 9. まとめと今後の展望 |
| 腸内細菌叢は、腸と脳の軸の重要な構成要素としての地位を確立しており、健康と病気の両方の基礎となっています。 腸-脳軸は研究分野の最前線であり、いくつかの研究では、腸内微生物叢の組成の変化が自閉スペクトラム症、パーキンソン病、アルツハイマー病、多発性硬化症、筋萎縮性側索硬化症などの神経疾患の病因に重要な役割を果たすことがわかっています。 腸内毒素症は、炎症性サイトカインと細菌代謝産物を増加させることによって神経炎症を促進する可能性があり、腸と血液-脳関門の透過性を変化させる可能性があります。 プレバイオティクスやプロバイオティクスなどの治療物質を使用して腸内細菌叢を変化させることで、さまざまな神経疾患の治療に有望な技術が開かれる可能性があります。 プロバイオティクス、プレバイオティクス、および糞便微生物叢移植の使用は、腸内微生物叢研究のブレークスルーから生じて、腸内細菌叢関連の病気の状態を回復させ、さまざまな症候性疾患管理における代替治療アプローチとしてかなりの可能性を提供します。 重要な微生物叢のメンバーの正確な識別、プロバイオティクスで使用される微生物株の複雑な選択、または理想的な共生生物を選択的に列挙するために提供されるさまざまな形態のプレバイオティクスはすべて、腸内細菌叢ベースの治療を将来の臨床診療に適用する際のハードルに追加されています。 腸内細菌叢に基づく疾患診断、予後モニタリング、予防、および治療の可能性のあるアプリケーションは、現在の疾患管理および治療方法に革命をもたらす可能性があり、楽しみにしています。 注目を集めている薬物研究の 1 つの領域は、ホウ素化合物の使用であり、神経炎症および認知障害を軽減するための将来の治療法として推奨されている [198,199]。 さらに、ホウ素ベースの食事とホウ素化学物質は、腸内細菌叢を改善するために重要な役割を果たし、今後数年間で研究者に新しい窓を開くでしょう。 この急速に拡大する研究分野では、私たちは表面をかじったにすぎない可能性が高く、これまでに私たちを獲得してきた複数の技術は、目の前にある魅力的な課題に取り組む際に目に見えるようになるでしょう。 |
| (略語) |
| GMBA - 腸内微生物叢 - 脳軸。 B - ホウ素; TLR - Toll 様受容体。 MAMPs—複数の微生物関連分子パターン。 NLR - NOD 様受容体。 AMP - 抗菌ペプチド。 PRR-パターン認識受容体; Treg - 制御性 T 細胞。 SIgA - 分泌型 IgA。 SCFA - 短鎖脂肪酸。 GPR - Gタンパク質共役受容体。 HDAC - ヒストン脱アセチル化酵素、IL - インターロイキン。 TNF-α-腫瘍壊死因子-α; NF-kB - 核因子カッパ B サブユニット。 BA - 胆汁酸; 5-HT-5-ヒドロキシトリプタミン; ANS—自律神経系。 HPA-視床下部-下垂体-副腎; ACTH - 副腎皮質刺激ホルモン。 NeuN - 神経核タンパク質。 MBP - ミエリン塩基性タンパク質。 NOS1-一酸化窒素合成酵素 1; IGF-1 - インスリン様成長因子-1。 IGFBP3 - インスリン様成長因子 (IGF) 結合タンパク質-3。 TGF-β - トランスフォーミング増殖因子ベータ。 CD3+ - 分化群 3; BT - Butyrae LPS - リポ多糖。 TMAO - トリメチルアミン N-オキシド; CNS - 中枢神経系。 ROS - 活性酸素種。 iNOS - 誘導型一酸化窒素合成酵素。 NDPH - ニコチンアミドアデニンジヌクレオチドリン酸; Aβ - アミロイドベータ。 PGE2 - プロスタグランジン E2; SOD - スーパーオキシドジスムターゼ。 APP - アミロイド前駆体タンパク質。 TMAO - トリメチルアミン N-オキシド; BBB - 血液脳関門。 ZO-1—オクルディン帯。 RAGE - 高度な糖化最終産物の受容体。 TJ - タイトジャンクション。 GF - 無菌。 RCT—ランダム化比較試験。 BN - 窒化ホウ素。 IC50 - 半最大阻害濃度。 |
| (訳者注:菌名の読み方) Firmicutes :ファーミキューテス Bacteroidetes:バクテロイデス Clostridium:クロストリジウム Blautia:ブラウティア、 Faecalibacterium:フェカリバクテリウム、 Enterococcus:エンテロコッカス、 Lactobacillus:ラクトバシラス Eubacterium:ユーバクテリウム、 Roseburium:ローズブリウム、 Ruminococcus:ルミノコッカス Bacteroidetes:バクテロイデス門 Bacteroides:バクテロイデス属 Prevotella:プレボテラ属 Proteobacteria:プロテオバクテリア、 Actinobacteria:アクチノバクテリア(放線菌)、 Fusobacteria:フソバクテリア、 Spirochaetes:スピロヘータ、 Verrucomicrobia:ウェルコミクロビウム Lactobacilli:ラクトバチルス(乳酸桿菌) Veillonella:ベイロネラ Helicobacter:ヘリコバクター Bacilli:バシラス Streptococcaceae:ストレプトコッカス(連鎖球菌) Corynebacteriaceae:コリネバクテリウム Lachnospiraceae:ラクノスピラ Enterobacter:エンテロバクター Escherichia:エスケリキア(大腸菌) Bifidobacterium:ビフィドバクテリウム(ビフィズス菌) Atopobium:アトポビウム Bifidobacterium longum:ビフィドバクテリウム・ロンガム Bifidobacterium catenulatum: Staphylococcus:スタヒロコッカス(ブドウ球菌) Bacteroides fragilis:バクテロイデスフラジリス E. coli :エスケリキア コリー(大腸菌) Salmonella :サルモネラ Shigella:シゲラ(赤痢菌) Helicobacter pylori :ヘリコバクター・ピロリ Vibrio, ビブリオ Lactobacilli:ラクトバチルス Klebsiella:クレブシエラ Akkermansia muciniphila:アッカーマンシア・ムシニフィラ Clostridium botulinum:クロストリジウム ボツリヌス(ボツリヌス菌) Rikenellaceae: リケネラ Desulfovibrionaceae,: デスルフォビブリオ Clostridium perfringens:クロスロリジウム パーフィリゲンス(ウェルシュ菌) Clostridium saccharolyticum:クロストリジウム・サッカロリティカム Campylobacter jejuni:カンピロバクター・ジェジュニ Bacillus Subtilis,:バチルス ズブチルス(枯草菌) |
参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
この文献は、Molecules. 2022 Jun; 27(11): 3402.に掲載された脳の発達と神経変性疾患におけるマイクロバイオームの役割を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |