ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(発酵乳、腸内細菌の科学:研究の最前線) |
腸内細菌叢の調節:結腸直腸癌の管理のためのツール |
Yan Wang and Hui Li |
|
|
概要 |
| 結腸直腸癌(CRC)は、癌による死亡の2番目の原因であり、3番目に頻繁に診断される癌です。 ライフスタイル、遺伝子およびエピジェネティックな変化、および環境要因に加えて、腸内細菌叢も結腸直腸癌の発生に重要な役割を果たします。 腸内細菌叢と宿主との間の共生関係の中断は、病原性細菌が腸内の主要な集団になる細菌集団の不均衡につながる可能性があります。 腸内免疫系を改変し、病原体のコロニー形成を防ぎ、プレバイオティクス、プロバイオティクス、ポストバイオティクス、抗生物質、糞便微生物叢移植(FMT)などの腸内細菌叢の活性と組成を変えるために、さまざまな治療戦略が開発されています。 採用された戦略は有望な結果を示していますが、クリニックへの応用には、潜在的な影響とリスクの評価、およびそれらの長期的な影響の評価が必要です。 この研究は、腸内細菌叢の不均衡とそれらの結腸直腸癌との関係、および化学療法と免疫療法を含む結腸直腸癌療法に対するそれらの影響をレビューするために設定されました。 さらに重要なことに、腸内細菌叢を調節するために使用されてきた戦略、結腸直腸癌の治療に対するそれらの影響、および各戦略の課題を確認しました。 |
|
|
| 目次(クリックして記事にアクセスできます) |
| 背景 |
| 結腸直腸癌の腸内細菌叢 |
| ヘリコバクター ピロリ |
| フソバクテリウム ヌクレアタム |
| 大腸菌 |
| バクテロイデス フラジリス |
| ペプトストレプトコッカス アナエロビウス |
| ポルフィロモナス ジンジバリス |
| エンテロコッカス フェカーリス |
| ストレプトコッカス ガロリチカス |
| 腸内細菌叢と化学療法の意味 |
| 腸内細菌叢と免疫療法の意味 |
| 腸内細菌叢調節の戦略 |
| プロバイオティクス |
| プレバイオティクス |
| ポストバイオティクス |
| 抗生物質 |
| 糞便微生物叢移植 |
| 結論 |
背景 |
|
GLOBOCANの推定によると、結腸直腸癌は、約190万人の新規症例と935,000人の死亡を伴い、2020年の癌による発生率と死亡率のそれぞれ3番目と2番目のランクにあります[1]。 結腸直腸癌は社会経済的発展の兆候と見なすことができます。 大きな変化を遂げている国では、人間開発指数(HDI)の増加に伴い発生率が着実に増加する傾向があります[2、3]。 以前は低リスクで人間開発指数の低い国での増加は、動物性食品の摂取量の増加や座りがちな生活への移行など、食事やライフスタイルの要因の変化を反映している可能性があります。 これは結腸直腸癌のリスクに関連しています[4]。 遺伝的、後成的(訳者注: DNA塩基配列の変化を伴わない細胞分裂後も継承される遺伝子発現あるいは細胞表現型の変化)および環境的要因に加えて、腸内細菌叢と、免疫、代謝、宿主との相互作用などの関連パラメーターが、宿主の健康と病気を決定します[5]。 |
| 消化管(GI)に生息する微生物である腸内細菌には、ほぼ3×1013の細菌だけでなく、真菌、ウイルス、原生生物、古細菌も含まれます[6]。 腸内細菌叢は通常、その宿主と共生関係にありますが、この関係は細菌の組成の変化によって中断される可能性があります。 変調は、病原性細菌が無害な共生細菌に取って代わる細菌集団の不均衡によって発生します。 これらの病原菌は腸壁に侵入し、炎症を引き起こし、発癌性のシグナルや代謝物を誘発し、宿主に損傷を与える可能性があります[7]。 したがって、これらの細菌は結腸腫瘍を促進する可能性があります。しかし、これらの細菌が結腸直腸癌の原因なのか結果なのかは明らかではありません[8]。 腸内細菌叢がどのように癌を促進し、治療の結果に影響を与えるかについての理解が深まるにつれて、腸内細菌叢の恒常性を回復することを目的とした腸内細菌叢の調節は、結腸直腸癌の予防と治療のための潜在的な戦略になりました。 このレビューは、結腸直腸癌の発生における腸内細菌叢の役割と、腸内細菌叢を調節および調節するために適用されるさまざまな戦略を要約することを目的としています。 |
結腸直腸癌の腸内細菌叢 |
腸内細菌叢は、感染に対する防御バリアとしての役割、粘膜免疫系の発達など、さまざまなメカニズムによって腸の恒常性において極めて重要な役割を果たしていますが、 代謝機能に関与することで、人間の腸内細菌叢の不均衡が結腸直腸発癌と関連しているという大規模なメタゲノム研究からの証拠が増えています[9]。 ある研究で、Zhaoらは、腸内毒素症と、結腸直腸癌患者の癌性組織と隣接する非癌性組織との違いを決定するコホート研究をレビューし、結腸直腸癌における微生物叢の不均一性を明らかにしました[10]。 表1は、健康な患者と結腸直腸癌患者の間の系統、属、種の主な違いを説明するためのいくつかの研究をレビューしています。 |
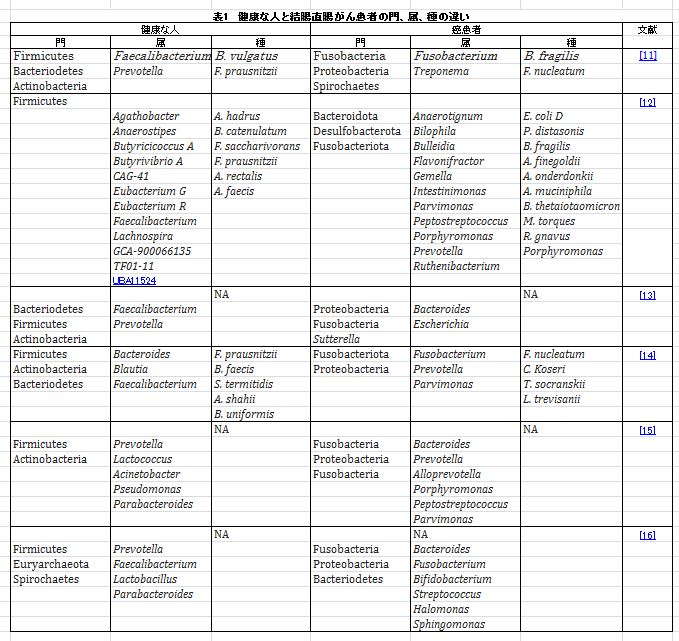 |
| Helicobacter pylori, Fusobacterium nucleatum, Escherichia coli, Bacteroides fragilis, Peptostreptococcus anaerobius, Helicobacter hepaticus, Porphyromonas gingivalis, Enterococcus faecalis, および Streptococcus gallolyticusが結腸直腸癌の発生に関係している[17]。表2は、結腸直腸癌患者のいくつかの細菌の存在量をまとめたものです。 特定の種は、特定のさまざまなメカニズムを通じて結腸直腸癌を発症する可能性があります。 主に、微生物叢の腫瘍促進活性について、病原性細菌病原性因子/毒素、細菌代謝産物、および免疫調節/反応の3つのメカニズムが特定されています[18]。 たとえば、 E. coli またはSalmonellaによってそれぞれ分泌されるコリバクチンまたは腸チフス毒素は、炎症性サイトカインおよび細菌の付着を引き起こします。 また、B. fragilis および F. nucleatumは、抑制性免疫細胞に好ましい微小環境を提供します[19]。 |
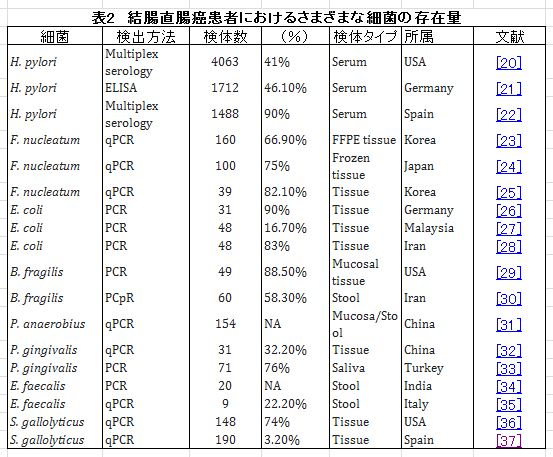 |
| 図1は、腸内細菌叢が関与する結腸直腸癌の主な病原性メカニズムを示しています。 |
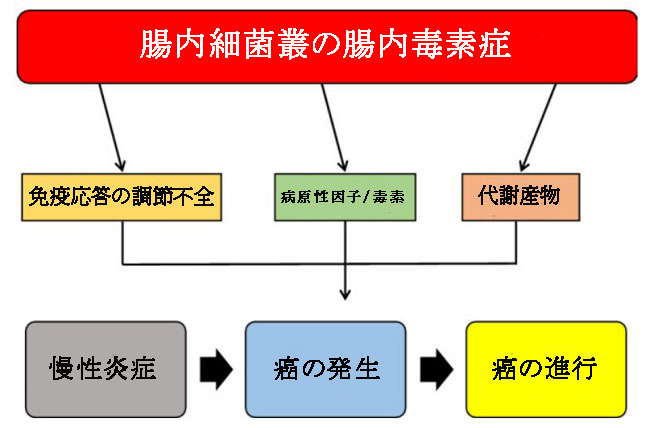 |
| 図1 腸内細菌叢の腸内毒素症とその結腸直腸癌との関係 |
| 腸内細菌叢の腸内毒素症と病原性微生物叢の存在量の増加は、慢性炎症を促進し、その後、免疫応答、病原性因子/毒素、代謝産物の調節不全を含む3つのメカニズムを通じて癌の発症と進行を促進する可能性があります。 |
| ヘリコバクター ピロリ |
| H. pyloriの感染は、結腸直腸癌のリスク増加と関連していることが示されています。 たとえば、Teimoorian et alは、H. pylori感染が、健康な被験者よりも腺腫性ポリープと結腸癌の患者でかなり蔓延していることを発見しました[38]。 17,416件の結腸直腸癌症例と55,811件の対照症例を含む47件の研究を含むメタアナリシス研究ではH. pylori感染と結腸直腸癌リスクの増加との間に正の関連性が明らかになりましたが、この関連性は研究の領域に関連していました[39]。 一部のH. pyloriの病原性因子として、細胞毒素関連遺伝子A(CagA)および空胞化細胞毒素A(VacA)は、炎症経路を促進および活性化する可能性があります[40]。 さらに、H. pyloriのいくつかの菌株による活性酸素種(ROS)および活性窒素種(RNS)の直接的および間接的な産生が結腸腫瘍形成に関与しているという証拠があります[41]。 他の細菌による腸のコロニー形成の変化やガストリン放出の増加など、結腸直腸癌発癌へのH. pyloriの間接的な寄与についての他の仮説もあります[42]。 |
| フソバクテリウム ヌクレアタム |
| メタアナリシス研究で、Gethings-Behnckeらは、結腸直腸癌患者の組織および糞便サンプルにF. nucleatumがより豊富に存在することを発見しました。 彼らはまた、結腸直腸癌患者における高いF. nucleatumの存在量が、全体的な生存率の低下と関連していることを示した[43]。 他の2つのメタアナリシス研究により、腸および糞便のF. nucleatumが結腸直腸癌の貴重な診断マーカーになり得ることが確認されました[44、45]。 F. nucleatumの結腸直腸癌腫瘍形成には、病原性因子、代謝産物、免疫調節、miRNAなどのさまざまなメカニズムが関与しています[46]。 たとえば、細菌のFap2タンパク質は、腫瘍で発現したD-ガラクトース-β(1–3)-N-アセチル-D-ガラクトサミン(Gal-GalNAc)に結合することにより、結腸直腸癌中にF. nucleatumの濃縮を仲介します[47]。 FadAはF. nucleatum別の病原性因子であり、E-カドヘリンに結合することでβ-カテニン経路を活性化し、Wnt7b、NFκB、サイクリンD1、およびMycのレベルを上昇させます[48]。 さらに、F. nucleatumは、接着分子および血管内皮増殖因子受容体1および2(VEGFR1およびVEGFR2)としてのCD31およびCD34の発現を増加させることにより、血管新生を促進します[49]。 血管新生は癌の特徴であり、腫瘍細胞に酸素と栄養素を供給して腫瘍細胞の成長を保証します[50、51]。 |
| 大腸菌 |
| 大腸菌が結腸直腸癌病変および隣接する上皮に頻繁にコロニーを形成しているという証拠が蓄積されています[52、53]。 たとえば、Iyadorai et alは、pks + E. coliが健康な症例よりも結腸直腸癌患者でより頻繁に見られ、pks+E.coli株が腫瘍の発生と発症に重要な役割を果たすことを発見しました[27]。 大腸菌はIL-10欠損マウスに結腸直腸癌を誘発する可能性があることが示され、炎症が腫瘍形成にとって極めて重要であることが示唆されています[54]。 さらに、病原性大腸菌は、サイクル阻害因子、細胞毒性壊死因子(CNF)、コリバクチン、細胞致死性膨張性毒素(CDT)などのシクロモジュリンと呼ばれる毒素を合成できます。これらは遺伝子毒性であるか、細胞周期を妨害します[55、56]。 大腸菌はまた、マクロファージ阻害性サイトカイン1(MIC-1)を誘導することにより、腫瘍細胞の生存を促進します。 トランスフォーミング増殖因子β活性化キナーゼ1(TAK1)の発現が増加し、病原体感染後のRhoA GTPaseが増加し、COX-2の発現が持続します[57、58]。 |
| バクテロイデス フラジリス |
| B. fragilisは、腸管毒素原性B. fragilis(ETBF)と非毒素原性B. fragilis(NTBF)を含む2つのサブタイプに分類されます[59]。 腸管毒素原性B. fragilisは、健康な対照よりも結腸直腸癌の前癌性および癌性病変にかなり豊富であることが報告されており、粘膜B. fragilis毒素(BFT)は結腸直腸癌発症の危険因子およびスクリーニングマーカーと見なすことができます[60]。 粘膜B. fragilis毒素は、Wnt、NFκB、MAPK経路など、結腸の上皮細胞シグナルの伝達に関与する亜鉛依存性メタロプロテアーゼ毒素です。 それは炎症性メディエーターの産生を誘発し、結腸直腸癌の発症を促進します[61]。 また、腸管毒素原性B. fragilisは、STAT3の活性化を介してMinApc±マウスでIL-17依存的に大腸炎と腫瘍の形成を誘発する可能性があります[62]。 これらの経路の活性化は、CXCケモカインを誘導することによって未成熟骨髄細胞の動員をもたらし、炎症誘発性環境の作成につながります[63]。 |
| ペプトストレプトコッカス アナエロビウス |
| 結腸直腸癌患者の粘膜および便サンプルのメタゲノム分析により、P. anaerobius が結腸直腸癌に富む別の腸内細菌であることが発見されました[64、65]。 Tsoi et alは、P. anaerobius に曝露されたマウスは、細菌を投与されなかったマウスよりも結腸異形成の発生率が高いことを報告しました(63%対8.3%)。 結腸細胞は、対照細胞と比較して、P. anaerobius に曝露されたときにかなり高いレベルの増殖を示した。 機械論的に、P. anaerobius は、トール様受容体2および4(TLR2およびTLR4)を活性化して、コレステロール合成とその後の細胞増殖を増加させる活性酸素種の細胞内レベルを高めることにより、腫瘍形成を促進します[31]。 別の研究では、Longらは、P. anaerobius が正常な腸上皮細胞と比較して結腸直腸癌細胞株に優先的に付着することを示しました。 彼らは、この付着の原因であるP. anaerobius の表面タンパク質である推定細胞壁結合リピート2(PCWBR2)を特定しました。 推定細胞壁結合リピート2は、結腸直腸癌腫瘍で頻繁に過剰発現する受容体であるインテグリンα2/β1に直接結合し、結腸直腸癌細胞のPI3K-Aktシグナル伝達経路の活性化を促進し、NFκBの活性化と細胞増殖の増加をもたらします。 NFκBの活性化は、炎症誘発性反応と、IL-10およびIFN-γP. anaerobius で治療されたマウスのレベルを増加させます。 さらに、P. anaerobius で治療したマウスは、腫瘍随伴マクロファージ(TAM)、顆粒球腫瘍関連好中球、および慢性炎症と腫瘍発生に関与する骨髄由来抑制細胞の集団の有意な増加を示しました[66]。 |
| ポルフィロモナス ジンジバリス |
| P. gingivalisは、大腸に感染し、さまざまな病気の病因に寄与する可能性のある口腔微生物叢です[67]。 マウスにPrevotella intermedia とP. gingivalisを経口投与すると、全身性炎症、内毒素血症、腸管バリアの破壊、腸内毒素症が引き起こされることが示されています[68]。 最近、Muらは結腸直腸癌における.P. gingivalisの役割とその作用機序を調査しました。 彼らは、P. gingivalisが結腸直腸癌細胞に付着して侵入し、それらの増殖を有意に増強し、細胞周期のS期にある細胞の割合を増加させる可能性があることを発見しました。 機械論的に、P. gingivalisは、MAPK/ERK経路を有意に活性化することによって結腸直腸癌細胞の増殖に寄与します[69]。 別の研究では、Wangらは、ApcMin/+マウスへのP. gingivalisの強制経口投与が結腸直腸癌腫瘍形成を促進することを実証しました。 骨髄由来の免疫細胞を優先的に増加させることにより、マクロファージやDCなど、炎症誘発性の状態を誘発し腫瘍微小環境(TME)を調節します。 彼らはまた、P. gingivalisがヌクレオチド結合オリゴマー化ドメイン様受容体ファミリーのパイリンドメイン含有3(NLRP3)インフラマソームを活性化することによって結腸直腸癌を促進することを示した[32]。 |
| エンテロコッカス フェカーリス |
| 結腸直腸癌におけるE. faecalisの役割については、物議を醸すデータがあります。一部の研究では、結腸直腸癌に対する役割または保護的役割が示されていませんが、他の研究では有害な役割が示されています[70]。 たとえば、De Almeida et alは、健康な症例と比較して、結腸直腸癌患者の糞便中のE. faecalisの存在量が減少していることを発見しました(22.2%対44.4%)[35]。 同様に、Viljoen et alは、E. faecalisの感染と結腸直腸癌との間にかなりの臨床的関連性を発見しませんでした[71]。 一方、Wang et alは、E. faecalisがWnt/β-カテニン経路の活性化と多能性転写因子の誘導を介して結腸直腸癌の誘導に寄与することを示しました。 彼らは、結腸上皮細胞をE. faecalisに感染したマクロファージに曝露すると、染色体の不安定性、遺伝子変異、および細胞の形質転換を通じて結腸直腸癌の開始が促進されることを示しました[72]。 |
| ストレプトコッカス ガロリチカス |
| 以前はS. bovisと呼ばれていた S. gallolyticusのコロニー形成は、結腸直腸癌の発症と有意に相関していることが示されています[73、74]。 109例のケースコントロール研究では、 S. gallolyticusが対照症例よりも結腸直腸腫瘍患者で有意に高かったことが明らかになりました(70%対32%)[75]。 機械論的に、 S. gallolyticus抗原は、COX-2発現を誘導することによって細胞増殖と血管新生を促進する可能性があります[76]。 さらに、 S. gallolyticusの壁抽出抗原は、結腸直腸癌細胞株に結合することによって炎症性サイトカインの産生を誘導します[77]。 |
| 腸内細菌叢と化学療法の意味 |
| 化学療法に対する宿主の反応は、免疫調節、微生物転座、誘発された生態系多様性、酵素分解、代謝などのさまざまなメカニズムを通じて腸内細菌叢で調節できるという新たな証拠があります[78]。 |
| Viaudらは、マウスモデルの小腸微生物叢の組成に対するシクロホスファミド(CTX)とドキソルビシンの効果を調査しました。 彼らは、両方の化学療法剤による治療が、小腸絨毛の短縮、腸上皮バリアの不連続性、炎症細胞の蓄積、および共生細菌の二次リンパ器官への移行につながることを発見しました。 シクロホスファミドによる治療は、7日後にマウスの小腸の総細菌数を減少させませんでしたが、腸球菌と乳酸桿菌の存在量は減少を示しました。さらに、Enterococcus hirae、Lactobacillus murinus、Lactobacillus johnsoniiなどの一部のグラム陽性菌は、病原性のTヘルパー17(pTh17)および1型Tヘルパー(Th1)細胞の生成を刺激しました。 一方、抗生物質でこれらの細菌を殺すと、病原性のTヘルパー17応答が減少し、シクロホスファミドに対する腫瘍耐性が強化されました[79]。 別の研究では、Daillèreetalは、Barnesiella intestinihominisとEnterococcus hiraeがシクロホスファミドに対する宿主の応答に関与していることを示しました。彼らは、B.intestinihominisが結腸に蓄積し、IFN-γ産生γδT細胞の腫瘍内回復を誘導するのに対し、E.hiraeは小腸から二次リンパ器官に移行し、腫瘍微小環境のCD8/Tregの比率を増加させることを発見しました。 結腸のB.intestinihominisは効果のオーケストレーターとして機能し、E. hiraeはシクロホスファミドによって誘発された抗腫瘍効果を回復する可能性があります[80]。 Yuらは、オートファジーを活性化することにより、F. nucleatumが5-フルオロウラシル(5-FU)およびオキサリプラチンに対する結腸直腸癌細胞の耐性を促進できることを実証しました。 メカニズムの観点から、F. ucleatumはTLR4とMYD88を含み、miR-4802とmiR-18a*の発現をダウンレギュレーションしました。 miR-4802およびmiR-18a*レベルの低下は、それぞれATG7およびULK1のアップレギュレーションにつながり、オートファジーの活性化をもたらします[81]。 腸内細菌叢と化学療法の調節の別の例は、トポイソメラーゼI阻害剤であるSN-38の形で活性化されるプロドラッグであるイリノテカンの副作用と代謝の制御です。 宿主の肝酵素はグルクロニドSN-38を不活性化型(SN-38G)に変換する可能性があります。 腸に到達した後、SN-38Gは細菌のβ-グルクロニダーゼ酵素を介して加水分解されてSN-38に戻り、重度の下痢と腸の損傷を引き起こします[82]。 |
| 腸内細菌叢と免疫療法の意味 |
| 癌免疫療法のアプローチは、患者の免疫応答が抗腫瘍効果を発揮するように誘発される癌を治療するための有望な治療方法になりました。 腸内細菌叢が免疫療法への反応を調節する可能性があるという証拠があります。 Eubacterium limosum、Alistipes shahii、B.fragilis、Akkermansia muciniphila、Faecalibacterium sppなどの特定の腸内細菌が免疫療法における腫瘍反応を高める可能性があることが示されています[83–87]。 たとえば、ある研究の結果から、Bifidobacteriumの存在量が大幅に増加すると、腫瘍浸潤リンパ球(TIL)のレベルが上昇し、プログラムされた細胞死タンパク質1リガンド1(PD-L1)抗体による治療に対する腫瘍の反応が促進されることが明らかになりました [88]。 最近、Shiらは、腸内細菌叢が腫瘍微小環境と腸内免疫を変化させることにより、抗CD47免疫療法の有効性に影響を与える可能性があることを示しました。 Bifidobacteriumは腫瘍微小環境内に蓄積し、非応答性腫瘍を応答性腫瘍に変換し、IFNおよびインターフェロン遺伝子刺激因子(STING)依存的に抗CD47による免疫療法を行う可能性があります[89]。 別の研究では、Lvらは、マイクロサテライトの安定性を備えた結腸直腸癌での腸内細菌叢のリモデリングによってPD-1遮断の有効性が高まる可能性があることを発見しました。 彼らは、伝統的な中国の薬であるGegen Qinlian煎じ薬(GQD)が、抗PD-1抗体による免疫療法の有効性を高め、in vivoでの腫瘍増殖を阻害する可能性があることを発見しました。 Gegen Qinlian煎じ薬と抗PD-1の組み合わせは腸内細菌叢を調節し、Bacteroides acidifaciensとBacteroidales S24-7属を増加させます。 併用療法はまた、腫瘍組織および末梢血におけるCD8 + T細胞の割合を著しく増加させ、IFN-γおよびIL-2の発現を上昇させました。 機械論的に、Gegen Qinlian煎じ薬と抗PD-1の組み合わせは、グリセロリン脂質とスフィンゴ脂質の経路の代謝を増加させました[90]。 両方の代謝物が結腸直腸癌患者のモニタリングにおいてバイオマーカーとして機能する可能性があることが示されています[91、92]。 |
腸内細菌叢調節の戦略 |
| 腸内細菌叢および微生物分泌代謝物が結腸直腸癌との闘いにおける治療戦略として標的とされる可能性があるという証拠があります。 たとえば、Bhalla et alは、葉酸などの微生物代謝物が結腸直腸癌細胞の生存率を抑制できることを示しました。 したがって、抗がん代謝物を生成するための腸内細菌叢の調節は、結腸直腸癌の治療に使用できます[93]。 次のセクションでは、治療戦略として腸内細菌叢を調節するために適用されてきた戦略について考察します。 |
| プロバイオティクス |
| プロバイオティクスは、細菌、酵母、カビなどの微生物であり、適切な量で送達されると宿主の健康を改善することができます。 Lactobacillus, Bacillus, Bifidobacterium, Streptococcus, および Enterococcus は、プロバイオティクスとして最も使用されている細菌属です[94]。 プロバイオティクスは、1)免疫調節、2)病原菌のコロニー形成の阻害、3)腸のバリア機能の強化という、3つの主要なメカニズムを通じて結腸直腸癌の予防と治療において腸に影響を与えます。 図2は、腸内細菌叢の調節に適用される戦略と、各戦略の作用機序を表しています。 |
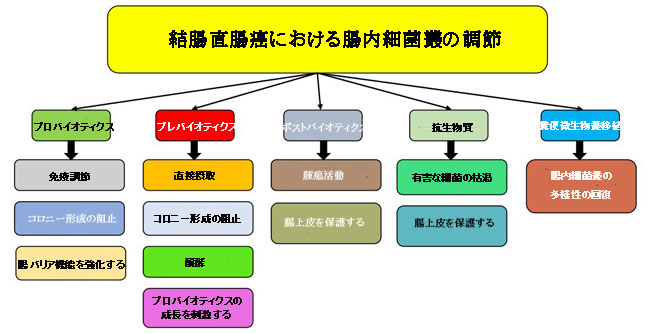 |
| 図2 結腸直腸癌における腸内細菌叢の調節に適用される戦略と各戦略の作用機序 |
| プロバイオティクス:プロバイオティクスは、免疫調節、病原菌のコロニー形成の阻害、および腸バリア機能の強化を通じて、腸内細菌叢の組成に影響を及ぼします。 プレバイオティクス:腸内細菌による直接取り込みと抗炎症作用、病原菌との相互作用による病原菌のコロニー形成の防止、腸内細菌叢による発酵、および有益な腸内細菌の刺激を通じて、腸内細菌叢の調節要素として機能します。 ポストバイオティクス:腫瘍細胞を選択的に阻害し、上皮細胞のアポトーシスを阻害してIgA分泌を増加させることにより腸上皮を保護することにより、殺腫瘍機能を発揮します。 抗生物質:抗生物質は、有害な細菌から腸を枯渇させ、腸上皮を保護する可能性があります。 糞便微生物叢移植:この戦略は、腸内の微生物叢の多様性を回復するのに役立ちます。 |
| 腸内毒素症の状態は、炎症性腸疾患(IBD)および結腸直腸癌の発生につながる一酸化窒素(NO)および炎症性サイトカインとしてのIL-8の産生を増加させる経路およびMAPKやNF-κBなどの転写因子を活性化する可能性があります。 プロバイオティクス投与が、調節性T(Treg)細胞を介してTGF-β2やIL-10などの抗炎症性サイトカインの分泌を増加させることにより腸内細菌叢のバランスを再確立するという証拠が蓄積されています[95]。 たとえば、Bifidobacterium breveおよびBifidobacterium infantisは、結腸樹状細胞を活性化し、Foxp3 + TregおよびIL-10産生1型Treg(Tr1)を発現させる可能性があります[96、97]。 Leeらは、プロバイオティクス特性を含むBacillus polyfermenticusが、TNF-α、NO、COX-2などの炎症性サイトカインのレベルを低下させることにより、CRC細胞株HT-29およびLoVoに対して抗増殖効果を示すことを実証しました[98 ]。 一方、プロバイオティクス細菌は炎症誘発性反応を誘発する可能性があることが示されています。 Hradicka et alは、マクロファージ細胞のin vitroモデルにおけるプロバイオティクス特性を含む6つの乳酸桿菌の免疫調節効果と、結腸直腸癌のラットモデルにおけるそれらの抗腫瘍特性を研究しました。 彼らは、乳酸桿菌とM1およびM2様マクロファージとの共培養において、細菌がIL-1β、TNF-α、IL-18、およびIL-23を含む炎症誘発性および抗炎症性サイトカインの放出を誘発する可能性があることを示しました。 一方、細菌の経口投与は、腫瘍の多様性、数、および体積の減少、ならびに結腸の長さの回復およびIL-18の産生の増加をもたらしました[99]。 別の研究では、臨床試験により、プロバイオティクスSaccharomyces boulardiiの経口投与により、IL-10、IL-23A、IL-1βなどの炎症性サイトカインと抗炎症性サイトカインの両方のレベルがダウンレギュレートされる可能性があることがわかりました。 一方、プロバイオティクスを投与された患者の感染性合併症の発生率は、対照群の38.8%と比較して13.3%でした[100]。 |
| プロバイオティクスの他の有益な特性は、腸を分解して有毒な化合物を放出する病原菌のコロニー形成の抑制です。 プロバイオティクスは、管腔のpHを下げ、抗菌ペプチドを生成し、腫瘍形成促進酵素を減らすことにより、有害な微生物の増殖を抑制する可能性があります[101]。 たとえば、Fayol-Messaoudi et alは、Lactobacillusのプロバイオティクス菌株が非乳酸および乳酸物質を生成し、続いてpHを下げることによって病原菌であるSalmonella entericaの増殖を阻害する可能性があることを示しました [102]。 さらに、プロバイオティクスがStaphylococcus aureusやClostridium difficileなどの病原菌による結腸感染を予防できるという証拠があります[103、104]。 プロバイオティクスはまた、腸への病原体の付着を阻害する可能性があります。 Collado et alは、プロバイオティクス菌株が付着した病原体を追い出すか、腸粘液への病原体の付着を阻害する可能性があることを発見しました。 一方、プロバイオティクスのさまざまな組み合わせは、アドヒアランス阻害の割合が高いことを示しました[105]。 別の研究では、Behbahani et alは、プロバイオティクスL.plantarum株L15の腸細胞への接着能力を明らかにしました。 この菌株は、E.coliに対して、競合、阻害、置換機能などの抗付着活性を示しました[106]。 |
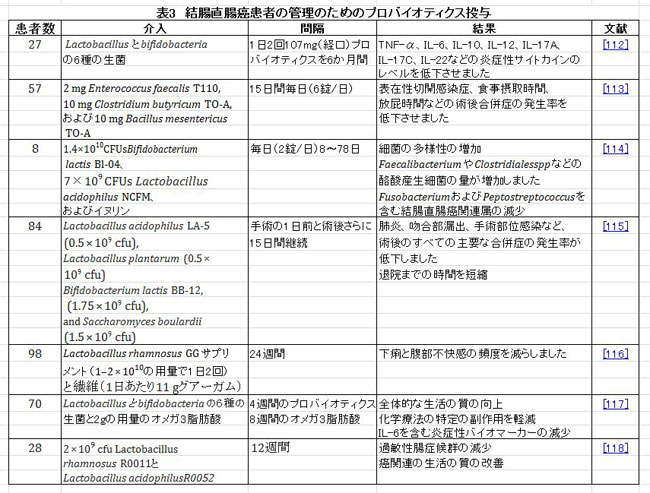
| 腸上皮は物理的な障壁として機能し、病原性毒素、ストレス要因、および細菌から薄層環境を保護します。 バリア機能には、タイトジャンクション(TJ)、接着タンパク質ジャンクション(AJ)、デスモソームの3つの要素があります。 この物理的障壁の破壊は、炎症性腸疾患および炎症性腸疾患関連結腸直腸癌の発症、ならびに腫瘍の浸潤および転移の加速をもたらすことが示されている[107、108]。 いくつかのプロバイオティクス菌株が腸バリアの機能を改善するという証拠があります。 臨床試験の研究では、手術を受けている結腸直腸癌患者に術前と術後の両方でプロバイオティクスを経口投与すると、経上皮抵抗が増加し、タイトジャクションの発現がアップレギュレートされる可能性があることが明らかになりました。 プロバイオティクスを投与された患者は、糞便中の細菌の多様性が増加し、血液中の腸内病原菌が減少しました[109]。 Liuらは、手術後の結腸直腸癌患者の腸バリアに対するプロバイオティクスの効果を調査するためにメタアナリシス研究を実施しました。 17の研究の評価は、プロバイオティクス投与が腸透過性、オクルディンレベル、分泌型免疫グロブリンA(SIgA)レベルを増加させ、細菌の移動を減少させることにより、腸の物理的および生物学的障壁を保護できることを示しました[110]。 別の研究では、 L. acidophilus, B. bifidum, および B. infantum (LBB療法)を含むプロバイオティクス株の投与は、腫瘍数、腫瘍サイズ、および腫瘍発生率を低下させることにより、結腸直腸癌の発症を阻害する可能性があります。 LBB療法はまた、腸内細菌叢を変化させ、病原菌の存在量を減少させました。 さらに、LBB療法は、オクルディン、閉鎖帯-1(ZO-1)、およびムチン2(MUC2)の発現を増加させることにより、腸上皮および粘膜バリアの完全性を高めました。 LBB治療はまた、TLR2の発現を増強し、TLR4、COX2、カスパーゼ-3、およびβ-カテニンのダウンレギュレーションを介してCRCを予防しました。 これらの結果は、プロバイオティクスLBB療法の抗がん効果は、TLR2依存性の腸バリアの増強と、炎症、アポトーシス、およびβ-カテニン経路の阻害によるものであることを示唆しています[111]。 表3は、結腸直腸癌患者の管理におけるプロバイオティクスの適用をまとめたものです。 |
 |
| プレバイオティクス |
| プレバイオティクスは、「腸内細菌叢の組成および/または宿主の健康と健康に利益をもたらす活動に特定の変化を引き起こす選択的に発酵させた成分」です[119]。 ポリフェノール、多価不飽和脂肪酸(PUFA)、およびガラクトオリゴ糖(GOS)、キシロオリゴ糖(XOS)、フラクトオリゴ糖(FOS)、フルクタン、イヌリンなどの炭水化物は、プレバイオティクス特性を備えています。 プレバイオティクスは、さまざまなメカニズムを通じて機能を発揮します。 (1)有益な腸内細菌の刺激、 (2)腸内細菌叢による発酵、 (3)腸による直接摂取と抗炎症作用の発揮、および (4)病原体と相互作用することによる病原体のコロニー形成の防止[5]。 |
| プレバイオティクス投与は、Akkermansia, Rosebura, Ruminococcus, および Faecalibacteriumを含むプロバイオティクスの豊富さを高める可能性があることが示されています[120、121]。 Zhengらは、C.butyricum(プロバイオティクスとして)と化学修飾デキストラン(プレバイオティクスとして)を組み合わせたプレバイオティクスカプセル化プロバイオティクス胞子(spores-dex)を調製し、それらの抗がん効果を結腸腫瘍モデルで評価しました。 彼らは、経口投与後の胞子-dexによる結腸癌の濃縮を示した。 C. butyricum による病変内のデキストランの発酵は、抗癌活性を有する短鎖脂肪酸(SCFA)の産生をもたらしました。 さらに、spores-dexは、RoseburiaやEubacteriumなどの短鎖脂肪酸産生細菌を増加させ、腫瘍の成長を大幅に阻害する可能性があります。 短鎖脂肪酸産生菌は、腸内に腫瘍抑制微小環境を構築することにより、腫瘍抑制に寄与します[122]。 短鎖脂肪酸として、酪酸は結腸細胞の主要なエネルギー燃料であり、酢酸塩とプロピオン酸塩はエネルギー生成と糖新生のために筋肉と肝臓によって代謝されます[123]。 さらに、酪酸は結腸直腸癌アポトーシスを誘発し、酸化ストレスを調節し、上皮バリアを強化し、炎症をダウンレギュレーションする可能性があります[124]。 ヒストンデアセチラーゼ阻害および細胞内代謝活性に加えて、短鎖脂肪酸は、GPR43(FFA2)、GPR41(FFA3)、およびGPR109Aと呼ばれる腸内のGタンパク質共役受容体(GPCR)を介してほとんどの機能を発揮します。 たとえば、結腸上皮のFFA2に酪酸、酢酸、プロピオン酸が結合すると、シグナル伝達カスケードがトリガーされ、細胞周期の停止、アポトーシスの促進、炎症の抑制につながります。 FFA2の発現は、 Bifidobacterium属の成長を促進し、Prevotellaceae属とH.hepaticusを阻害します。 一方、酪酸を産生するButyricicoccus pullicaecorumは、FFA2の発現を増加させることにより抗腫瘍形成機能を示します[125]。 |
| プレバイオティクスはまた、病原菌の上皮細胞および腸への付着を妨害します。 たとえば、Ribeiro et alは、プレバイオティクス特性を備えたオリーブ搾りかす粉末が、微生物叢による短鎖脂肪酸の生成を促進するだけでなく、病原体に対する抗酸化抗接着活性も示すことを示しました。 彼らは、不溶性食物繊維の主要な供給源としてのパルプ濃縮粉末が、B. cereus および L. monocytogenesの付着をそれぞれ最大22%と20%阻害することを実証しました[126]。 別の研究では、Leongらは、ヤギの乳児用調製粉乳に含まれるオリゴ糖のプレバイオティクス特性とその作用メカニズムを調査しました。 彼らは、プレバイオティクスオリゴ糖がlactobacilliaとbifidobacteriaの増殖を著しく促進し、結腸直腸癌細胞株Caco-2へのS.typhimuriumとE.coliNCTC10,418の付着を減少させる可能性があることを報告しました[127]。 ヤギの乳汁中のオリゴ糖と腸表面の炭水化物との構造的類似性のため、これらのオリゴ糖、特にフコシル化およびシアル化されたオリゴ糖は、宿主細胞受容体の可溶性類似体として作用するか、そのような構造を変化させることにより、腸上皮細胞への病原菌の付着を減少させます`式[128、129]。 |
| ポストバイオティクス |
| ポストバイオティクスは、細胞分画、不活化微生物細胞、および発酵中にプロバイオティクス生細胞で作られた細胞代謝物として定義され、宿主にとってさまざまな健康上の利点が含まれています。 ポストバイオティクスは細菌培養の馴化/上清培地に存在するため、生存可能な微生物よりも安全です。 ポストバイオティクスは、以下によって抗腫瘍活性を発揮します。 1)腫瘍細胞を選択的に阻害し、 2)上皮細胞のアポトーシスを抑制し、IgA分泌を増加させることで腸上皮を保護します[5]。 |
| L. plantarumなどの一部の細菌によって産生される生物後代謝物は、結腸直腸癌細胞を含む腫瘍細胞に対して細胞毒性および抗増殖効果を示すことが示されています[130、131]。 Leeらは、3D培養システムで結腸直腸癌細胞に対してL.fermentumを使用したプロバイオティクス無細胞上清処理の殺腫瘍機能を評価しました。 彼らは、細菌培養上清が、Bax、Bak、Bid、Noxa、およびカスパーゼ-3をアップレギュレートすることによって結腸直腸癌細胞株のアポトーシスを誘導できることを発見しました[132]。 別の研究では、小西らは、結腸癌細胞に対するL.caseiATCC334の培養上清中のフェリクロームの腫瘍抑制効果を実証しました。 フェリクロームは、c-jun N末端キナーゼ(JNK)を介したアポトーシスを誘導することにより、癌細胞の生存率を低下させました。 さらに、癌細胞に対するフェリクロームの殺腫瘍効果は5-FUおよびシスプラチンよりも大きかったが、非癌性腸細胞に対するフェリクロームの毒性は化学療法剤よりも小さかった[133]。 |
| ポストバイオティクスの他の作用機序は、腸の炎症を抑制し、腸のバリアの完全性を維持する能力に関連しています。 たとえば、Izuddin et alは、離乳後の子羊にL. plantarum RG14由来のポストバイオティックを補給すると、ルーメン上皮の成長、TNFやIL-1βなどの炎症性サイトカインおよびタイトジャンクションプロテイン1(TJP1)、クローディン-1(CLDN-1)、およびCLDN-4のアップレギュレーションによる腸バリアの完全性の改善のダウンレギュレーション、および抗炎症性サイトカインIL-10が生じることを示しました [134]。 別の研究では、Gaoらは、分泌タンパク質であるHM0539が、L. rhamnosus GGの培養に有益な効果があることを確認しました。 彼らは、HM0539が腸のムチンの発現を増加させ、腸のバリア損傷を防ぐことにより、腸のバリアの完全性を維持する上で保護的な役割を果たすことを発見しました[135]。 腸上皮に影響を与えるL. rhamnosus GGのもう1つの生物後タンパク質はp40タンパク質です。 分解から保護するために、YanとPolkはハイドロゲルコーティングp40を使用し、p40タンパク質が保護免疫応答を促進し、腸上皮細胞のアポトーシスを減少させ、上皮成長因子(EGF)受容体依存性マナーの結腸のバリア機能を保護することを示しました [136]。 |
| 抗生物質 |
| 抗生物質の使用は、細菌の生物多様性の低下、抗生物質耐性菌の選択など、腸内細菌叢に悪影響を与えることが示されています。 代謝機能の変化により、抗生物質関連下痢と C. difficile感染症の再発が起こります[137]。 抗生物質が結腸直腸癌リスクを高めるという証拠が増えていますが[120、121]、腸内細菌叢を操作することで腫瘍のサイズと数を減らすこともできます[138、139]。 たとえば、DeStefano Shields et alは、セフォキシチン抗生物質による治療が、腸管腫瘍マウスモデルにおける腸管毒素原性B.fragilisのコロニー形成を完全かつ永続的に除去できることを報告しました[140]。 |
| 抗生物質は、腸の粘膜バリアを維持する上でも保護的な役割を果たします。 赤身の肉を大量に摂取すると結腸直腸癌のリスクが高まるため、Ijssennaggerらは、赤身の肉の色素および代用物としてのヘムが腸内細菌叢および結腸直腸癌の発生に及ぼす影響を調査しました。 彼らは、ヘムを含む食餌を与えられたマウスは、腸上皮の損傷と過剰増殖を示し、結腸癌を引き起こしたが、ヘム+抗生物質療法を投与されたマウスは、上皮の損傷と過剰増殖を示さなかったことを示した。 機械論的に、微生物によって生成された硫化水素は、粘液バリアを開くことによって腸上皮を細胞毒性ヘムにさらします。 抗生物質は微生物の硫化物の生成を抑制し、それによって誘発された過剰増殖を防ぐ粘液バリアの完全性を維持します[141]。 上記のメカニズムにもかかわらず、腸内細菌叢、抗生物質、および結腸直腸癌の関係は非常に複雑であり、この関係と結腸直腸癌療法への応用を決定するために注意深く研究する必要があります。 |
| 糞便微生物叢移植 |
| 興味深く最も革新的な生物療法法である糞便微生物移植(FMT)は、健康な個人から患者に便移植を移すこととして定義されています。 糞便微生物移植はC. difficile感染症(CDI)の治療に承認された方法ですが[142]、肥満、炎症性腸疾患、糖尿病、非アルコール性脂肪性肝疾患、および心血管疾患の治療にも有望な可能性を示しました。 前向き臨床試験研究は、糞便微生物移植が血液障害のある患者の抗生物質耐性菌(ARB)の腸内コロニー形成を阻害する可能性があることを明らかにしました。 糞便微生物移植は、患者からの抗生物質耐性菌の完全な(75%)および部分的な(80%)脱植民地化をもたらしました[143]。 糞便微生物移植は腸内細菌叢の多様性を回復しようとします。 糞便微生物移植は、無病で健康な微生物集団を不均衡なコミュニティに導入することによって微生物の恒常性を回復し、C. difficile感染症、炎症性腸疾患、過敏性腸症候群などのいくつかの胃腸障害を改善するための有用なツールとして機能することが示されています[144]。 Rosshart et alは、実験用マウスへの野生マウスの糞便移植が、適応を促進し、変異原/炎症剤による結腸直腸腫瘍形成誘導に対する耐性を改善できることを示した[145]。 興味深いことに、Wongらは、結腸直腸癌患者の糞便微生物叢が、発癌性物質としてアゾキシメタンを投与された従来の無菌マウスで腫瘍形成を増加させ、微生物の存在量を減少させたことを示しました。 マウスはまた、Th1細胞とTh17細胞の割合を増加させ、C-X-Cモチーフケモカイン受容体1(CXCR1)、CXCR2、IL-17A、IL-22、およびIL-23をアップレギュレートしました[146]。 別の研究では、Sobhaniらは、結腸直腸癌患者から無菌マウスへの新鮮な糞便の移植が、結腸直腸癌患者の変化パターンと同様にいくつかの遺伝子の高メチル化を誘発する可能性があることを示した[147]。 糞便微生物移植の臨床試験は、抗PD-1療法(NCT04729322)への非応答者における転移性結腸直腸癌の治療のために募集されています。 |
結論 |
| 結腸直腸癌状態での腸内細菌叢の調節に関するいくつかのレビュー研究がありますが、治療戦略の効率と結果に対する腸内細菌叢の影響を包括的に研究しているレビューはほとんどありません。 ここでは、腸内細菌叢の変調がどのように結腸直腸癌の開始と進行につながるかについてのメカニズムの概要を把握しようとしました。 腸内毒素症は一般に結腸直腸癌発癌で発生するため、プロバイオティクス、プレバイオティクス、ポストバイオティクス、抗生物質、糞便微生物移植など、腸内細菌叢を変化させるためのさまざまな治療戦略が開発されています。 これらの戦略は、免疫調節機能、腸バリアの完全性の維持、腸内毒素症の回復、殺腫瘍活性、コロニー形成抵抗性、抗がん製品の製造など、さまざまなメカニズムを介して結腸直腸腫瘍治療を管理します。 有望な結果にもかかわらず、腸内細菌叢の調節に適用される戦略に関連するいくつかの懸念があります。 たとえば、日和見病原体または病原性因子の存在、および腸内微生物集団における耐性の原因となる遺伝子の拡散は、プロバイオティクス戦略の安全上の懸念事項です。 さらに、腹痛、軽度の発熱、下痢、鼓腸、疲労感、倦怠感などの副作用が糞便微生物移植の主な課題です。 したがって、信頼性が高く包括的な結果を達成するために、長期試験および大量のサンプルを使用した各戦略のリスクとベネフィットの可能性の評価を研究に含める必要があります。 |
参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
この文献は、Journal of Translational Medicine (2022) 20:178に掲載されたGut microbiota modulation: a tool for the management of colorectal cancer.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |
結腸直腸癌の腸内細菌叢 |