ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(発酵乳、腸内細菌の科学:研究の最前線) |
腸内細菌と筋肉機能: プロバイオティクスは筋肉を強化することができますか? |
Muriel Giron et al., |
|
|
概要 |
| 証拠は、腸内微生物叢の組成と多様性が骨格筋の代謝と機能の決定要因となり得ることを示唆しています。 これは、異化作用(サルコペニアおよび悪液質)または同化作用(運動またはアスリート)の状況に当てはまります。 腸内微生物叢は、肥満やインスリン抵抗性などの代謝調節不全表現型の発生と悪化の原因であることが知られているため、骨格筋量と機能を少なくとも部分的に調節できます。 骨格筋は生理的に腸から離れています。 腸内微生物叢 (微生物代謝産物、腸ペプチド、リポ多糖、およびインターロイキン) との相互作用により腸によって生成されるシグナルは、腸内微生物叢の活動と骨格筋との間のリンクを構成し、全身/組織の炎症およびインスリン感受性の調節を介して筋肉の機能を調節します。 . サルコペニアや悪液質を制限したり、げっ歯類の健康状態を促進したりできるプロバイオティクスは、主に乳酸菌とビフィズス菌です。 ヒトでは、同じ細菌がテストされていますが、研究の希少性、個体群のばらつき、および正確な測定の難しさがあります。 再現性の高い筋肉量と機能により、筋肉量と機能を最適化できる特定の株を強調することはできませんでした。 個別化された栄養を設計するためには、より明確な集団に関するさらなる研究が必要です。 高齢者の場合、サプリメントを摂取する前に、フレイルの程度、栄養状態、またはサルコペニアの程度に応じてプロバイオティクスの有効性をテストすることが不可欠です。 運動については、レクリエーションおよび/またはエリートアスリート、レジスタンス、および/または持久力運動で効率的になることができるプロバイオティクスの選択も、さらに注意を払う必要があります。 最終的には、筋肉の同化作用を刺激することが知られているプレバイオティクスやその他の「伝統的な」サプリメント(例:タンパク質)、すべての年齢または患者の健康な個人の筋肉機能を維持するための最良の方法である可能性があります。 |
| 1.序章 |
| 2.微生物叢組成の変化と筋肉量と機能の変化: 微生物叢、腸、筋肉の関係は? |
| 2.1サルコペニア |
| 2.2低栄養 |
| 2.3悪液質(カヘキシア) |
| 2.4身体活動 |
| 3.微生物叢 - 筋肉軸を説明するために動員できる筋肉機能の通常のメカニズム |
| 3.1インスリン感受性 — 短鎖脂肪酸の役割 |
| 3.2炎症 |
| 4.プロバイオティクスは異化状態と同化状態で筋肉量と機能をどのように調節できるのでしょうか? |
| 4.1異化状態におけるプロバイオティクス |
| 4.1.1げっ歯類における筋肉の異化状態 |
| 4.1.1.1がん |
| 4.1.1.2エージング |
| 4.1.2ヒトの筋肉異化状態 |
| 4.1.2.1がん |
| 4.1.2.2エージング |
| 4.2運動中またはアスリートのプロバイオティクス |
| 4.2.1げっ歯類モデル |
| 4.2.2ヒト |
| 5.結論—展望 |
1.序章 |
|
骨格筋は、人間の総質量の 40% を占め、可動性 (移動と姿勢) や体温調節、糖/アミノ酸代謝の維持など、さまざまな基本機能に関与しています。これらの組織は、体の全タンパク質の 50 ~ 75% を占めており、異化状態で要求される可能性のあるタンパク質貯蔵庫と見なすこともできます。 筋肉タンパク質の状態は、タンパク質の合成と分解の間の厳密な調節の結果であり、絶食状態での異化作用を相殺する食事摂取後のタンパク質同化作用につながります。 アナボリズム/カタボリズムの毎日の食事に関連するサイクルは、年齢、生理学的状態、および/またはライフスタイルおよび/またはフィットネスによって長期的に修正される可能性があります。 そして、筋肉量の減少 (サルコペニアおよび悪液質) または増加 (ストレスおよびレジスタンス運動後の回復) につながります。 これらの要因は、数日、数週間、または数か月以内に筋肉量に大きな影響を与えることが知られています。 栄養戦略は、筋肉量を改善したり、損失を制限したりするためにすでに開発されています(1)。 これには、インスリン感受性を高めたり、軽度の炎症や酸化ストレスを制限したりするタンパク質の補給や微量栄養素が含まれます(1)。 単独で、または組み合わせて実施されるこれらの戦略は、筋肉量と機能を刺激することが証明されていますが、その効率は最適ではなく、対象とする集団に大きく依存します(2)。 高齢者の場合、筋肉量の減少 (サルコペニア) は多因子性であることが示されており(3)、食欲減退と身体活動の低下によって増強されます。 一方、運動人口では、筋肉は努力によって強く求められ、繰り返し (微小) 損傷を受け、常に損傷からの回復に必要な最適なアナボリック状態の稜線に沿っています。 |
これらの多様な集団では、現在使用されているものを補完する筋肉同化作用を最適化できる、より具体的または新しい戦略を開発することに関心が高まっています。 いくつかは腸内微生物叢に基づいており、これはいくつかの代謝障害の発症の原因であると疑われています。 実際、微生物叢の変化は、肥満、糖尿病、および心血管疾患の発症と相関し、時には関連していることが示されています。これらの病理はすべて、一般的にインスリン抵抗性状態の確立と軽度の炎症に関連しています。 並行して、筋肉の異化作用は、多くの生理学的状態で、炎症やインスリン抵抗性と頻繁に関連していることが繰り返し実証されています。 |
| 腸内微生物叢組成の変化、生理学的状態の障害、および筋肉の異化状態の間の一致は、微生物叢が直接的または間接的に筋肉量の状態と調節に影響を与える可能性があることを示唆しています。 そのため、特にサルコペニアの高齢者において、「腸 - 筋肉軸」の仮説 (すなわち、腸内微生物叢と宿主腸との相互作用が骨格筋の代謝と機能に及ぼす影響) がいくつかの著者によって提案されています( 4、5、6)。 潜在的なメカニズムの中で、腸内微生物叢は同化刺激に対する骨格筋の感受性の調節に介入する可能性があります( 4 , 7)。 これは、最近の研究プロジェクトが、筋肉量と機能を目標とするプロバイオティクスを含む栄養戦略を開発した理由を説明しています。 プロバイオティクスは、十分な量を摂取すると健康に良い影響を与える生きた微生物です。 長い間、市販されているプロバイオティクスは、主にヒトまたは食物由来の乳酸菌 (LAB) およびビフィズス菌株でしたが、現在の戦略は、腸内に既に存在する細菌の使用にますます焦点を当てています。 プロバイオティクスの作用メカニズムは複雑で、ほとんど理解されていませんが、その目的の 1 つは、変化した腸内微生物叢の組成を調節することです。 |
| 本レビューの目的は、最初に相互作用に関する最近のデータと、微生物叢と筋肉の間の関連性を説明するために実証または仮説化された基礎となるメカニズムを報告することです。 次に、異化状態または同化状態にある集団の筋肉量の減少を制限したり、筋肉量の増加を促進したりするためにこれまでに開発されたプロバイオティクス戦略に焦点を当てます。 |
| 2.微生物叢組成の変化と筋肉量と機能の変化: 微生物叢、腸、筋肉の関係は? |
| 2.1.サルコペニア |
| サルコペニアは、骨格筋量と筋力の進行性かつ全身性の喪失と定義されており(8)、最近、世界保健機関によって疾患として分類されました。 この疾患は依然として老齢に関連することが多く、人生の早い段階でも発生する可能性があります (悪液質を伴う炎症性疾患および臓器不全)。 その診断には、筋力、筋肉の量/質、および重症度の指標としての身体能力の 3 つのパラメーターの低レベルの存在が必要です(8)。 高齢者の数と寿命が延びるにつれて、サルコペニア高齢者の割合は、特定するのは困難ですが、60 歳以上の人々の 20% に達する可能性があります。 さらに、サルコペニアは、それが伴う多くの結果(例えば、転倒、入院、および依存のリスク)のために主要な公衆衛生問題であると考えられている虚弱症候群(フレイル)の病態生理学における中心的要因として認識されています(9)。 虚弱症候群は多次元的であり、身体的側面 (すなわち、Fried らによって説明された虚弱の表現型) および認知的および社会的側面を含む身体のいくつかのシステムおよび機能の累積的な低下を特徴とします。 その結果、虚弱表現型の一部には、サルコペニアのいくつかの特徴 (大幅な体重減少、握力の低下、および歩行速度) が含まれます(9)。 |
| 過去 10 年間、いくつかのグループが、サルコペニア、フレイル、またはより頻繁に高齢者の集団における微生物叢の組成を研究してきました (図 1)(1)。 腸内微生物叢の構成が年齢とともに進化し、重要な個人間変動を伴う微生物叢の多様性の減少によって特徴付けられることは、現在十分に確立されています(10,11)。 高齢者のコホートにおける腸内微生物叢の組成の分析は、健康状態が重要な役割を果たしている年齢の関数としての微生物叢の組成の変化を示しました(12、13)。 虚弱な高齢者の腸内細菌叢の構成は、健康な同年代の人々と比較してより変化しています。(10, 14,15)。 これに応じて、虚弱スコアが高い人(グローニンゲン虚弱指標またはロックウッド臨床虚弱スケールで決定)は、虚弱スコアが低い人に比べて、lactobacilli, Bacteroides/Prevotella, and Faecalibacterium prausnitziiおよびFaecalibacterium prausnitziiの相対存在量が大幅に減少し、Enterobacteriaceaeが増加しました(14,16)。F. prausnitziiは特に、 F. prausnitzii によって生成される代謝物 (酪酸など) が抗炎症特性を示すため、良好な腸の健康の指標として認識されていることに注意してください。 並行して、コホートを使用した研究では、非サルコペニア対照と比較してサルコペニアは短鎖脂肪酸 (非消化性食品の発酵に由来する酢酸、プロピオン酸、酪酸を含む) 産生細菌 (例えば、Roseburia, Eubacterium ) の減少と関連していることが示されています(13,15)。 |
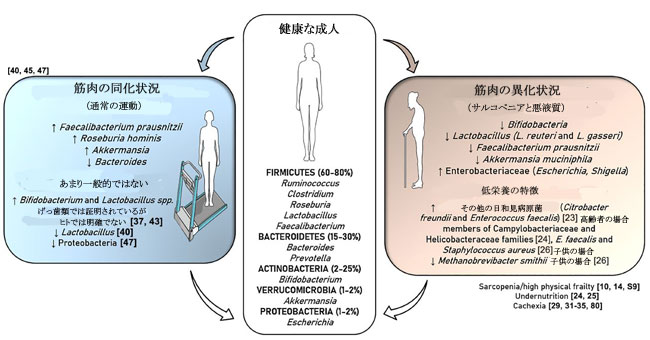 |
| 図1 健康な成人の腸内微生物叢の主要な門と属、および運動とサルコペニア/低栄養と悪液質の影響 健康な腸内微生物叢の主要な門は、Firmicutes, Bacteroidetes, Actinobacteria、そして程度は低いものの、Verrucomicrobia とProteobacteriaです。 運動は、Faecalibacterium, Roseburia,およびAkkermansia属の割合を増加させます。 げっ歯類を対象とした研究では、運動を行うとBifidobacteriumとLactobacillus spp.が増加することが明確に示されています。 フレイル(およびその主要な構成要素であるサルコペニアと低栄養)と悪液質は、これらの有益な属の減少とEnterobacteriaceae科の日和見病原体の増加に関連しています。 低栄養(高齢者と子供の両方)は、さまざまな病原体の大幅な増加に関連するという特殊性を持っています。 |
| 無菌マウスなどの実験計画/モデルにより、筋肉量と機能の制御に対する微生物叢の潜在的な因果関係に関する貴重な証拠が追加されました。 無菌および抗生物質で処理されたマウスは、従来の対応するマウスと比較して、筋肉量と筋力が減少しています(17, 18, 19)。 興味深いことに、これらの筋肉量と機能の変化は、微生物叢の移植または自然な再播種によって回復することができます(17,18). 微生物叢の因果的役割を客観化することは、投薬、食欲減退、栄養状態または関連する慢性病状などの微生物叢の組成を変更することが知られている多くの交絡因子のために高齢者では複雑です(20)。 しかし、増加する証拠は、腸内微生物叢が他の要因の中で、高齢者の筋肉量の減少と機能の発生における因果的役割を果たしている可能性があることを示唆していま(4,5,7)。 (HF)または(LF)の高齢ボランティアから無菌動物に糞便微生物移植(FMT)を行ったところ、は不良なグループと比較してより大きな筋力を示すことが示された(21)。 身体機能が良好なマウスの微生物叢は、不良なマウスと比較して Prevotella と Barnesiella に富んでいました。 これらの細菌属には、短鎖脂肪酸(酢酸、プロピオン酸、酪酸など)を生成する酵素をコードする遺伝子が含まれています。これは、以前のサルコペニア集団で説明されたものとは反対です。 これらの有望な結果は、多様な高齢者集団で確認する必要があります。 |
| 2.2.低栄養 |
| 前述の脆弱性の問題と部分的に関連して、低栄養 (すなわち、必要量に対して食事からの栄養素の摂取が不十分) とその微生物叢への影響が、年齢の点で極端な 2 つの集団 (子供と高齢者) で調査されています。 先進国では、高齢者が低栄養に苦しむ可能性があり、特に虚弱で入院している人々がそうです。 加齢による食欲不振(すなわち、老年期の食欲および/または食物摂取量の減少)は、加齢に伴う筋肉量の減少を悪化させ、高齢者を依存段階に突入させる可能性があります(22)。 |
| ある研究では、介護施設の居住者 (65 歳以上) と栄養失調人口のマイクロバイオームに類似性があることが示されました(23)。 これら 2 つの集団は両方とも、最近炎症性関節疾患に関連する種である Ruminococcus gnavus S12 の増加と、酪酸生産菌 (Roseburia intestinalis) の減少を示しました (図 1)(1)。 栄養失調または栄養失調の危険にさらされている人々の特殊性は、深刻な感染症を引き起こす可能性がある特定の日和見病原体の大幅な増加です(23)。 同じ国の同じ年齢の健康な子供と比較して、重度の栄養失調の子供では、微生物叢はProteobacteria門(EnterobacteriaceaeまたはCampylobacteriaceaeおよびHelicobacteraceae科)に富んでいます(24, 25, 26)。 |
| このコロニー形成は、小腸の形態の変化 (表面積の減少、透過性の増加、および上皮細胞のアポトーシス) につながる腸の炎症の発生の原因となり、最終的には腸の吸収不良につながります(24)。 |
| Smith et al.は、極度の低栄養状態であるクワシオルコル症候群における腸内微生物叢との因果関係を最初に示唆した(27)。 クワシオルコル症候群の有無にかかわらずマラウイの双生児患者から無菌マウスへの糞便微生物移植を行った。 マラウイの食事(タンパク質と微量栄養素が不足している植物ベースの食事、および対象集団の通常の食事)とクワシオルコル双子からのマイクロバイオームの組み合わせにより、レシピエントマウスの栄養不足の子供の表現型(つまり、体重減少)がもたらされました。 ブラントンらによって同じ結論が得られました( 28) .クワシオルコル微生物叢を移入した動物にすぐに使用できる治療食を再給餌しても、げっ歯類の健康を大幅に改善することはできませんでしたが、これらのマウスの成長は、健康な微生物叢を持つマウスと一緒に飼育すると回復しました(28). 2 つの分類群 (R. gnavus と Clostridium symbiosis) は、タンパク質の酸化を変化させるのではなく、タンパク質合成 (および除脂肪体重の成長) を促進することにより、そのような栄養状態での体重増加と成長の調節における潜在的な重要な決定要因として注目されています(28)。 成長率と筋肉量、および人間への翻訳に対するこれらの特定の細菌の役割については、さらに調査する必要があります。 |
| 2.3.悪液質(カヘキシア) |
| 悪液質集団では、急速な筋肉量の減少と腸内微生物叢組成の変化も発生します。 悪液質は、全身の疲労、体重、骨格筋、脂肪量の減少、および食物摂取量の減少を特徴とする多因子症候群です。この用語は、がんの文脈で頻繁に使用されています(29,30)。 この症候群は、C反応性タンパク質(CRP)などの炎症誘発性マーカーの割合が高いことを特徴とする全身性炎症状態にも関連しています(30)。 微生物叢の変化は癌悪液質で観察され (図 1)(1)、いくつかの仮説のメカニズム (全身性炎症または腸バリア機能障害) は、悪液質の発生における微生物叢の因果的役割を示唆しています(29)。 がん患者(結腸直腸がん、乳がん、肺がん)では、Bifidobacterium, Lactobacillus, およびFaecalibacterium属の存在量の減少が、EnterobacteriaceaeとEnterococcusの増加と同時に観察されます(31 , 32 , 33)。 同様の微生物結腸パターンは、慢性腎臓、心臓、または肝臓疾患を示す他の非癌性悪液質患者にも見られます(29, 34, 35)。 関連する循環炎症性サイトカインの増加は、さまざまなメカニズム (インスリン抵抗性、炎症、および関連する酸化ストレス) によって筋萎縮を誘発する可能性があります。これについては、後続のテキストで詳しく説明します。 |
| 2.4.身体活動 |
| 身体活動と運動は、健康状態の強力な決定要因であることが知られています。 身体活動の欠如が健康状態の悪化や微生物叢の不均衡と相関しているとしても、そのような状況で健康を促進する微生物叢の因果的役割は、まだ詳細に研究されていません。 特にヒトにおける、腸内微生物叢の組成および/または機能に対する運動の影響を調査した研究はほとんどありません。 しかし、栄養状態と身体活動は一般的に密接に絡み合っているため、健康上の結果に対する運動自体の正確な役割は、依然として評価が複雑です(36)。 運動と栄養がより管理されているげっ歯類の研究では、定期的な身体活動は、腸/糞便微生物叢の細菌の多様性と豊富さに関連しています(37, 38)。 |
| 一般に、Firmicutes/Bacteroidetes比の減少が観察され( 37, 38, 39, 40)、基本的に異化作用の状況で変更された微生物は、運動または運動選手集団で逆方向に変更されます (図 1)(1)。 それでも、運動の性質および/または強度は、微生物叢の組成を差別的に変化させる可能性があります(自発的な運動と強制的な運動 (41, 42)。 たとえば、自発的なアクセスホイール運動を受けたげっ歯類では、細菌の多様性と主な門の存在量は大きく変化しませんでしたが、強制的な運動(トレッドミル)は、自由に運動した動物や座りがちなコントロールと比較して、有害で炎症誘発性の細菌の割合の増加につながりました(43)。 ある研究では、強制的なトレッドミル運動は炎症を悪化させ、死亡を引き起こしますが、大腸炎のマウスモデルでは自発的なホイールトレーニングが保護的であることが実証されました(44)。 |
| 前述のように、運動をしている人は、食物繊維の摂取量が多いなど、より「健康的な」食事をしている傾向があります(36)。 これは、F. prausnitzii、Roseburia hominis (酪酸生産菌)、Akkermansia muciniphila (酢酸塩とプロピオン酸生産菌) などの良好な免疫学的および代謝的健康に関連するいくつかの細菌が、身体活動によってより豊富になる傾向がある理由を説明できます(45 , 46 , 47)。 同様に、運動は他の酪酸生産菌(40,45)または乳酸生産菌(37,43)の豊富さと関連していることが示されており、乳酸は腸内の酪酸生産菌によって酪酸に変換されることが知られています。 これは、運動と微生物叢を標的とした栄養の組み合わせをより綿密に調査する必要があることを示唆しています。 |
| この意味で、筋肉量と機能に対する微生物叢の因果的役割を評価するために異化作用の状況ですでに行われたことと同様に、研究では運動したマウスから無菌のげっ歯類まで糞便微生物移植が使用されてきました。 高脂肪給餌マウスでは、通常給餌または運動させた動物からの糞便微生物移植がレシピエントの表現型に与える影響が研究されています( 48)。 食事が運動よりも腸内細菌叢を形作るのに強力であるとしても、運動させた動物からの糞便微生物移植(通常または高脂肪食を与えた)は、高脂肪食を与えられた座りがちな動物と比較して、脂肪量と炎症の減少(肝臓でのTNF-αとIL-1の発現)、耐糖能の増加、および血清低密度リポタンパク質(LDL)の減少につながりました(48)。 運動させたマウスの微生物叢の抗炎症特性は、Allen らによってさらに証明されています。実際、炎症を起こした結腸(大腸炎)にさらされた無菌マウスで6週間、回転輪に自由にアクセスできるマウスからの糞便微生物移植は、座りがちなマウスからの微生物叢を移植された同じ動物と比較して、より高い微生物叢の多様性、大腸炎の制限、および体重の改善を可能にしました(49)。 |
| 3.微生物叢-筋肉軸を説明するために動員できる筋肉機能の通常のメカニズム |
| 多くのレビューは、微生物叢がさまざまな生理学的/病理学的状態で健康状態をどのように調節できるかを要約しています。 これらには、代謝調節(エネルギー消費、脂質生成/脂肪分解、インスリン分泌など)、食欲制御、および認知機能が含まれます(50)。 これらの機能の中には、筋肉に関係するものもあります (炎症、エネルギー/栄養素の代謝、インスリン抵抗性、酸化ストレス)。 最近のレビューでは、微生物叢によって合成された分子が、微生物叢と全身代謝の調節との間のリンクとしてどのように考えられるかについても詳しく説明しています(51)。 筋肉量と機能に関与する主要な微生物叢関連の調節メカニズム、つまりインスリン感受性と炎症に焦点を当てます。両方のメカニズムは、酸化ストレスの存在と筋肉レベルでの栄養素代謝の変化と密接に関連しています。 |
| 3.1.インスリン感受性 — 短鎖脂肪酸の役割 |
| インスリンは、食後の状態で筋肉細胞へのグルコースの侵入と使用を刺激し、タンパク質の分解を阻害することで筋肉量を維持するため、骨格筋のホメオスタシスの調節において重要です。 高齢者が筋肉量と筋力の低下に関連するインスリン抵抗性を発症することは何十年も前から知られています。 脂肪毒性の増加に伴う局所/全身の炎症および酸化環境の導入は、高齢者または肥満集団における筋肉インスリン感受性の低下の最も実証された原因です。 それどころか、適度な身体活動を実践すると、インスリン感受性が向上し、筋肉のグルコース取り込み、グリコーゲン合成、およびタンパク質同化作用が増加します(52)。 このため、運動トレーニングは、インスリン抵抗性とサルコペニアに対する治療戦略と見なすことができます。 微量栄養素/抗炎症/抗酸化物質(ビタミンAとE、ポリフェノールなど)やアミノ酸補給(ロイシンまたはロイシンリッチタンパク質)など、多くの栄養戦略もその分野でテストされています. 新しいパラダイムは、Bäckhed et al および Turnbaugh et al による、組織のインスリン感受性における腸内微生物叢の因果的役割に光を当てた、栄養的に誘導されたインスリン抵抗性のげっ歯類モデルにおけるプリンケプス研究から生じます(53 , 54)。 これらのデータは、健康な患者から末梢インスリン感受性が改善されたメタボリックシンドロームの個人への糞便微生物移植を行ったヒトで確認されています(高インスリン正常血糖クランプ下でのグルコース消失率を使用)(55)。 ただし、インスリン感受性パラメーターに対する糞便微生物移植の有益な効果は、常に観察されたわけではありません(56)。 栄養自体の役割は、依然としてインスリン感受性の主要な要因と考えられており、糞便微生物移植の実施 (ドナー効果) に関する補完的な研究の開発が必要です。 |
| 腸管腔内の微生物叢によって生成され、微生物叢、腸、および筋肉のインスリン感受性の間のリンクを構成する可能性のある多数の代謝産物の中で、短鎖脂肪酸は広く研究されています(57)。 短鎖脂肪酸産生菌は、サルコペニアや悪液質の患者とは対照的に、運動によって増加することが繰り返し示されています。 腸管腔に見られる短鎖脂肪酸のほとんどは、微生物による非消化性食物繊維の嫌気性発酵の最終生成物であり、主に遠位回腸と結腸で産生されます。 酢酸、プロピオン酸、酪酸の結腸内でのモル比は約 3:1:1 で、全短鎖脂肪酸の 95% 以上を占めます。 その後、短鎖脂肪酸は腸細胞に吸収され、酪酸はアセチル CoA として TCA サイクルに入り、結腸細胞のエネルギー需要の 60 ~ 70% を提供します(57)。 その後、残りの 短鎖脂肪酸は門脈で発見され、肝臓に到達します (プロピオン酸の約 80% は肝臓に、酢酸は約 40% が取り込まれます)。プロピオン酸は糖新生経路の基質として使用されます(57,58)。 最終的に、短鎖脂肪酸のごく一部 (主に酢酸塩) が骨格筋に到達します (図 22)。 |
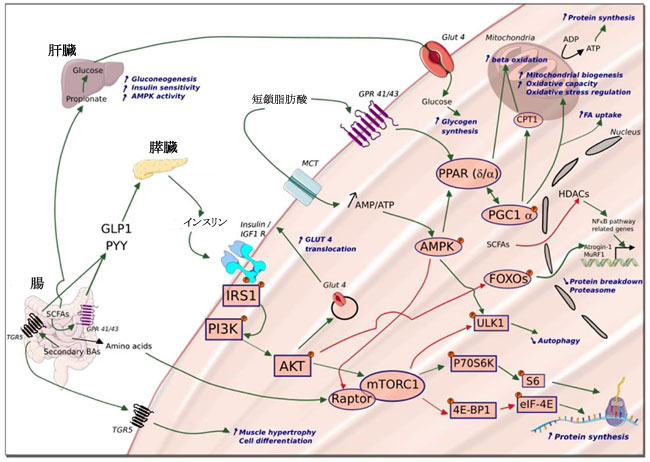 |
| 図 2 骨格筋の健康維持にプロバイオティクスがどのように関与できるか:調節シグナルと代謝産物 |
| プロバイオティクスは、短鎖脂肪酸 (SCFA)、二次胆汁酸 (BA)、および最終的に筋肉機能を調節できるいくつかのアミノ酸などの代謝産物の産生を促進することができます。 短鎖脂肪酸は、回腸および結腸の G タンパク質共役受容体 (GPR) 41/13 受容体との相互作用を介して、腸の L 細胞によるグルカゴン様ペプチド-1 (GLP-1) およびペプチド YY (PYY) の分泌を促進します。 次に、膵臓によるインスリン分泌を刺激し、末梢臓器のインスリン感受性を改善します。 筋肉レベルでは、酢酸塩の取り込みと酸化、および酢酸塩の G タンパク質共役受容体との相互作用により、AMPK のリン酸化と、ペルオキシソーム増殖因子活性化受容体δおよび α の活性化が起こります。 並行して、インスリン、IGF-1、およびアミノ酸は、ホスホイノシチド 3-キナーゼ (PI3K)/プロテインキナーゼ B (AKT)/mTOR 経路を刺激し、[グルコース輸送体 4 型 (GLUT4) を介して] グルコース取り込みを増加させ、タンパク質合成を促進します。 (S6Kの活性化を介して)、フォークヘッドボックスO(FOXO)およびUnc-51様オートファジー活性化キナーゼ(ULK1)因子の阻害を介して筋肉のタンパク質分解を減少させます。 |
| 短鎖脂肪酸は、核因子κB(NF-κB)経路関連遺伝子の発現を含む、核内のさまざまな遺伝子の発現の調節に関与するHDAC(訳者注:ヒストン脱アセチル化酵素)も阻害できます。 二次胆汁酸 はまた、G タンパク質共役胆汁酸受容体 Gpbar1 (TGR5) の活性化を介して GLP-1 分泌を促進し、さらなる調査が必要なメカニズムによって筋栄養において役割を果たします。 4E-BP1:真核翻訳開始因子 4E 結合タンパク質 1。 CPT1:カルニチン パルミトイルトランスフェラーゼ I。 eIF-4E:真核翻訳開始因子 4E。 IGF1-R:インスリン様成長因子受容体。 IRS1:インスリン受容体基質 1。 MCT:モノカルボン酸トランスポーター。 mTORC1:ラパマイシン複合体 1 の機械的標的。 P70S6K:p70S6キナーゼ。 PGC-1α:Pparg コアクチベーター 1 アルファ。 PPARα:ペルオキシソーム増殖因子活性化受容体-α。ラプター:mTOR の調節関連タンパク質。緑の矢印: 活性化。赤い矢印: 阻害。 |
| 短鎖脂肪酸は、グルコース/脂質恒常性、炎症の調節、および腸と他の距離組織との関連に関与しています(57,59)。 無菌および抗生物質で処理されたげっ歯類では、以前は減少していた短鎖脂肪酸補給により、筋肉量および/または筋力が回復/改善されました(17, 18, 19 , 60)。 酢酸塩の補給(食事または皮下注射)は、ラットの骨格筋におけるグルコースの取り込みとグリコーゲン含有量を促進し、ウサギでは脂肪酸の取り込みと酸化の増加を通じて脂質の筋肉内蓄積を減少させることが示されています(61)。 同様に、マウスへの経口酪酸補給は、ミトコンドリア機能と生合成、および骨格筋のタイプ I (酸化) 繊維の数を増加させることにより、酸化ストレスと筋肉量の減少を防ぎます(60 , 62)。 短鎖脂肪酸のこれらの代謝効果は、末梢血に存在する酢酸塩については骨格筋に直接影響する可能性がありますが、酪酸とプロピオン酸は末梢血流にわずかしか到達しないため、おそらくそうではありません。 しかし、これらの効果は、グルカゴン様ペプチド1(GLP-1)分泌の刺激、インスリン分泌の刺激、肝臓でのグルコース貯蔵、および骨格筋でのグルコース取り込みに関与する腸ホルモンを介して間接的である可能性もあります(63)。 筋肉に対する短鎖脂肪酸のその他の間接的な効果には、エピジェネティックなメカニズムによる血流の増加と抗炎症特性が含まれます(64)。 コハク酸は、糖新生の基質であるプロピオン酸のようなものであり、De Vadder らは、腸細胞によってコハク酸から生成されたグルコースが、門脈で検出されると (ポータル信号)、満腹感、エネルギー消費、耐糖能、およびインスリン感受性を増加させる可能性があることを示唆しました(65 , 66)。 |
| 3.2.炎症 |
| 変更された微生物生態系組成は、腸の抗炎症反応と炎症誘発反応の間の不均衡に関連している可能性があり、高齢者集団では「炎症」とも呼ばれる慢性全身性低悪性度炎症につながると仮定されています(67)。 炎症の考えられる原因の 1 つは、加齢に伴う免疫系の規制緩和 (免疫老化) であり、病原性細菌や「腸漏出」による腸内コロニー形成に対抗する能力が低下します。 これは、循環エンドトキシンレベル(リポ多糖など)を高めることによって全身性炎症 (68) を引き起こします。 リポ多糖(LPS) は、リポ多糖結合タンパク質によって血中を輸送され、自然免疫細胞に局在する「パターン認識受容体」[toll-like receptor 4 (TLR4) を含む] によって認識されます。 リポ多糖が TLR4 に結合すると、さまざまな細胞内部分 (MAPK など) が動員され、炎症誘発性経路 NF-κB などの細胞シグナル伝達カスケードがトリガーされます。注目すべきことに、NF-κB 経路は、悪液質、激しい運動トレーニング、またはサルコペニアでさえ産生が増加する分子である、炎症性サイトカインおよび 活性酸素(ROS) によっても活性化される可能性があります。 NF-κB は、筋タンパク質の分解 (ユビキチン-プロテアソーム経路) に関与し、炎症を誘発し、筋線維の再生を阻害することにより、筋萎縮に関与することが示されています(70) (図 33)。 |
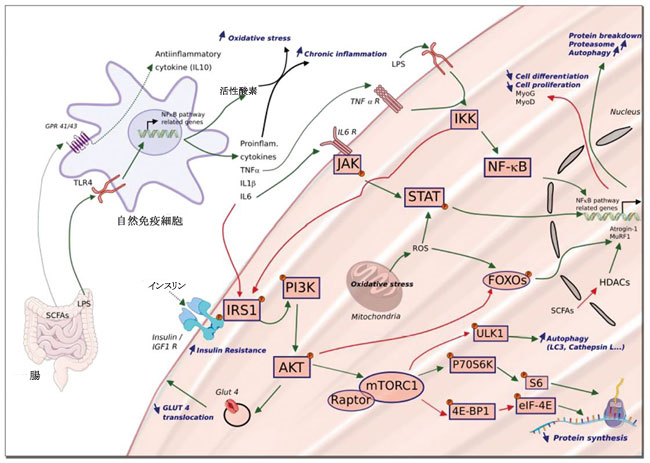 |
| 図 3 炎症の増加に関連した微生物叢と骨格筋の相互作用 |
| 腸内微生物叢の生態系の組成が変化すると、腸からの漏出や末梢血中のリポ多糖類 (LPS) などの細菌エンドトキシンの侵入につながる可能性があります。 これは、腸内腔において有益な代謝産物[例:短鎖脂肪酸 (SCFAs)]の産生の減少にも関連しています。 リポ多糖は、Toll 様受容体 4 (TLR4) 受容体を介して、マクロファージによる炎症性サイトカインおよび活性酸素種 (ROS) の産生を引き起こす可能性があります。 骨格筋では、腫瘍壊死因子α (TNF-α) が核因子κB (NF-κB) 経路関連遺伝子の発現を活性化し、細胞の分化と増殖を減少させます (ミオゲニンとミオDの阻害を介して)。 インターロイキン (IL)-6 および IκB キナーゼ (IKK) は、インスリン受容体基質 1 (IRS1) を阻害することができます。 これは、インスリン抵抗性の誘導に関連して、ラパマイシン複合体 1 (mTORC1) の機械的標的の活性化を制限し、したがってタンパク質合成を制限します。 さらに、プロテインキナーゼ B (AKT) が活性化されなくなったため、フォークヘッドボックス O (FOXO) に対して阻害的役割を発揮できなくなり、これにより、ユビキチン E3 リガーゼである Atrogin-1 と MuRF1 の発現が増加し、筋肉のタンパク質分解が促進されます。 同様に、mTORC1 によって阻害されない Unc-51 様オートファジー活性化キナーゼ (ULK1) は、オートファジーを阻害できません。 これらの調節機構が慢性化すると、タンパク質の分解・合成のバランスが崩れ、筋萎縮を引き起こします。 |
| 4E-BP1:真核翻訳開始因子 4E 結合タンパク質 1。 eIF-4E:真核翻訳開始因子 4E。 GLUT4:グルコース輸送体タイプ 4。 GPR:Gタンパク質共役受容体。 IGF1-R:インスリン様成長因子受容体。 IL-6 R:インターロイキン-6受容体。 JAK:ヤヌスキナーゼ。 P70S6K:p70S6キナーゼ。 PI3K:ホスホイノシチド 3-キナーゼ。 Raptor:mTOR の調節関連タンパク質。 STAT:シグナルトランスデューサーおよび転写のアクティベーター。 TNF-α R:腫瘍壊死因子-α受容体。緑の矢印: アクティベーション。赤い矢印: 阻害。 |
| 老化は、腫瘍壊死因子α(TNF-α)、C反応性タンパク質(CRP)、インターロイキン6(IL-6)などの炎症マーカーの漸進的な増加と関連しています。 TNF-α および IL-6 の血漿レベルは、がん悪液質(71) またはがん悪液質患者のげっ歯類モデルでも上昇しています(72)。 これらのサイトカインの上昇は、がん患者の体重や筋肉の減少、転帰の悪化に関与することが知られています(71)。 同様に、高脂肪食によって誘発される肥満や糖尿病は、炎症マーカーレベルの上昇と関連しており、骨格筋のアナボリズムに悪影響を及ぼします(53)。 対照的に、運動は炎症状態にプラスまたはマイナスの影響を与える可能性があり、さまざまな炎症マーカー間のバランスも考慮に入れる必要があると指摘しています(73)。 それにもかかわらず、定期的な身体運動は、座りがちな人や肥満の人に全体的な抗炎症効果があることを示唆する証拠が増えています(74)。 しかし、運動選手のように運動がより激しくなると、炎症誘発性サイトカインの放出を伴う骨格筋の急性炎症反応につながる可能性があります(73)。 |
| 高循環リポ多糖は、免疫細胞による炎症誘発性サイトカインの発現と産生の増加を誘発し、タンパク質恒常性とミトコンドリア機能を含むいくつかの代謝経路を調節することにより筋萎縮を引き起こすことが知られています。 |
| 急性炎症の動物モデルでは、筋肉タンパク質分解(ユビキチン-プロテアソーム経路)の刺激、タンパク質合成の減少(mTOR制御による翻訳開始による)、だけでなく、細胞アポトーシスの誘導および、またはサテライト幹細胞の分化の阻害も観察されます(75)。 このような同様のメカニズムは、軽度の炎症の状況でも発生する可能性があります。これは、腸の炎症の診断または誘発、筋肉の抑制を伴わずに、α-2 マクログロブリン血漿レベル(ラットの全身性炎症のマーカー)がわずかに増加したラットで示したようにです。 食後のタンパク質合成ですが、成体または炎症を起こしていない老齢動物では通常刺激されます(76)。 興味深いことに、同じラットをイブプロフェンで治療すると、鈍化したタンパク質合成が回復し、炎症状態が食物同化刺激に対するタンパク質合成の応答の欠如の原因であることが示唆されました(76)。 したがって、ヒトでは、タンパク質合成のダウンレギュレーションを伴う高齢者または特定の癌患者で観察される軽度の慢性炎症に関連する場合、IL-6 は筋萎縮とも相関します(4 , 77)。 腸漏れと関連する循環リポ多糖は、激しい運動の長時間の練習中に、ミトコンドリア機能障害、アポトーシス、またはオートファジーを伴う骨格筋内でインスリン抵抗性を生成する可能性がある酸化ストレスの原因となる重要な活性酸素産生を促進することもできます(53)。 この意味で、酸化ストレスや筋肉のインスリン感受性への影響から炎症を分離することは非常に複雑です。 たとえば、IL-6 の影響は、ヒトの 2 型糖尿病のリスク増加に関連する筋肉のインスリン シグナル伝達経路に影響を与える可能性があります(69)。 しかし、IL-6 の役割は逆説的であり、げっ歯類における投与量と投与方法、および他のサイトカインの存在に依存します(77)。 IL-6 は抗炎症効果を持ち、運動中に筋肉自体から分泌されると、マイオカインとして筋肉の成長 (衛星細胞への効果) に重要な役割を果たします。 |
| 4.プロバイオティクスは異化状態と同化状態で筋肉量と機能をどのように調節できるのでしょうか? |
| 微生物叢の活動と多様性が、さまざまな生理学的/病理学的状況における筋肉量と機能の決定要因になる可能性があることを以前に開発したように、腸内微生物叢を標的とすることができる戦略は、筋肉量の減少と機能 (サルコペニアと悪液質) と戦うため、または筋肉のパフォーマンスを最適化するため (アスリート、サルコペニア、病状後の回復) の手段と見なすことができます。 そのような場合、プロバイオティクスは既存の戦略を補完する可能性があり、インスリン抵抗性の発生を制限したり、さまざまな組織の代謝経路を調節したり、炎症や酸化ストレスを制限したりする能力を繰り返し実証しています(78)。 さらに、一部のプロバイオティクスは、非異化作用または同化作用の状況であっても、これらの代謝機能または調節メカニズムの一部を特異的に標的とすることができるようです. 例として、2 種類のプロバイオティクス (Lacticaseibacillus casei LC122 または Bifidobacterium longum BL986) が 10 ヶ月齢 (つまり、老齢ではない成体) のマウスで 12 週間テストされました(79)。 2つのプロバイオティクス株は、同年齢の対照と比較して、これらのラットの骨格筋量と握力を増加させました(79)。 |
| 興味深いことに、 Lactobacillus casei が抗酸化能を示し(筋肉と腸での活性酸素とGPx活性の増加)一方、B. longum は抗炎症特性を示した(筋肉と結腸の TNF-α、IL-6、および IL-1β レベルの低下) ため 2 つの細菌種は異なる細胞経路に作用しているように見えました(79)。 残っている問題は、これらのプロバイオティクスが、挑戦されたとき、つまり、動物モデルと人間の筋肉の喪失(サルコペニアと悪液質)または筋肉の増加(運動後)の状況でも効果的であるかどうかを強調することです。 |
| 4.1.異化状態におけるプロバイオティクス |
| 4.1.1.げっ歯類における筋肉の異化状態 |
| 4.1.1.1がん |
| 乳酸菌は、がんによって誘発された悪液質にさらされたげっ歯類を使用した 2 つの実験でテストされています(80, 81) (表 1)。 最初の実験では、Limosilactobacillus reuteri 100-23 と Lactobacillus gasseri 311476 を 2 週間補給した白血病マウスを使用しました (飲料水、108 CFU/日) (80)。 2 番目の実験は、腸ポリポーシスを示すマウスを対象に、L. reuteri ATCC-PTA-6475 を 3 か月間、以前の研究よりも低用量 (105 CFU) で毎日補給することで構成されていました。 両方の実験は、プロバイオティクスが骨格筋量の減少を制限することを示しました(80, 81)。これは繊維サイズの増加に関連している可能性があります(81)。 ここで、タンパク質分解の阻害が観察された(MuRF1およびAtrogin-1によるユビキチン-プロテアソーム経路は遺伝子発現を減少させた 80 )。 MuRF1 と Atrogin-1 は、腺癌を呈し、Leuconostoc mesenteroides CJ LM119 と Lactiplantibacillus plantarum subsp plantarum CJ LP133 を含む凍結乾燥キムチを添加したマウスの筋肉でもダウンレギュレートされました(82)。 これらすべての実験で、がん移植に伴う炎症はプロバイオティクスの補給によってダウンレギュレートされました。プロバイオティクスは、An et al (2019) のように結腸癌のマウスのアナボリズム (タンパク質合成) を刺激することもでき、そこでは AKT および mTOR リン酸化に対するプラスの影響と、AMPK リン酸化の減少も観察されました。 最後に、Bindels らは、白血病マウスで 2 週間共生生物 (オリゴフルクトースと結合した L. reuteri 100-23) の効果もテストしました(83)。 これにより、腸内のLactobacillus菌数が回復し、筋肉量が維持され、腸内の透過性と免疫に関与する遺伝子が対照動物のレベルまで発現しました。 これらすべてのデータは、一部の細菌 (主に 乳酸菌) が、プレバイオティクスに関連するかどうかにかかわらず、タンパク質分解の減少につながる癌誘発性炎症の抑制とタンパク質合成の潜在的な刺激の両方を介して、骨格筋の同化作用を維持できることを確認しています。 |
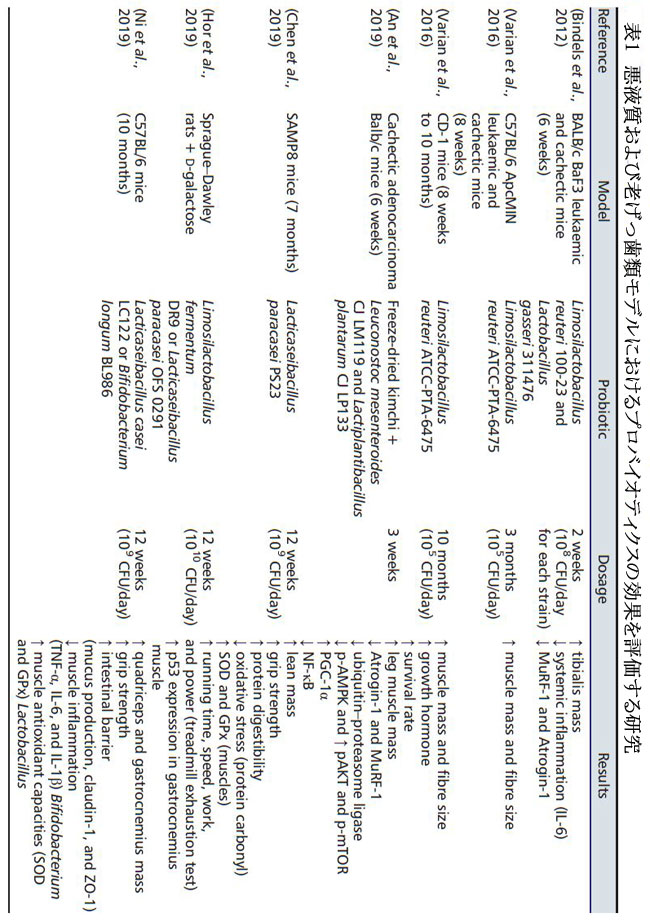 |
| 4.1.1.2.エージング |
| サルコペニアの分野(表1)1)では、げっ歯類の年齢に注意が必要です。 実際、例えばラットでは、加齢に伴う筋肉の喪失は一般に生後 18ヶ月以降に顕著に発生します(84)。 除脂肪量の点で成長していない成体ラット (すなわち 12 ヶ月) は、年配の人間と同等とは見なされず、中年の成体と見なされます。 これらの考慮事項を念頭に置いて、いくつかの研究では、げっ歯類におけるさまざまな期間 (数週間から全生涯まで) のプロバイオティクス補給の影響が調査されています。 Varian らは、L. reuteri ATCC-PTA-6475 株の補給が正常なマウスの筋肉特性と平均寿命に及ぼす影響をテストしました。 プロバイオティクスを長期間摂取した場合、筋肉量、繊維サイズ、および動物の生存率が増加しました。注目すべきことに、この研究で使用されたマウス系統は、他の系統と比較して平均余命が短いか、老化プロセスが加速されていることを特徴とするCD1でした。 実際、すべての CD1 動物は生後 500 ~ 600 日で死亡していました。ただし、この効果は、プロバイオティクス補給の期間がさらに短い他の特異性の低い系統 (C57/BL6 マウス) で発見されています。 |
| 加速老化のモデルは、プロバイオティクス サプリメントの効果を実証するためにも使用されています。 これは、老化促進マウス プローン 8 (SAMP8) および d-ガラクトース誘発老化げっ歯類の場合です。 Lacticaseibacillus paracasei PS23 を 26 週齢の老化促進マウス プローン 8マウス 85 に 12 週間補給すると、除脂肪体重が増加し、筋力 (握力) が 16 週齢の老化促進マウス プローン 8 マウスと同じレベルに回復しました。 これは、筋肉およびミトコンドリア生合成における酸素消費率の改善と関連していました。 並行して、酸化ストレスのマーカーである筋肉タンパク質のカルボニル含有量は低く、筋肉内の抗酸化酵素 SOD および GPx の発現は、老化促進マウス プローン 8動物と対照群で増加しました。 炎症状態は、血漿と筋肉の両方で炎症誘発性/抗炎症性バランスが減少することによっても改善されました(IL-10レベルと発現の増加、炎症誘発性サイトカインの減少)。 Horらは、d-ガラクトース誘発性早期老化Sprague-Dawleyラットのモデルにおいて、異なる乳酸菌菌株(Limosilactobacillus fermentumDR9、L. paracasei OFS 0291、またはLactobacillus helveticusOFS 1515)による12週間の治療によって未処理のラットと比較して、トレッドミル疲労試験後の運動パフォーマンスの向上につながることを示した(86)。 各プロバイオティクスを補給した年配のげっ歯類の筋力と持久力は、d-ガラクトースを投与していない若いラットとほぼ同じレベルで増加しました。 これは、老化マーカー(アポトーシス誘導に関与するp53)の減少、筋肉および骨における同化因子(IGF-1 mRNA)の増加(86)、および炎症状態の改善によって説明されました。 興味深いことに、テストされたすべての菌株は同じ筋肉経路を標的としていませんでした: 酸化状態、AMPKa2、および筋原性因子 (Myf5/MyoD など)。これは、アナボリックな状況では、プロバイオティクス菌株の効果が異なるシグナル伝達または代謝経路および組織を標的とする可能性があることを示唆しています。 |
| まとめると、異化状態にさらされた動物モデルでプロバイオティクスを補給した後に得られたデータは、筋肉量と機能に対する効果がゼロまたは同化であることを示しています。. 一般に、抗炎症効果が観察されますが、これはおそらく、腸からの漏出、炎症の増加、および腸、全身および筋肉レベルでの NF-κB 経路のアップレギュレーションによるものと思われます(87)。 筋肉の標的は多数あり、株間で異なる可能性があります。IGF-1 などの同化因子の刺激、筋原性因子または mTOR 経路の刺激、筋肉のエネルギー状態の調節 (AMPK による)、およびタンパク質分解と酸化状態の阻害です。 |
| 4.1.2.ヒトの筋肉異化状態 |
| この部分の序文として、がん患者または高齢者の筋肉量および機能に対するプロバイオティクスの影響を調査する研究は、存在しない場合でもほとんどないと言わなければなりません. しかし、プロバイオティクスは(単独で、または他の戦略と組み合わせて)、高齢者集団(88)とがん患者の両方で筋肉量と機能を維持するための非常に有望な新しいツールと見なされ始めています(89) |
| 4.1.2.1.がん |
| がん患者において、最も使用されるプロバイオティクス戦略は lactobacilli属であり、より具体的には Lacticaseibacillus rhamnosus CG です。 研究の大部分が結腸直腸癌に関連している場合でも、プロバイオティクスの補給は多くの種類の癌でも研究されています(89)。 腸の健康に対するプロバイオティクスの最も実証された影響は、下痢の発生の制限と、抗がん治療またはがん自体 (例えば結腸直腸の場合) によって損なわれる全体的な腸の健康と微生物組成の改善です(90)。 結腸直腸がんでは、プロバイオティクスが病原菌のコロニー形成を制限し、腸のバリア機能を改善し、ヒトの炎症と酸化ストレスを軽減することが示されています(91)。 筋肉量と機能へのプラスの効果は仮説が立てられただけで、効果的に測定されていません。 |
| 経腸栄養を受けているがん患者にプロバイオティクス補給 (L. plantarum 299v) を使用するプロトコルが最近発表されましたが、結果はまだ入手できていません(92)。 このようなプロトコルは、栄養状態と生活の質、筋肉の健康状態にリンクできる指標に関する貴重な情報を追加する可能性があります。 それでも、がん患者や免疫抑制患者では、たとえプロバイオティクスであっても、これらの患者に細菌を補給することから期待できるリスク/利益比に注意を払う必要があることに留意する必要があります。 |
| 4.1.2.2.エージング |
| 高齢者にプロバイオティクスのサプリメントを使用した研究の大部分は、消化器の健康と免疫調節を対象としており、主にビフィズス菌と乳酸菌の属または異なる菌株の混合物 (VSL#3 mix) に関係しています(93)。 全体として、高齢者集団におけるプロバイオティクスの補給は、場合によっては、病原体の減少を伴う腸内微生物叢の有益な変更につながりましたが、宿主の健康状態への影響は比較的軽微でした(94)。 主な効果は、腸の快適さのわずかな増加と便秘の減少でした(95)が、常にではありません。 いくつかの研究では、自然免疫の増加に対するわずかな正の効果が報告されています (NK 細胞の細胞毒性の増加(96) およびサイトカインプロファイルの改善(97))。 まとめると、高齢者集団で測定された健康パラメータで観察された全体的な小さな影響または観察された結果の不一致は、健康状態、依存 (虚弱概念(9))、および生活習慣 (コミュニティ居住者対居住型ケア)、これらすべての「基礎」パラメーターは、宿主のプロバイオティクス補給に対する反応に影響を与え、最終的に免疫または健康状態を評価するために使用されるパラメーターの妥当性に影響を与えます。 私たちの知る限り、がん患者に関しては、プロバイオティクスを補給した高齢者集団の筋肉量と機能を直接測定した研究はありません。それでも、観察された免疫状態と微生物叢組成のわずかな改善は、炎症の軽減による筋肉への潜在的な有益な効果と一致しています。 さらに、虚弱な高齢者にプロバイオティクス(フラクトオリゴ糖とイヌリン)を補給すると(腸内微生物叢も標的とする)、疲労状態と握力が大幅に改善され、13週間の毎日の補給後に虚弱指数が大幅に減少しました(98)。 これらのデータは、高齢者集団の筋肉機能に対するプロバイオティクスの実際の影響をより適切に評価する必要があることを示唆しています。 |
| 4.2.運動中またはアスリートのプロバイオティクス |
| 4.2.1.げっ歯類モデル |
| プロバイオティクスの補給は、健康な動物でテストされています (運動と相互作用があるかどうかに関係なく) (表 2)。 これらの研究の目的は、若いアスリートの代謝/生理学的反応を模倣できるモデルにおいて、プロバイオティクスが筋肉機能と身体能力を最適化できるかどうかを判断することでした。 |
 |
| 使用されるプロバイオティクスは、一般的に、乳酸菌とビフィズス菌でした( 99, 100, 101, 102)。 すべての研究で、筋肉機能および/または身体能力が改善されました。 L. plantarum TWK10 を 6 週間補給すると、筋肉量 (腓腹筋) が増加し、前肢の握力と水泳持久力がコントロール マウスと比較して改善されました。 Ligilactobacillus salivarius subsp salicinius SA-03 および Bifidobacterium 株 (B. longum OLP-01) を 3 ~ 4 週間処理すると、トレーニング介入なしで若いマウスの筋肉量と水泳持久力も改善されました(100,102)。 最後に、加熱殺菌したBifidobacterium breve B-3 を 4 週間補給すると、マウスの筋肉量と握力が改善され、後者の効果は 2 週間の治療後にすでに現れていました(101)。 他の 2 つのプロバイオティクス: Saccharomyces boulardii 103 と Veillonella atypica 104 もげっ歯類でテストされており、それぞれ VO2max/有酸素パフォーマンスと徹底的なトレッドミルの実行時間が改善されています。 |
| 代謝的に、筋肉レベルでは、パフォーマンスの向上は、酸化型 I 繊維の数の増加、グリコーゲン含有量の増加(100,102)、および疲労関連変数 (血漿乳酸、アンモニア、クレアチンキナーゼ、および血中尿素) の減少によって説明できます( 99, 100, 102)。 運動後に観察された血漿乳酸値の低下と血糖値の上昇(100, 102) は、乳酸からの肝臓の糖新生の改善 (Cori サイクル) と、酸化性筋繊維 (I 型繊維の数の増加) における乳酸異化作用の増加によって説明される可能性があります。 さらに、Leeらは、動物の身体能力と代謝パラメータが用量依存的にプロバイオティクスの補給に反応することを示しており、そのような概念はさらに調査する必要がある( 100,102)。 |
| これらの代謝適応は、加熱殺菌した B. breve を 4 週間補給した後に示されるように、AKT および AMPK リン酸化の刺激と PGC-1α の発現増加によって説明されます(101)。 運動後の VO2max の増加、筋肉の酸化能力およびグリコーゲン含有量の増加(99, 100, 102, 103) が観察されるため、プロバイオティクスを補給した動物ではインスリン感受性の増加が仮説として立てられ、パフォーマンスの改善が説明されます(105)。 プロバイオティクスの他の作用メカニズムは、抗炎症の役割です(99,103)。 しかし、そのような作用機序は、正常な動物または運動した動物で実証されたままです。 実際、筋肉パフォーマンスの分野では、プロバイオティクスが筋肉量および機能/パフォーマンス (つまり、主要な目標) に与える影響に関する研究が存在する場合、作用メカニズムはあまり調査されていません。 |
| 4.2.2.ヒト |
| アスリートや兵士の筋肉量と機能の改善は、既存の戦略に加えて/組み合わせて、プロバイオティック サプリメントを摂取することにより、より多くの文書で証明されています (表 3)。 当初、これらの研究は主にプロバイオティクスの免疫調節および抗炎症特性に焦点を当てており、アスリート集団の感染症や消化器系の不快感を管理または予防でした. (106)。 最近、プロバイオティクスのサプリメントの目的は、身体能力の向上に向けられています. (107)。 |
 |
 |
 |
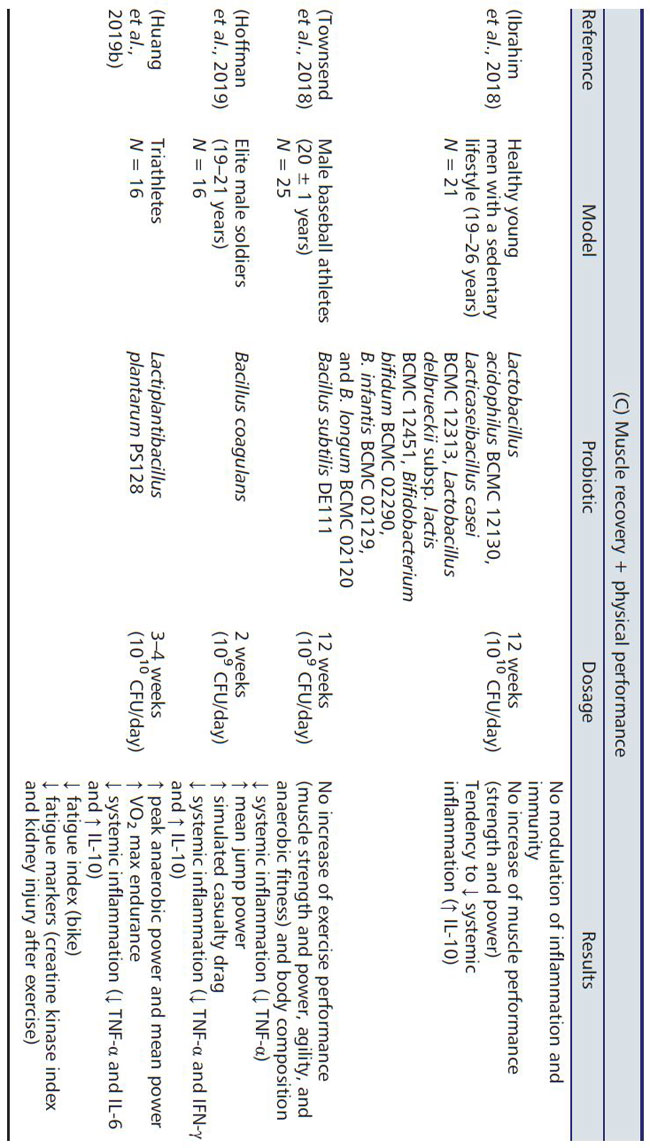 |
| アスリートや兵士のプロバイオティクス補給は、2 つの異なる問題に対処するために研究されました。 (i) 筋肉のパフォーマンス (走る速度と疲労までの時間、および筋力とパワー) の改善、および/または (ii) 激しいトレーニングの実践後の運動による筋肉の損傷の制限。 数週間のプロバイオティクス菌株(Spirulina platensis またはL. plantarum またはBacillus coagulans または複数菌株のプロバイオティクス)の補給は、プラセボ群と比較して疲労回復を改善しました(108,109,110,111)。 並行して、平均ジャンプ力(各ジャンプの強さと速度の評価)を含むいくつかのテストによって決定されたエリート兵士の身体能力、 シミュレートされた死傷者の抗力 (48 kg のバッグを引きずって 50 m の速さで歩くことからなる) は、2 週間の B. coagulans 補給で改善されました(110)。 しかし、プロバイオティクスの補給は、提案された他のエクササイズ (60 秒のプルアップと 100 m のシャトルラン) の後のパフォーマンスを改善しませんでした。 他の著者は、L. fermentum、Bacillus subtilis、および B. longum のサプリメントの効果を調査しましたが、身体能力に対する有意な効果については言及していませんでした(112 , 113 , 114 , 115 , 116 , 117 , 118)。 これは、人間の筋肉のパフォーマンスにおけるプロバイオティクスの補給の明確な有益な効果について結論づけることを許しません。実験結果のこの明らかな不一致は、さまざまな理由で説明できます。 アスリートのタイプ (アマチュア ランナーからエリートまで)、性別、スポーツ (サイクリング、ランニング、トライアスロン、兵士) また、使用したプロバイオティクスの菌株、治療期間 (2 週間から 4 ヶ月)、および研究で選択された参加者の数(108.117)によって研究が大きく異なることは明らかです。 実際、前に述べたように、微生物叢に対する運動の影響は、運動の強度によって大きく異なる可能性があります。 したがって、プロバイオティクスの有益な効果は、対象となる集団と彼らが実践する運動の強度によっても異なる可能性があります( 111 , 113 , 115 , 116 , 117 , 118)。 |
| 以前に議論された同じ研究では、レポートはプロバイオティクスの抗炎症特性に関してより明確です(115,117)。 L. fermentum、L. plantarum、B. coagulans、および B. subtilis の補給は、循環する炎症誘発性サイトカイン (例: TNF-α、IL-6、および IFN-γ) の産生を減少、および/またはIL-10などの抗炎症性サイトカインの産生を増加させます(109, 110, 112, 113)。 最後に、いくつかのプロバイオティック菌株 (Spirulina platensis, B. coagulans, および L. plantarum) は、筋肉疲労マーカー (クレアチンキナーゼや血清乳酸菌など) を減少させることが示されているため、運動後の筋肉の回復を促進する可能性があります( 109,119)。 したがって、これらの集団におけるプロバイオティクスの潜在的な有益な影響をより適切に評価するために、筋肉パフォーマンスおよび疲労のターゲットマーカーと筋肉パフォーマンステストの組み合わせを体系的に検討する必要があります。 |
| 5.結論—展望 |
| 現在までに、プロバイオティクス菌株 (および菌株の混合物) は、同化作用と異化作用の両方の状況でげっ歯類の筋肉量と機能を改善するのに効率的であることが証明されています。 人間に当てはめると、研究の不足、対象となる集団の多様性、および/または筋肉量と機能を正確かつ繰り返し測定することが困難なため、全体像はあまり明確ではありません。 動物モデルでテストされたプロバイオティクスに対するヒトの反応効率の低下を排除することはできません。 これまで、多くの研究で乳酸菌とビフィズス菌が使用されてきましたが、現在、リバース エンジニアリングのアプローチにより、特定のヒト微生物叢 (スポーツ選手など) から細菌株を分離することが可能です。 これらのプロバイオティクス菌株を補給すると、ドナーの表現型の特徴が再現されます。同様に、Faecalibacterium や Roseburia 株などの厳密な嫌気性細菌の使用は、異化作用の状況では過小評価されており、この状況では興味深い代替戦略となる可能性があります。 プロバイオティクスの混合物または糞便微生物移植(「完全な」微生物叢の移動)も、宿主微生物生態系により効率的かつ永続的に定着する可能性があるため、近い将来調査する必要があります。 最後に、筋肉と宿主微生物叢の機能を改善するために、プロバイオティクス株を、微生物叢 (プレバイオティクスやポリフェノールなど) や筋肉 (タンパク質やエネルギーなど) を標的とする他の栄養因子と組み合わせて、効果を最適化することができます。 したがって、腸内細菌叢 - 筋肉軸は、幅広い研究機会を提供します。 |
参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
この文献は、J Cachexia Sarcopenia Muscle. 2022 Jun; 13(3): 1460–1476.に掲載されたGut microbes and muscle function: can probiotics make our muscles stronger?を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |