ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(発酵乳、腸内細菌の科学:研究の最前線) |
ケフィアからの生物活性化合物と健康に対するそれらの潜在的な利点 :系統的レビューとメタ分析 |
Carla P. Vieira et al., Oxidative Medicine and Cellular Longevity Volume 2021, Article ID 9081738, 34 pages |
|
|
概要 |
| ケフィア投与による健康上の利点の証拠にもかかわらず、これらの利点に関連する生物活性化合物のメタアナリシスによる系統的レビューはまだ文献にありません。ケフィアは、共生における複雑な微生物相の代謝から生じる発酵乳です。最近の研究では、ケフィアに起因する予防および治療効果の原因となる生物活性化合物が調査されています。ただし、産業用ケフィアと職人技のケフィアの機能的可能性の違いについては、まだ議論の余地があります。まず、両方のタイプのケフィアの間で微生物組成の違いを特定しました。その後、invitroおよびinvivo研究の両方から、ケフィアからのさまざまな生物活性化合物の健康への作用に関する入手可能な証拠を要約して、効果の用量と介入時間、生産微生物、乳汁中の前駆体、およびアクションメカニズムの主要な結論を導き出しました。 メタアナリシスを実施して、介入と対照の間、および研究した各健康影響について両方のタイプのケフィアの間の統計的に有意な差(P <0.05)を調査しました。要約すると、より一般的に報告されている生物活性化合物は、ケフィラン、生物活性ペプチド、および有機酸、特に乳酸を含む細胞外多糖類でした。ケフィアの生物活性化合物は、メタアナリシスによって裏付けられた抗菌、抗癌、および免疫調節活性を示しました。ただし、これらの生物活性化合物の実用性を強化するには、臨床的証拠が緊急に必要です。 それらの作用のメカニズムは多様であり、それらが異なるシグナル伝達経路によって作用できることを示しています。 それでも、観察された健康への影響によると、機能的可能性に関して産業用ケフィアと職人用ケフィアは異なる可能性があります-OR 8.56(95%CI:2.27-32.21、P≤.001)、 これは、両方のタイプのケフィア間の微生物組成の違いに関連している可能性があります。 |
1.はじめに |
|
マトリックスの発酵はケフィアを生成します。 ミルクは一般的に使用されるマトリックスであり、酸性でわずかにアルコール性で、クリーミーな粘稠度の飲料になります[1]。 これは、ケフィア粒に共生している微生物による乳汁発酵の結果です。 ケフィアは、微生物の多様性の代謝結果であるため、他の発酵乳とは異なります。 乳糖発酵および非発酵酵母種(Kluyveromyces、Pichia、およびSaccharomyces)、 乳酸菌(Lactobacillus, Lactococcus, Leuconostoc, および Streptococcus)が優勢で、酢酸菌[2]に加えて、グレインの微生物相を構成します。 |
| 近年、飲料消費と治療効果との関連を動機とするケフィアに関する科学的研究が増加しています[3]。ケフィアの定期的な摂取は、炎症性腸疾患の重症度の軽減[4]、抗高血圧効果[5]、抗発癌効果[6]、インスリン感受性の増加[7]、脂質プロファイルの改善[8]、骨粗鬆症の治療効果[9]、および神経変性疾患[10]に関連しています。 健康へのプラスの効果は、ケフィア飲料による抗酸化能力[11]と腸内細菌叢の調節[12]に関連しています。ケフィアに存在する生物活性化合物は、飲料の発酵および貯蔵中に微生物によって生成され、これらの利点に起因しています。ケフィラン、エキソポリサッカライド、生物活性ペプチド、および有機酸は、ケフィアの治療可能性に一般的に関係している生物活性化合物です[13–17]。しかし、ケフィア飲料に含まれる生理活性化合物[13]について、各疾患の治療可能性に応じて区別するために、さらに深く議論する必要があります。 |
| さらに、職人によるケフィアと工業用ケフィアの機能的可能性の違いについては、文献で議論の余地があります。 ケフィア粒を使用すると職人技のケフィアが得られ[18]、以前に選択された細菌や酵母種のスターターカルチャーは商業用または工業用のケフィアになります[19]。 いくつかの研究では、微生物学的多様性が大きいため、治療の可能性が高い職人技のケフィアが報告されています[20、21]。 対照的に、他の研究では、工業用ケフィアは病気の治療に有望であると説明されていますが[22]、Ebner et al [23]は両方の間に有意差は見られませんでした。 この文脈では、メタアナリシスは、研究間で観察された矛盾を解明するのに役立つ可能性があります。 |
| したがって、近年、ケフィアからの生物活性化合物と健康上の利点に関するいくつかの報告があります。ただし、この研究分野にはまだ概要と詳細なアプローチが不足しているため、メタアナリシスによる系統的レビューがこの目的に関連します。したがって、微生物叢を構成するさまざまな微生物によって生成されたケフィアからの生物活性化合物、それらの有益な効果、作用メカニズム、およびミルク中のそれらの前駆体を要約する必要があります。この知識に基づいて、特定の病気を予防するために食品業界が機能性製剤を開発するための理論的基礎を提供することが可能になります。さらに、製薬業界は、ケフィアから単離された生物活性化合物から治療の可能性を秘めた製剤を調製する可能性があります。このシナリオでは、このレビューは、ミルクのケフィアからの生物活性化合物の抗菌、抗酸化、免疫調節、腸内微生物調節、抗癌、抗骨粗鬆症、抗高血圧、抗糖尿病、および脂質プロファイル調節の役割を要約しました。神経変性疾患の予防と治療もカバーされました。両方のタイプのケフィアの機能の違いがこのレビューを終えました。 |
2.方法 |
2.1 フォーカスの質問 |
質問の作成は、母集団、介入、比較、および結果(PICO)の方法に従って行われました。 作成された質問は次のとおりです。 ミルクケフィアとその生産微生物に含まれる生物活性化合物は何ですか? ミルク中のこれらの生物活性化合物の前駆体は何ですか? ケフィアの主な生物活性化合物の作用機序は何ですか? 健康への同じ効果を得るために、生物活性化合物は濃度と介入時間の点で異なりますか? 職人技のケフィア飲料は、工業的に製造されたものよりも機能的ですか? |
| 2.2データ収集プロセスと適格基準 |
| 2人の著者(C.P.VとA.P.A.C)は、英語で公開された研究論文の特定された要約とタイトルの予備選択を独自に実施しました。結果の原稿の数を制限しないように、タイムフィルターを削除しました。したがって、最初のスクリーニングの出版物は1986年から2021年までの期間をカバーしました。その後、論文が次の健康面のいずれも調査しなかった場合、この最初のスクリーニングで要約が削除されました:抗菌活性、抗酸化活性、癌への影響、神経変性疾患、脂質プロファイル、血圧、血漿グルコース、腸内細菌叢の調節、炎症、および骨粗鬆症の両方で、インビトロ、インサイチュ、インビボ動物、およびヒトの臨床試験。これらの特定の健康上の利点を選択するために使用された基準は、過去20年間に科学文献で最も調査されていたということでした[24–26]。社説、手紙、レビュー、解説、モノグラフ、プレプリント、博士号など、非ミルクケフィアに関する論文は除外されました。論文。論文の全体の読みに基づいて、本研究に含まれるすべての研究は、制御された実験であり、データ分析のための定量的アプローチでした。観察された健康への影響の原因となる生物活性化合物を決定することが可能であった研究のみが含まれていました。ケフィア粒の微生物組成またはミルクケフィアのスターターカルチャーに取り組む研究も含まれていました。従来の治療法、ケフィアの効果に関する最近のレビュー、どの研究ベースにも含まれていなかった研究された疾患の病理学など、現在の研究を構成するために不可欠であると考えられるいくつかの研究が、私たちの紹介と議論を構成するために追加されました。 |
| 最後に、生物活性化合物が由来するケフィアの種類(職人または工業用)、研究モデル、生物活性化合物の定義、および対照処理と比較したその効果に関する情報を要約しました。 結果は、システマティックレビューおよびメタアナリシス(PRISMA)ステートメントの優先報告項目と一致して報告されました。 |
| 2.3 情報源 |
| 私たちの検索プロトコル戦略では、Science Direct、Pubmed、Embase、Web of Science、Scopusの5つの電子データベース用に構築および適合された検索文字列を使用しました。 最初のスクリーニングプロセスは2021年2月から2021年3月まで実施されました。また、関連記事の参照リストをチェックすることにより、直接検索が実行されました。 調査の質問は、原稿が回収された検索文字列を要約するために使用されました。 この文字列は、職人および工業用ミルクケフィアの微生物組成、ミルクケフィア中の生物活性化合物、およびそれらの健康への影響に関連するキーワードの所定のグループに基づいています。 |
| ⅰ)検索コンポーネント1.微生物または「微生物組成」または「ケフィア粒」または「スターターカルチャー」および「ミルクケフィア」 |
| ⅱ)コンポーネント2.「生物活性化合物」または「機能性化合物」および「ミルクケフィア」 |
| ⅲ)検索コンポーネント3.「健康上の利点」または「健康上の効果」または「機能的効果」および「ミルクケフィア」 |
| 2.4 バイアス評価のリスク |
| バイアスの考えられる原因には、適格基準、欠落データの影響、欠落した一次結果、選択されたデータベース、選択された言語(英語)、およびレビューのために選択された記事タイプが含まれます。 |
| 2.5 メタアナリシス |
| メタアナリシスを実施する2人の著者(A.I.L.S.RとD.K.A.R)は、含まれている記事からデータを抽出しました。 取得したデータの不整合は、話し合いによって解決されました。 インビトロ、インサイチュ、およびインビボの論文を試験のためにプールした。 すべての出版物から、それぞれの異なる結果ポイントが独立した研究として抽出されました。 次に、介入と制御の間の統計的に有意な差を調査しました。 ただし、メタアナリシスには、適切な数の研究による健康への影響のみが含まれていました:抗菌剤(微生物の8つのグループを含み、15の論文から合計182のinvitroおよびinsitu研究)、抗酸化剤(149のinvitroおよびinsitu研究が含まれています 7件の記事)、抗がん剤(5件の出版物からなる44件のin vitro研究)、免疫調節(11件の論文から271件のinvitroおよびinvivo研究)、および微生物調節(5件の出版物から取得した40件のinvitroおよびinvivo研究)の効果。 |
| 分析は、さまざまな結果に対する健康上の利点の有無を評価するために実施されました。 選択された研究で異なる方法論が使用されたため、行動の存在/欠如の定義は出版物によって異なりました。 この場合、決議は特定の結果に基づいて3人の著者によって指示されました。 たとえば、抗菌薬の研究を考慮すると、結果がlog UFCで表される場合、ケフィア処理によってlogUFCが減少することは「作用の存在」と見なされました。 同様に、研究結果が抑制ゾーンとして定義された場合、ケフィア治療によるゾーンサイズ(mm)の縮小は「作用の欠如」と見なされました。 |
| さらに、研究間の不均一性を測定するために、I2テストが使用されました。 I2は、すべての研究が同質である、または各研究が同一の効果を測定しているという帰無仮説を仮定しているため、P値はこの仮説をテストします。 このシナリオでは、I2統計は、サンプリングエラー(チャンス)ではなく、不整合(不均一性)による研究全体の変動のパーセンテージを表します。 ここでは、0.05の有意水準を使用しました。 I2≤50%の研究は均質であると見なされました。 I2は次のように定量化されました[27]: |
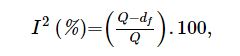 |
| ここで、I2は研究間の不一致、Qはコクランの不均一性統計、dfは自由度です。 |
| 続いて、職人によるケフィアと工業用ケフィアの機能の違いを健康への影響について調査しました。 両方のタイプのケフィアがテストされました:抗菌性(182のinvitroおよびinsitu研究)および抗酸化活性(149のinvitroおよびinsitu研究)。 次に、オッズ比(OR)と対応する95%信頼区間(CI)で評価し、工業用品種ではなく職人技のケフィア飲料を摂取した場合の健康への影響を比較するための研究を分析しました。 すべての分析は、Review Manager 5.4(Cochrane Collaboration、ロンドン、英国)を使用して実行されました。 |
3.結果 |
| 3.1 文献検索 |
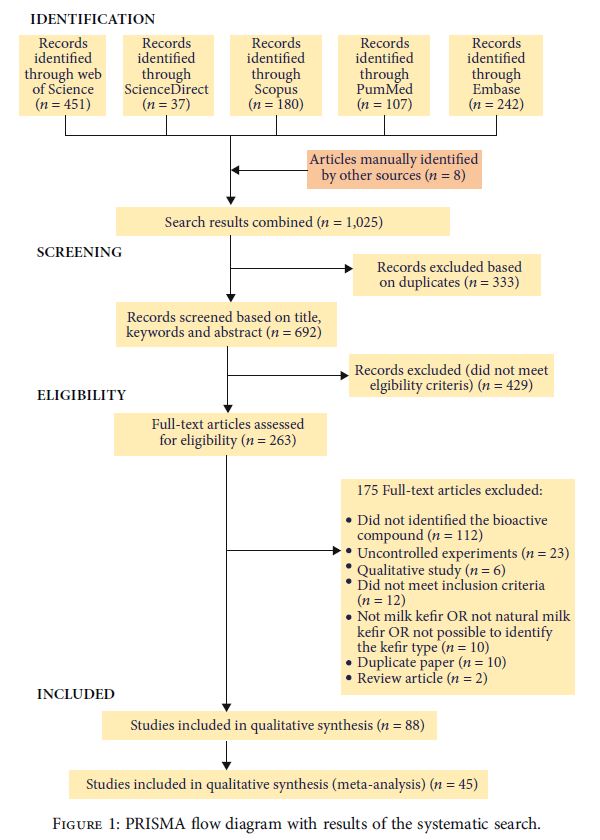
| Web of Scienceで451件、Embaseで242件、Scopusで180件、Pubmedで107件、ScienceDirectで37件の記事が特定されました。 それでも、神経変性疾患への影響を追加するために、他の情報源によって8つの記事が手動で特定されました。 これらのうち、333は重複または三重であり、除外され、692の論文が残っていました。 タイトルと要約を読んだ後、263の論文が完全に読まれるように選択されましたが、適格基準を満たしたのは88のみでした(図1)。 完全に読まれた記事の中で、除外の主な理由は次のとおりでした:生物活性化合物の非同定(n = 112)および制御されていない実験(n = 23)。 これは、より精巧な実験計画を用いた実験の必要性を浮き彫りにし、ケフィアの使用によって観察された健康への影響の原因となる生物活性化合物を特定します。 |
| 図1 体系的な検索の結果を含むPRISMAフロー図 |
 |
| 3.2 メタアナリシス:研究の選択と特徴 |
| メタアナリシスのデータ抽出は45の論文で構成されていました。 調査は、アルゼンチン、ブラジル、カナダ、中国、エジプト、イラン、イタリア、日本、マレーシア、韓国、タイ、台湾、トルコ、英国、およびアメリカ合衆国で実施されました。 研究の発表年は2005年から2021年の範囲でした。研究で使用されたケフィアの濃度は0.01から1000mg / mLの間で変化しました。 介入は0.5から1344時間続いた。 |
| 3.3 ケフィア粒の構造と微生物 |
| ケフィアは一般的にミルクから製造される飲料であり、「ケフィア粒」の微生物学的多様性から複雑な発酵プロセスを伴います[28]。 通常、ケフィア粒は5%から10%の割合でミルクに接種され[29]、スターターカルチャーや接種された微生物の混合に応じてわずかに酸性の味のクリーミーな発酵ミルクなどの特徴があります[1]。 |
| 「ケフィア粒」の多様性は、乳酸菌(LAB)、酢酸菌、菌類、および酵母の間の多数の共生相互作用を包含し[2]、酸性およびアルコール発酵をもたらします[30]。 ケフィアに存在するこれらの微生物は、タンパク質と多糖類で構成されるマトリックスに散在しています[31]。 巨視的には、このマトリックスは、白色から黄色がかった色まで変化する、粘稠で堅固なカリフラワーのような固体の粒子として特徴付けることができます[32、33]。 |
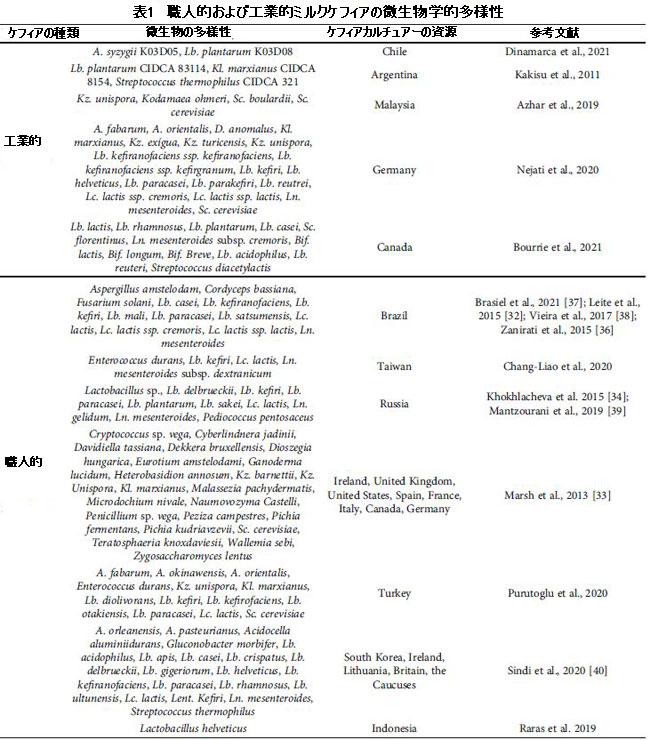
| Khokhlacheva et al [34]によると、適応能力と酵素活性を進化させる微生物間のコンソーシアムは、ミルクケフィアでの微生物の発生と生存の重要なメカニズムです。 ケフィア粒では、乳酸菌(LAB)は通常、酢酸菌と比較してより高いレベルを示します。 この組成は、発酵時間に応じて動的かつ一般的に変化し、発酵の初期段階でラクトバチルス・ケフィラノファシエンスの優勢が観察され、プロセスの最終段階でロイコノストック・メセンテロイデスの最も顕著な成長を引き起こします[35]。 ケフィアの微生物叢(表1)は地理的、気候的、文化的要因によって異なるため、世界中のいくつかの研究がその組成を決定することを目的としています[33、36]。 |
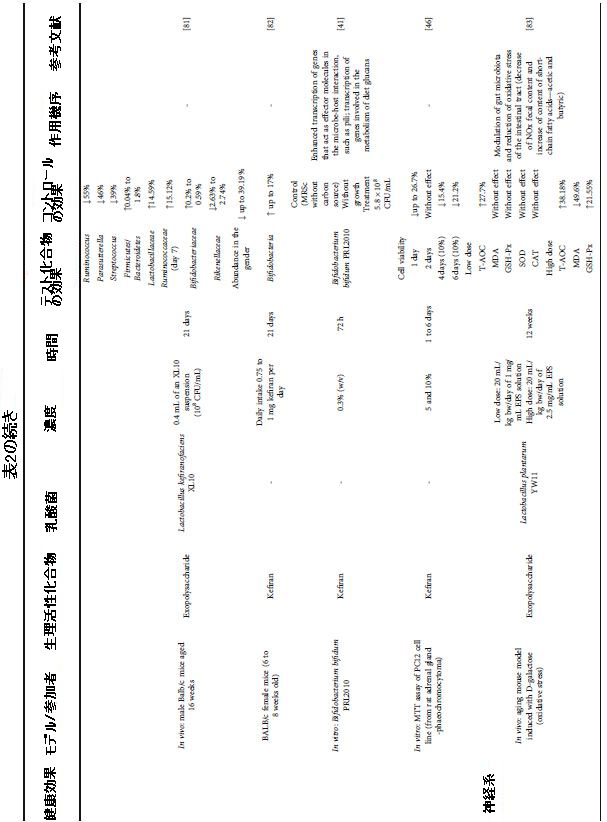
| 表1 職人および工業用ミルクケフィアの微生物学的多様性 |
 |
 |
| 接種されたケフィア粒は職人のケフィアを発酵させ[18]、以前に選択されたバクテリアと酵母種のスターターカルチャーは商業的または工業的なケフィアをもたらします[19]。 表1によると、Lactobacillus属とLactococcus属は両方のタイプのケフィアで優勢ですが、Enterococcusは工業用ケフィアでは報告されていません。 別の重要な側面は、職人のケフィアには一般的に見られない商業種によって工業用ケフィアで指摘することができます:Saccharomyces boulardii、Lactobacillus lactis、Bifidobacterium lactis、Bifidobacterium longum、Bifidobacterium breve、およびLactobacillusreuteri。 Korsak et al [29]はこれらの発見を裏付け、工業用ケフィアは通常、選択されたカルチャーで構成されており、従来は乳製品に使用されていると報告しています。 |
| 真菌の組成に関しては、両方のタイプのケフィアがSaccharomyces cerevisiae、Kluyveromyces marxianus、Kazachstania unisporaなどの一般的な種を示していますが、職人によるケフィアの真菌の多様性は劇的に大きかった。 さらに、Kazachstaniaexígua、Kazachstania turicensis、およびSaccharomyces florentinusは、工業用ケフィアでのみ報告されました(表1)。 両方のケフィアのその他の違いには、栄養素の競合条件下での効率の違い[41]、ケフィア粒の構造における微生物の位置と付着、さらには牛乳で成長する種の能力の違い[18]があります。 |
| ただし、これら2種類のミルクケフィア間の微生物の類似性と特殊性を定義するには、職人技と工業用ケフィアからのより多くの微生物叢の特性評価が必要です。 飲料の生物活性化合物プロファイルは、存在する微生物の生産と密接に関連しているため、この特性評価は非常に重要です[42、43]。 職人の飲料でのケフィア粒の使用は、最終製品の数が多く、胃腸管を通過する際の顕著な生存率と相関しており、工業カルチャーと比較して重要なプロバイオティクス特性を証明しています[42]。 したがって、ここで提案された分析は、両方のタイプのケフィアの間で考えられる機能の違いを正当化する可能性があります。 |
| ケフィア粒の微生物は、ケフィア飲料の発酵および貯蔵中に生物活性化合物を生成する能力を発揮する可能性があります。一貫して、ロシアのケフィア粒から分離された48株から、10種のラクトバチルス属菌が分離されました。プロバイオティクスの可能性が認められました[39]。 Saccharomyces cerevisiae KU200284などの一部の酵母菌株は、スターターカルチャーとプロバイオティクスという2つの重要性を示しています[44]。 韓国のケフィアでは、酢酸細菌株Acetobacter fabarum DH1801は、食品保存メカニズムとプロバイオティクス剤としての可能性を備えた機能的なスターターとしての実行可能性を持っていました[45]。 さらに、LABの種は、ケフィアの重要な生物活性化合物である細胞外多糖(EPS)の形成に基本的な役割を果たします[37]。 このシナリオでは、Lactobacillus kefiranofaciensは、その遺伝子がケフィア粒の構造を構成する細胞外多糖類(ケフィランなど)を生成する大きな能力を示すため[18]、ケフィア粒の形成の主要部分と見なされます[43]。 同様に、Lactococcus lactis ssp cremoris MRS 47は、Vieira et al [38]が、乳脂肪から共役リノール酸(CLA)、生物活性化合物を生成できることを発見しました。 |
3.4 ケフィアからの生物活性化合物とそれらの健康への影響 |
| 3.4.1 抗菌活性 |
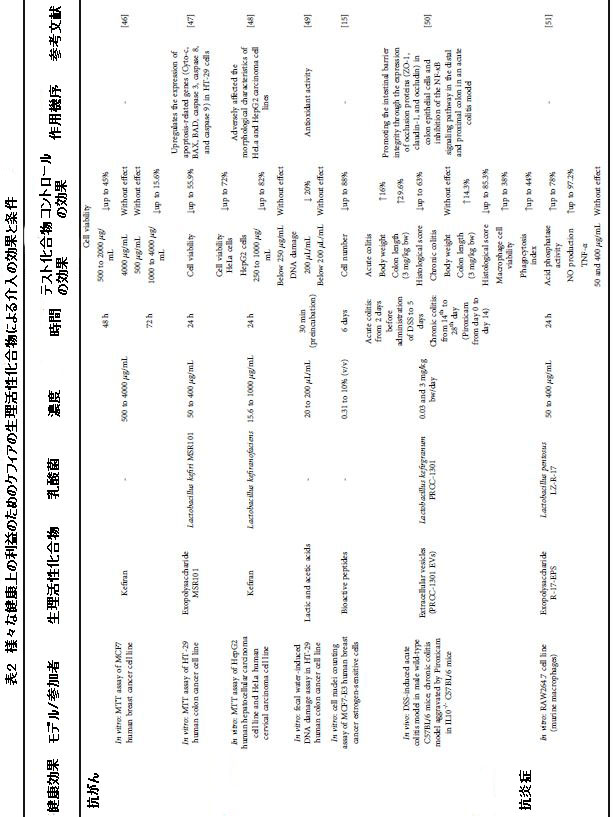
| ケフィアの抗菌活性は、主にエキソポリサッカライド(EPS)、特にケフィラン、および有機酸、特に乳酸に起因していました。 ただし、抗菌活性を持つ生物活性ペプチドも特定されています(表2)。 |
| 表2 さまざまな健康上の利益のためのケフィア生物活性化合物による介入の効果と条件 |
 |
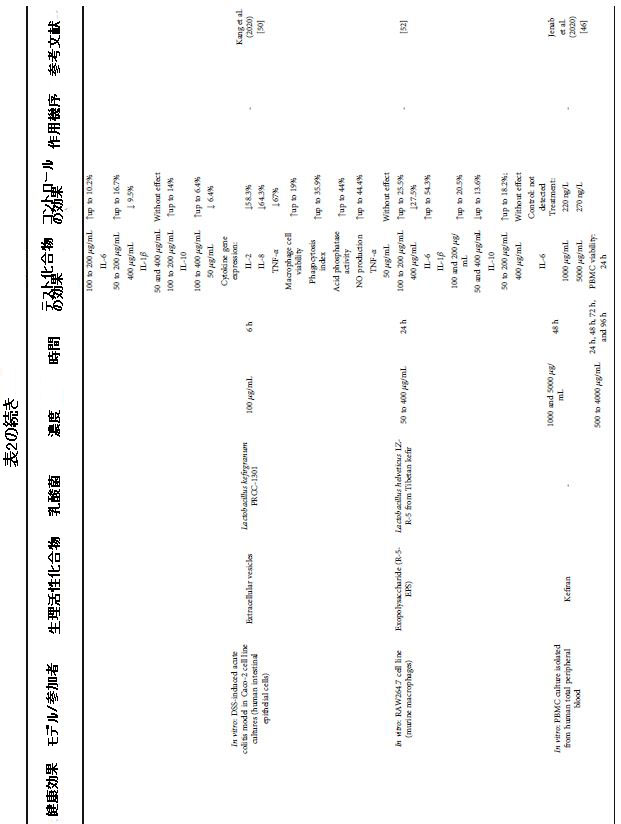 |
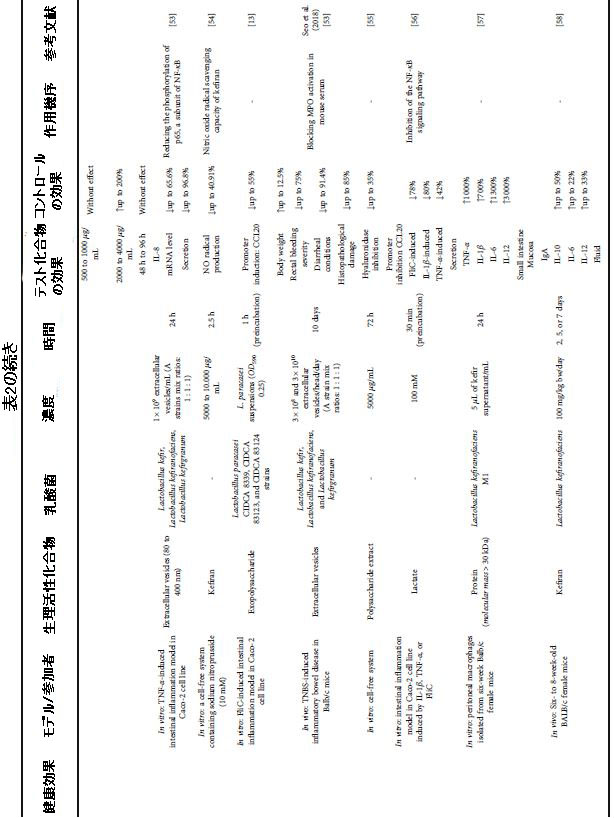 |
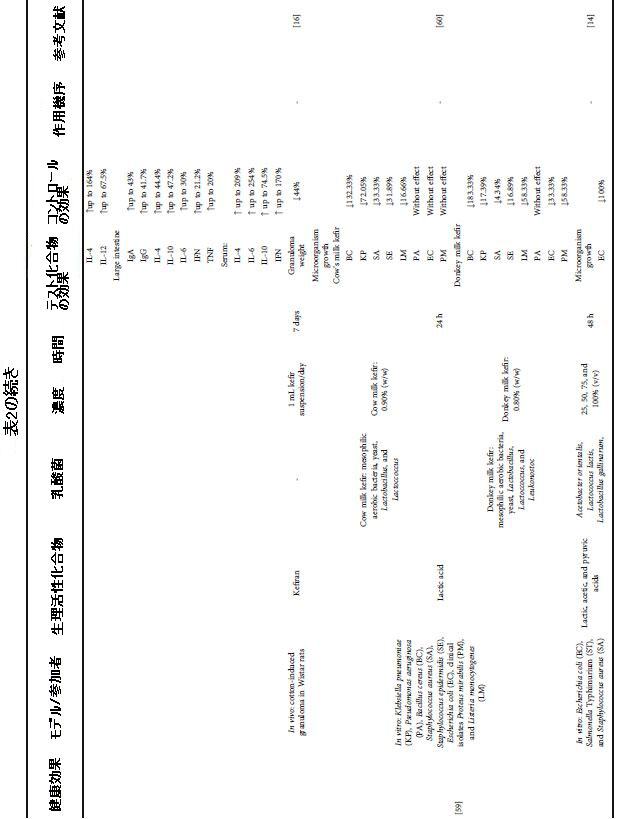 |
 |
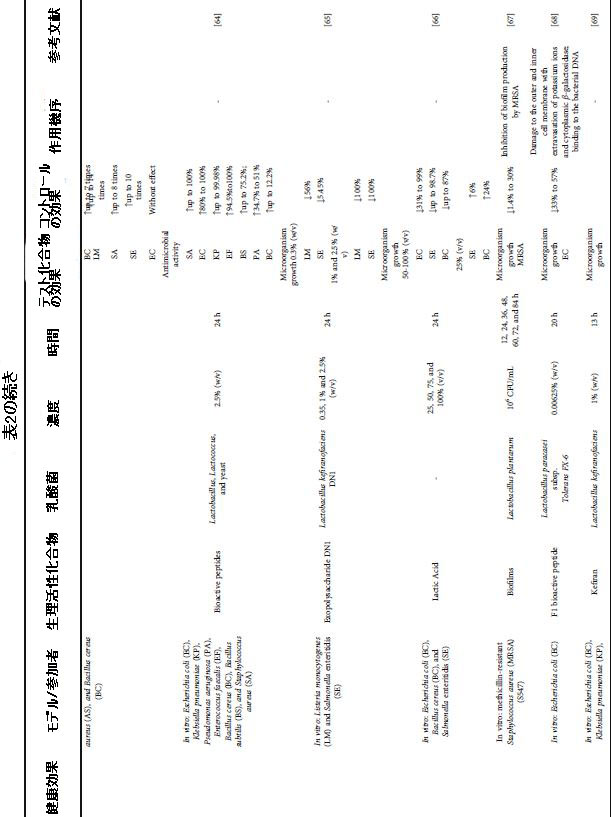 |
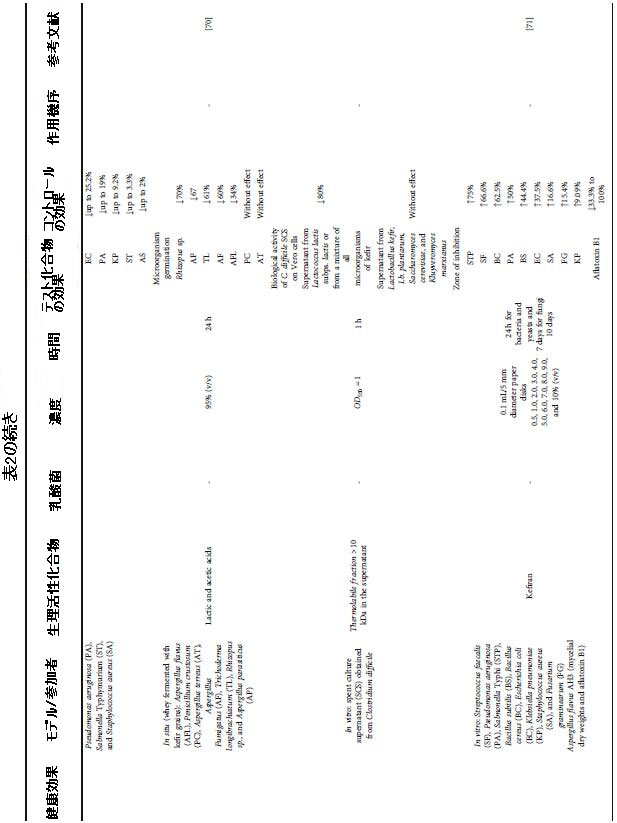 |
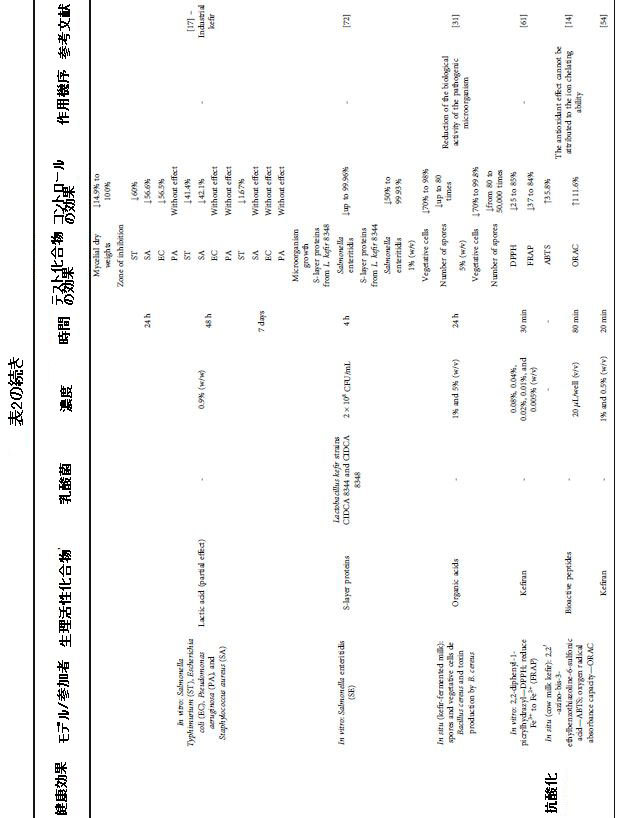 |
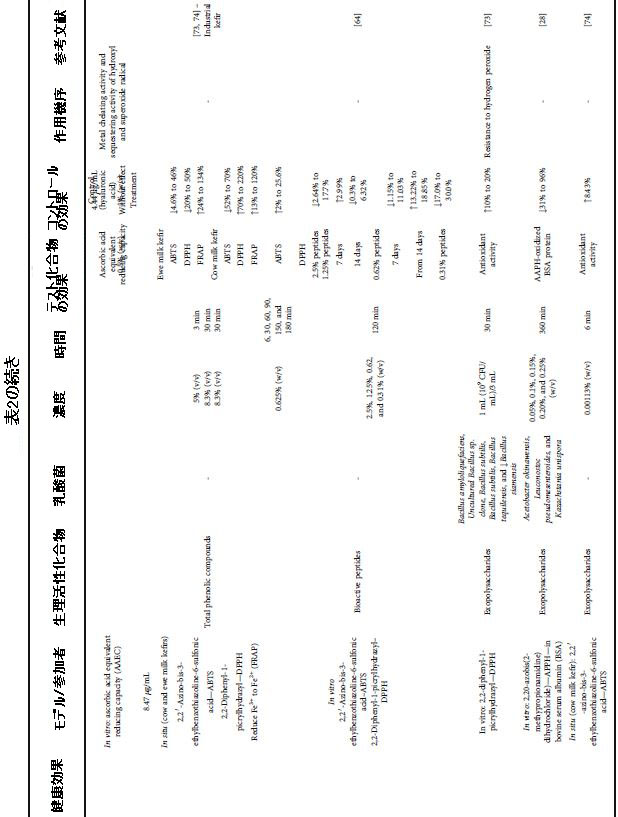 |
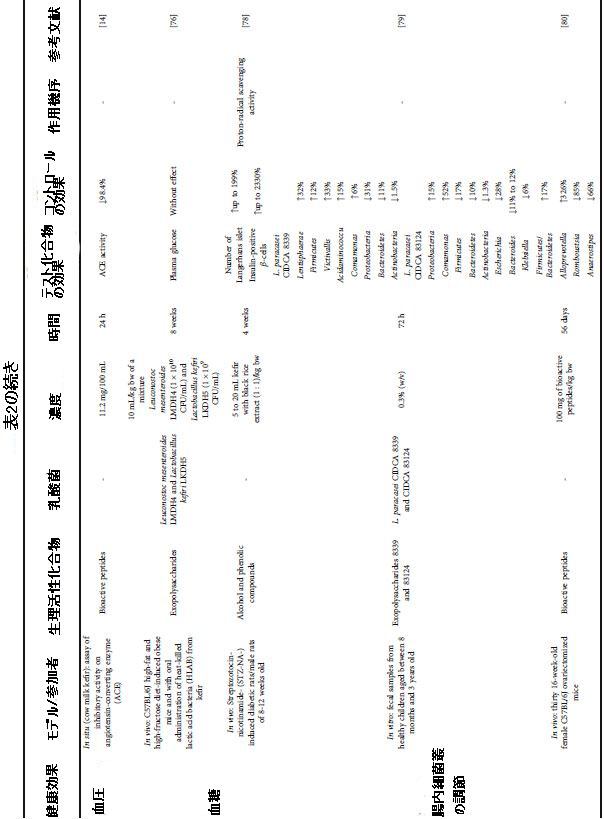 |
 |
 |
| 生産微生物に関しては、ケフィランとEPSはLactobacillus kefiranofaciensによって生産され、生物活性ペプチドはラクトコッカス属と酵母に属する種であるLactobacillus paracaseiによって合成された。バイオフィルムとS層タンパク質は、オンになり、それぞれLactobacillus plantarumとLactobacillus kefirによって生成されました(表2)。これは、ラクトバチルス属の微生物がケフィアで抗菌活性を持つ生物活性化合物の生産に関連していることを示しています。さらに、化合物の抗菌効果に対処する系統的レビューから選択された18の記事のうち、ミルク中の化合物の前駆体を調査したのはそのうちの1つだけでした。したがって、ケフィア中の抗菌化合物の前駆体の調査が現在必要です。Escherichia coli、Klebsiella pneumoniae、Pseudomonas aeruginosa、Enterococcus faecalis、Bacillus cereus、Bacillus subtilis、およびStaphylococcus aureusに対するinvitro抗菌活性は、牛乳中のβ-カゼイン、k-カゼイン、αs1-カゼイン、およびαs2-カゼインから生成された生物活性ペプチドに起因していました [64] |
| ケフィランは、Streptococcus faecalis、Pseudomonas aeruginosa、Salmonella typhi、Bacillus subtilis、Bacillus cereus、Escherichia coli、Klebsiella pneumoniae、Staphylococcus aureus、Streptococcus faecalis、およびFusarium graminearumに対して抗菌効果を示しました。さらに、ケフィランはアフラトキシンを産生する真菌(Aspergillus flavus AH3)にも作用しました。アフラトキシンB1産生の100%から33.3%への減少は、菌糸体の乾燥重量の減少を伴いました[71]。それでも、ケフィランは黄色ブドウ球菌よりも大腸菌に対してより劇的な抗菌効果を示しました。これは、黄色ブドウ球菌の細胞壁にあるペプチドグリカンに起因する可能性があります。次に、ペプチドグリカンは、細胞を介した抗菌剤の拡散を妨げます。さらに、報告によると、シプロフロキサシンとの相乗効果によって示されるように、ケフィランは抗菌効果を得るために必要な抗生物質の濃度を下げることができます[69]。シプロフロキサシンの長期投与は胃と腸の副作用を引き起こすので興味深いです[69]。 |
| それでも、天然の抗菌力としてのケフィランの有望な結果を考えると、ケフィアからの抽出方法も、その抗菌効果を最大化するために研究されてきました。 ケフィラン抽出プロセスで熱水と組み合わせた超音波は、分離された方法によって抽出されたケフィランと比較して、抗菌活性に関して相乗的な結果を示しました[61]。 Lactobacillus kefiranofaciens DN1は、マンノース、アラビノース、グルコース、ガラクトース、ラムノースで構成されるEPSを生成し、0.3%でリステリア菌とサルモネラ菌に対して静菌効果を示しました。 高濃度(1%および2.5%)では、両方の微生物の増殖を完全に阻害することによって殺菌効果が得られ、天然の抗菌剤として使用できる新しい生物活性化合物と見なされています[65]。 |
| 有機酸は、ケフィア微生物によって生成される他の抗菌化合物です。これに関連して、乳酸は牛乳ケフィアとロバ牛乳ケフィアの抗菌効果に関連する生物活性化合物でした。ロバミルクケフィアは、カナマイシン抗生物質と比較して、Klebsiella pneumoniae, Bacillus cereus, および Proteus mirabilisを8%、37%、および58%減少させました。対照的に、牛乳ケフィアはB. cereusを12.9%減少させましたが、カナマイシンと比較してProteus mirabilisに同じ影響を与え、乳酸に関連する驚異的な抗菌力を示しています[60、66]。しかし、緑膿菌に対する抗菌効果はありませんでした[17、60]。酢酸やピルビン酸など、ケフィアに存在する他の有機酸も、乳酸と一緒に投与すると抗菌効果を示しました[14、31、70]。柿須ほか[31]は、有機酸がセレウス菌の胞子の発芽に対して用量依存的に作用することを確認しました。ミルクの発酵に高濃度のケフィア粒(5%)を使用すると、pH値がより大幅かつ迅速に低下し、胞子の発芽が抑制されました。このより重要なpHの低下は、培地中でのより顕著な有機酸生成と関連している可能性があり、その結果、より重要な抗菌効果と関連している可能性があります。さらに、5%のケフィア粒はB. cereusによるNheA毒素の排出量を33.3%から61.8%の範囲で減少させました[31]。 |
| ケフィアの微生物によって産生される生物活性タンパク質であるバクテリオシンは、いくつかの微生物に対して抗菌効果を示しました。 これに関連して、Lactobacillus plantarum、Micrococcus luteus、Listeria monocytogenes、Salmonella enterica serovar Enteritidis、Staphylococcus aureus、およびBacillus cereusは、ネガティブコントロール(滅菌脱イオン水)と比較した場合に効果的でした。 対照的に、大腸菌はバクテリオシンの抗菌作用に耐性があることが証明されました。 ただし、大腸菌は、食品業界で防腐剤として一般的に使用されている天然の抗菌ペプチドであるポジティブコントロール、ナイシン、およびポリリジンにも耐性があることに注意する必要があります。サルモネラ菌については、ナイシンと同様の抗菌効果が観察されました[40]。 |
| ケフィアの一部の微生物は、Clostridium difficile, Pseudomonas putida, Pseudomonas aeruginosaおよびメチシリン耐性Staphylococcus aureusに対して抗菌活性を持つ画分を生成しました[59、62、63]。 25 mg / mLのFK-1000は、緑膿菌の増殖を91%抑制しました。 ただし、低濃度(0.25 mg / mL)の場合でも、FK-1000はストレプトマイシンとの相乗効果を示し、この抗生物質の5倍の結果を増強しました。 これは、併用療法で使用するためのFK-1000の可能性を示しています。 もう1つの重要な側面は、50 mg / mLの濃度のFK-1000がヒト上皮細胞に対して毒性がなく、治療としてのこの化合物の使用の関連性を高めていることです[62]。 |
| さらに、Lactococcus lactis subsp lactis CIDCA8221によって生成された10kDaを超える熱不安定性画分は、C. difficileの毒素産生効果を阻害することが示されました。 この画分は、C. difficilによって引き起こされるアクチンネットワークの中断とベロ細胞の移動を抑制しました。 また、ClostridiumによるTcdAおよびTcdB毒素の形成の減少が観察されています[59]。 |
| 発酵中のケフィアの微生物によるミルクのタンパク質分解は、抗菌活性を持つ生物活性ペプチドをもたらします。 F1生物活性ペプチドは、33%から57%の範囲で大腸菌の増殖を抑制しました。 一貫して、ケフィール由来の生物活性ペプチドの混合物は、Klebsiella pneumoniae, Pseudomonas aeruginosa, Enterococcus faecalis, Bacillus cereus, Bacillus subtilis, および Staphylococcus aureusなどのいくつかの微生物に対して抗菌活性を有していた[64、68]。 |
| ケフィア粒に存在する微生物であるLactobacillus plantarumは、抗菌剤として作用するバイオフィルムを生成し、メチシリン耐性Staphylococcus aureusの増殖を1.4〜30%阻害します。 培地の時間、温度、および通気は、ケフィアの微生物によるバイオフィルムの生成に影響を及ぼしました。これは、細胞の成熟、酵素反応、および特定の遺伝子の活性化発現に関連している可能性があります。 したがって、外的要因および使用される菌株に関連する要因は、Lactobacillus plantarumによるバイオフィルムの生産に関連する抗菌活性に影響を与える可能性があります[67]。 |
| 最後に、病原体に関するケフィアに存在するいくつかの微生物の抑制効果は、それらの細胞膜の外面上のS層タンパク質の存在に関連している可能性があります。 したがって、Lactobacillus kefirとS.enteritidisのプレインキュベーションにより、L.kefirのS層タンパク質を介したそれらの間の直接相互作用が可能になり、病原体の細胞膜上の腸細胞に結合する部位が減少しました。 さらに、Lactobacillus kefir株のS層タンパク質も、Salmonella enteritidisの生存率を低下させました。 興味深いことに、サルモネラ菌と相互作用しない非凝集株から得られたS層タンパク質でさえ、S.enteritidisと相互作用する可能性があります。 S層に存在するコンフォメーションと活性基は、溶液中に分離されたタンパク質と細菌の表面に存在するタンパク質(Lactobacillus kefir)の間で異なる場合があります。 したがって、S層タンパク質は天然の抗菌剤として使用される可能性があります[72]。 |
| 結果は、細菌および真菌に対するケフィア生物活性化合物の統計的に有意な全体的な効果を示しているため、抗菌カテゴリーのメタアナリシスは、ここでの系統的レビューの結果と一致します(補足図S1)。明確な結果の測定値は試験間で異なるため、標準化された平均差(SMD)を使用して効果を推定しましたが、負のSMD値は微生物の減少を示します。 したがって、標準平均差(SMD)は、補足図S1に示されているように全体的な抗菌効果(SMD(-1.35)(95%CI:-1.79–-0.91、P≤.001))と、真菌(SMD(-7.18)(SMD(-7.18))について別々に推定されました。 95%CI:-9.08–-5.28、P≤.001))、Bacillus cereus(SMD(-0.25)(95%CI:-1.06–0.56、P = 0.54))、Escherichia coli(SMD(-0.67)( 95%CI:-2.26–0.92、P = 0.41))、Klebsiella pneumoniae(SMD(-1.79)、(95%CI:-4.22–0.63、P = 0.15))、Pseudomonasspp。 (SMD(-1.98)、(95%CI:-3.48–-0.49、P = 0.009))、Salmonella spp.(SMD(-0.53)、(95%CI:-1.12–0.06、P = 0.08))、およびStaphylococcus spp. (SMD(-0.88)、(95%CI:-1.81–0.06、P = 0.07))。 これに関連して、ケフィアは全体的な抗菌効果に関して統計的有意性を示しましたが、ケフィアからの生物活性化合物はfungi および Pseudomonas spp.に対してのみ有意な効果を示しました。ただし、統計的に有意ではありませんが、調査したすべての微生物に対してケフィア処理を好む傾向が見られます。 |
| データの不均一性を評価するために、9つの分析すべてのI2テストは、fungi (I2 = 83%, P ≤ .001)および全体的な抗菌効果(I2 = 72%、P≤.001)、Bacillus cereus(I2 = 63%、P≤.001)、Escherichia coli(I2 = 67%、P≤.001)、Klebsiella pneumoniaeについて統計的に有意な中程度の不均一性 肺炎菌(I2 = 64%、P = 0.004)、Pseudomonas spp. (I2 = 61%、P = 0.0003)、およびSalmonella spp. (I2 = 68%, P ≤ .00001)に対して統計的に有意なかなりの不均一性を示しました。 |
| 抗菌剤のメカニズムを調査する研究はほとんどありませんが、これらのメカニズムは多様であるように見え、生物活性化合物の種類ごとに特異点を示しています。 生理活性ペプチドの場合、作用は細胞膜と病原体のDNAで起こりますが、有機酸は生物活性を低下させます。 次に、バイオフィルムは病原性微生物によるバイオフィルムの生成を阻害します(図2)。 これに関連して、生物活性化合物は病原体の膜に細孔を形成し、原形質膜の完全性を損ない、その透過性を高め、その結果、カリウムイオンの流出、タンパク質、および核酸の漏出を引き起こします。 これらの化合物は、病原体のゲノムDNAに結合することも説明されています[64、68]。 |
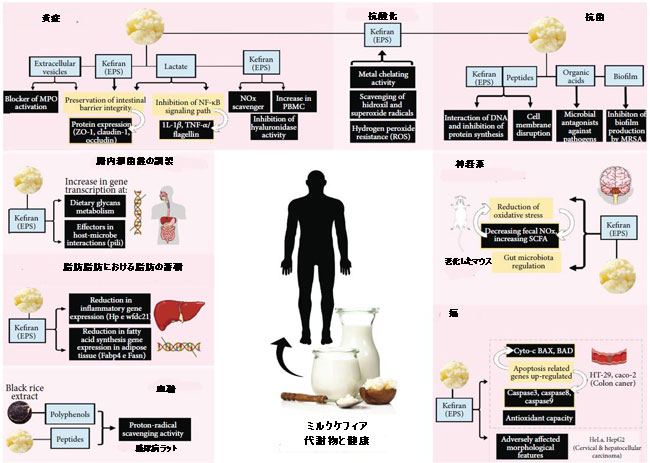 |
| 図2 ミルクケフィアにおける生物活性化合物の作用機序 |
| EPS:エキソポリサッカライド; MPO:ミエロペルオキシダーゼ; ZO-1:閉鎖帯-1; IL:インターロイキン; TNFα:腫瘍壊死因子-α; NO:一酸化窒素; PBMC:末梢血単核細胞; ROS:活性酸素種; MRSA:メチシリン耐性黄色ブドウ球菌; Hp:ハプトグロビン-2; Wfdc21:Wfdc21タンパク質; Fabp4:脂肪酸結合タンパク質4; Fasn:脂肪酸シンターゼ; SCFA:短鎖脂肪酸; Cyto-c:シトクロムc; BAX:BCL2関連Xタンパク質; BAT:細胞死のBCL2関連アゴニスト。 |
| invitroおよびinsitu研究はケフィアからの生物活性化合物の抗菌的役割を示唆しているが、制御されたin vivo研究がないことは、生理学的システムにおけるこの効果の程度の評価を制限する文献のギャップであることを強調することが不可欠です。 |
| 3.4.2 抗酸化活性 |
| 抗酸化分子はフリーラジカルと相互作用し、連鎖化学反応を終わらせ、細胞損傷を引き起こすタンパク質[28]およびDNA [49]への攻撃を減らすため、抗酸化分子による介入は非常に重要です[84]。 |
| ケフィアからの抗酸化活性で同定された主な生物活性化合物は、ケフィランを含むEPSでしたが、生物活性ペプチドとフェノール化合物も記載されていました(表2)。 さらに、抗酸化特性を持つ化合物の生産微生物を特定する研究はほとんどありません。 しかし、Acetobacter, Leuconostoc, Bacillus, および Kazachstaniaの菌株が報告されました(表2)。 ミルクの前駆物質に関しては、彼らの調査は明らかに不足していました。 生物活性ペプチドの前駆体がβ-カゼイン、k-カゼイン、αs1-カゼイン、およびαs2-カゼインであると特定した研究は1つだけでした[64]。 |
| アスコルビン酸当量容量(AAEC)の還元活性を低下させることによって測定した場合、invitroでのケフィランの抗酸化活性はそれぞれ1%および0.5%の濃度で8.47μg/ mLおよび4.44μg/ mLでした。 興味深いことに、ゴールドスタンダードのビスコサプリメント処理で使用されたヒアルロン酸は、ケフィランに使用されたものと同様の濃度で還元力を示しませんでした。 これは、ケフィアから単離された化合物の抗酸化能を示しています[54]。 フリーラジカルの除去と第二鉄イオンの還元は、ビタミンC活性のそれぞれ25%から85%と37%から84%を表しており、これらの活性のポジティブコントロールと考えられています。 ケフィランのテストされた比較的低い濃度(0.005%から0.08%)は、ビタミンCと比較してその劣った効果を正当化することができます[61]。 |
| ケフィア内の微生物によって生成されたEPSの抗毒素活性は、酸化からの生体分子の保護を促進しました。インラインでは、低濃度のEPS(0.05%〜0.25%)は、APPH(2,20-アゾビス(2-メチルプロピオンアミジン)二塩酸塩)によって誘発される酸化からBSAタンパク質を保護する可能性があります。タンパク質の酸化減少は31%から96%の範囲でした。興味深いことに、この保護は、ネガティブコントロール(酸化を誘発しないタンパク質)で観察されたものよりも高かった[28]。さらに、EPSに関連するinsituでの抗酸化能は、24時間の発酵後に8.43%高かった[74]。 DPPH(1,1-ジフェニル-2-ピクリルヒドラジル)フリーラジカルを除去する能力は、10%から20%に増加しました。この事実は、発酵中のケフィア微生物の代謝活性に起因する可能性があり、EPSの生成と蓄積につながります。したがって、研究により、ケフィアに存在するEPSは、抗酸化剤としての可能性を示していることがわかりました[73]。 EPSはまた、過酸化水素に対する耐性を付与し、ケフィアに存在する微生物の細胞増殖に対するこの化合物の有害な影響を完全に逆転させます。一貫して、EPSのない細胞は過酸化水素に対する耐性を大幅に失います[73]。 |
| 生物活性ペプチドなどのケフィアに存在するより小さな画分も、抗酸化効果を示しました。 酸化2,2'-ジノビス(3-エチルベンゾチアゾリン-6-スルホン酸)(ABTS)の除去能力の向上は、対照と比較して、羊乳ケフィアで2%から25.6%、牛乳ケフィアで35.8%の範囲でした。 [14、64]。 牛乳ケフィアの酸素ラジカル吸収能(ORAC)の大幅な改善(111.6%)により、生物活性ペプチドの抗酸化活性が確認されました[14]。 羊乳ケフィアと牛乳ケフィアの抗酸化活性の間に観察された違いは、特にタンパク質含有量と生物活性ペプチドの前駆体であるそのデザインについて、牛乳間の組成の違いに関連している可能性があると考えています。 この発見は、ミルク中の生物活性化合物の前駆体を特定して、その機能的可能性を高めることの関連性を浮き彫りにしています。 |
| ケフィア粒によるミルクの発酵プロセスは、フェノール化合物を合成することによってその抗酸化活性を高めることを示しました。職人技のケフィアでは、抗酸化能はABTSで測定すると最大120%、ORACで最大40%増加しました[75]。対照的に、市販のケフィアの場合、ABTSによる抗酸化能力は低下しましたが、FRAP(鉄の抗酸化力)による抗酸化能力は増加しました[85]。したがって、スターターカルチャーによって生成されたフェノール化合物は、それらの還元力によって抗酸化作用を発揮するように見えますが、酸化された化合物の除去能力に関しては効果がありません。それでも、DPPHスカベンジングはケフィアの種類によって異なり、対照と比較して雌羊のケフィアが減少し、牛のケフィアが増加しました[85]。したがって、マトリックスによれば、フェノール化合物のプロファイルは、その場でのラジカル捕捉能力に影響を与える可能性があります。実際、SatirとGuzel-Seydim [75]は、没食子酸、カテキン、エピカテキン、カフェー酸、p-クマル酸、クロロゲン酸、フェルラ酸、および光カテキン酸の存在下で抗酸化活性の増加を観察しました。ケフィア微生物の代謝活性から生じるフェノール化合物は、発酵中のその場での抗酸化能力の最大120%を改善することができます[85]。 |
| 介入時間は、同様の方法論のEPSおよびフェノール化合物よりも生物活性ペプチドの方が顕著でした(表2)。 この発見は、ペプチドが有意な抗酸化活性を発揮するのにより長い時間を必要とすることを示唆している。 一方、フェノール化合物は、他の生物活性化合物の最大3倍の濃度を必要としました(表2)。 |
| 記載された発見と一致して、メタアナリシスはケフィア生物活性化合物の抗酸化傾向を示しました。 ただし、信頼区間が狭いことを観察したにもかかわらず、対照治療と比較して統計的に有意であるとは見なされませんでした。 その結果、(SMD(-0.83)(95%CI:-1.65–0.00、P = 0.05))の標準平均差(SMD)が得られました(補足図S2)。 さらに、研究の結果は、有意な実質的な不均一性(I2 = 84%、P≤.001)を示しました。これは、有意性の欠如に寄与した可能性があります。 |
| 文献には作用機序の記述が不足しているが、EPSは金属キレート活性とヒドロキシルラジカルおよびスーパーオキシドラジカルの隔離活性を介して効果を発揮し、その結果過酸化水素に耐性を示すようである。 対照的に、生物活性ペプチドの抗酸化活性は、イオンキレート能力に起因するものではありません[14]。 |
| 抗菌効果について以前に報告されたように、ケフィアからの生物活性化合物の抗酸化能を評価する制御されたvivo実験の欠如は、生理学的システムにおけるこの効果の程度の評価を制限する文献のギャップです。 |
| 3.4.3 腸内細菌叢の調節 |
| 腸内細菌叢を宿主と共生させることは、腸内バリアの完全性、免疫系のバランス、および炎症過程の制御に有利に働くため、人間の健康にとって不可欠です。 したがって、腸内細菌叢の調節に有利なプロバイオティクスまたは生物活性化合物の検索は、最近の文献で広く研究されています。 |
| ケフィランを含むエキソポリサッカライド(EPS)は、invitroおよびinvivoモデルで腸内細菌叢を調節することができる主に記載された生物活性化合物でした(表2)。 ただし、これらの生物活性化合物の前駆体を特定する研究は不足しています。 さらに、ミルクでそれらを生成する微生物を決定する研究の不足は、文献のギャップを浮き彫りにします。 興味深いことに、生物活性ペプチドを使用した濃度と介入時間は、invivo動物モデルのEPSを使用した場合よりもかなり長かった。 ペプチドの使用濃度は最大2.5倍でした。 介入時間は、EPSの使用と比較して最大2.6倍長かった(表2)。 これは、EPSが腸の調節に対してより強力な生物活性化合物である可能性があることを示唆しています。 ただし、この前提はさらに調査する必要があります。 |
| L. paracasei CIDCA8339およびCIDCA83124によってケフィアで生成されたEPSは、乳児の糞便サンプルに存在する微生物叢を変更し、その結果、短鎖脂肪酸プロファイル(SCFA)を変更することを示しました。 生成される酪酸とプロピオン酸は、健康上の利点に関連する生物学的活性を持つ化合物です。 これらの利点の中で際立っているのは次のとおりです:腸上皮バリアを強化し、肝臓でのコレステロール合成を阻害します。 それでも、レプチン、YYポリペプチド(PYY)、およびそれらによって促進されるグルカゴン様ペプチド1(GLP-1)の発現は、脂肪組織の脂質生成と食欲を調節します[79]。 |
| 門のレベルを考慮すると、表2に示すように、EPSと生物活性ペプチドの使用により、ファーミキューテス門は一般にバクテロイデス門に悪影響を及ぼしました。属レベルに関しては、KlebsiellaやEscherichiaなどの病原性に関連する微生物の減少が報告されています。 健康な腸内細菌叢に寄与するEPSの可能性を示しています[79]。 Rikenellaceaeの付随する減少は、結腸上皮組織の健康の促進剤でもあります[81]。 対照的に、EPSはVictivallis、Acidaminococcus、Comamonas、およびRuminococcaceae科を支持しました[79、81]。 Acidaminococcus属の特定の種を選択することは、腸のレベルで有益であるプロピオン酸や酪酸などの有機酸の増加に関与している可能性があります。 |
| 一方、ラクトバチルス菌やビフィズス菌など、健康に有益であると考えられている他の属の調節は、介入中に使用された生物活性化合物に依存していました(表2)。したがって、LactobacillusとBifidobacteriumは、Bengoa et al [79]のEPS8339と83124によって支持されませんでした。対照的に、Xing et al [81]の研究では、成体のオスのマウスを使用して、Lactobacillus kefiranofaciens XL10によって生成されたEPSは、乳酸桿菌で14.59%、ビフィドバクテリア科で最大0.59%の成長を示しました。ケフィランを使用した場合、成体雌マウスのビフィズス菌で最大17%の増殖も観察されましたが、ラクトバチルスの個体数に変化は観察されませんでした[82]。In vitroでは、0.3%のケフィランがBifidobacterium bifidumPRL2010の個体数を5.8×108CFU / mLまで上昇させました。対照的に、対照(炭素源のない培地)ではビフィズス菌の個体数の増加はありませんでした。この事実は、各介入における腸の調節に最も適切な生物活性化合物を決定する際に、標的微生物を考慮すべきであることを示しています。 |
| 興味深いことに、EPSは、食品業界で一般的に使用されているプレバイオティクスであるイヌリンよりも、腸内細菌叢の多様性を促進するのに強力でした。 それでも、EPSは異なる短鎖脂肪酸プロファイルをもたらし、酪酸含有量を増加させ、イヌリンよりも腸内微生物集団に利益をもたらしました[79]。 ケフィランはまた、Bifidobacterium bifidum PRL2010の増殖において、グルコースよりも優れた炭素源であることが証明されました。 ケフィランは、グルコースの使用と比較して、菌株の発生を20〜700%増加させました[41]。 |
| ケフィアに存在する他の生物活性化合物である生物活性ペプチドも、腸内細菌叢の変化に関連していた。雌マウスへのペプチドの経口投与は、卵巣摘出術によって引き起こされた腸内細菌叢への有害な影響を部分的に逆転させました。ケフィアからのペプチドは、Parasutterellaの46%とStreptococcusの39%を減らしました。クレブシエラ属とエシェリキア属に属する潜在的に病原性の細菌も減少しました。 Romboutsiaは、Streptococcusとともに肥満に関連しており、86%の減少を示しました(表2)。ケフィア由来の生理活性ペプチドは、卵巣摘出術によって誘発される腎脂肪の蓄積を減らすことにより、Alloprevotellaの個体数を30%以上増加させました。Ruminococcus-1、SCFA産生菌は、卵巣切除マウスで増加し、その後、生物活性ペプチドの投与により減少しました。卵巣切除マウスでのエストロゲン欠乏後の酪酸産生細菌の増殖は、腸内でのSCFAの有害な蓄積に悪影響を与える可能性があることを強調することが不可欠です。対照的に、生物活性ペプチドは、卵巣摘出術によって引き起こされたデフェリバクター門の減少を回復することができませんでした。Deferribacteresの減少は、ビタミンとアミノ酸代謝の有害性に関連しています[80]。 |
| したがって、生物活性ペプチドは、卵巣切除された女性の腸内の有益な細菌を濃縮し、潜在的に有害な病原体を減少させることができます[80]。 それでも、これらの発見は、腸内細菌叢とエストロゲン欠乏症との関係、およびこの関係におけるケフィアからの生物活性ペプチドの役割を分析するさらなる研究が、より良い理解のために必要であることを明らかにしています。 |
| 微生物叢の豊富さと多様性に関しては、興味深いことに、卵巣切除されたマウスの生物活性ペプチドの影響を受けません。 しかし、ペプチドは、卵巣切除術を行わない対照と比較して、腸内細菌叢の多様性を大幅に改善します[80]。 |
| メタアナリシスに関しては、生物活性化合物による治療は、対照治療と比較して腸内細菌叢の調節に統計的に有意な影響を示しませんでした(SMD(-0.39)(95%CI:-1.32–0.54、P = 0.41))(補足図S3)。 しかし、我々の発見と一致して、生物活性の有効性を支持する傾向に気付くことができます。 この分析は、統計的に有意な中程度の不均一性(I2 = 58%、P <.001)を示しました。これは、少なくとも部分的に有意性の欠如に寄与している可能性があります。 |
| 最後に、ほとんどの研究は、腸内細菌叢の調節に対する生物活性化合物の作用機序を評価できず、文献のギャップを表しています。 ただし、微生物とホストの相互作用に関与する遺伝子の転写の強化は、ケフィランの作用のために提案されました。 |
| 3.4.4 免疫応答調整 |
| ケフィアからの生物活性化合物は、モデルの炎症性傷害の有無に応じて、抗炎症性または炎症誘発性の影響を及ぼしました。 したがって、それらは炎症性疾患のモデルにおいて抑制効果を発揮したが、炎症性傷害のないモデルに対しては免疫刺激効果を示した(表2)。 |
| 主に研究された炎症性疾患モデルは、invitroおよびinvivoの両方での急性大腸炎のモデルでした。大腸炎、主にEPSとアポトーシス小胞、さらに乳酸塩については、DSS(デキストラン硫酸ナトリウム)、TNFα、FliC(フラゲリン)、IL-1βおよびTNBS(2,4,6-トリニトロベンゼンスルホン酸)などのさまざまな急性炎症性発作に対して抗炎症作用があると報告されています。。 慢性大腸炎の場合、細胞外小胞はピロキシカムに対して抗炎症効果を示しました。次に、ケフィランは、ラットのワタ誘発肉芽腫に対して抑制効果を示しました(表2)。 L. kefirgranum、L.kefir、L.kefiranofaciens、およびL. paracasei種は、これらの生物活性化合物の生成に関与していました。したがって、ラクトバチルス属はケフィアで抗炎症化合物を作るのに関連しているようです[13、50、53]。ガラクトースとグルコース、そして程度は少ないがマンノース、アラビノース、ラムノースは、生物活性化合物の多糖類成分の重要な前駆体でした[55]。 |
| L.kefirgranumからの100μg/ mLの細胞外小胞は、DSS誘発性急性大腸炎モデルのCaco-2細胞におけるIL-2、IL-8、およびTNFα炎症性サイトカインの遺伝子発現をそれぞれ58、64、および67%減少させました [50]。 細胞外小胞(1×109粒子/ mL)もTNFα誘発性大腸炎を抑制しました。それらは、Caco-2細胞株においてIL-8の発現と分泌をそれぞれ65%と96%減少させました。興味深いことに、細胞外小胞は、クローン病(炎症性腸疾患)の治療に一般的に使用される糖質コルチコイドステロイドであるブデソニド[53]と同じくらい効果的でした[86]。それでも、細胞外小胞によるCaco-2細胞の治療は、EPSまたは乳酸による治療よりも長い介入時間を示しました。さらに、EPSまたは乳酸塩との細胞のプレインキュベーションで観察された効果(表2)は、腸の炎症の予防剤としてのこれらの化合物の可能性を示しています。ただし、これらの生物活性剤の予防的および治療的可能性を解明するために、炎症性傷害に関するインキュベーション前およびインキュベーション後を調査するさらなる研究を実施する必要があります。 |
| マウスでは、細胞外小胞の経口投与は急性および慢性大腸炎を軽減する可能性があり、invitroでの以前の発見を裏付けています。 DSS誘発性急性大腸炎の場合、小胞の高用量と低用量の両方で、マウスの体重減少が最大16%防止され、結腸組織への損傷が最大63%減少しました。 しかし、最高用量(3 mg / kg体重)のみが結腸萎縮を29.6%減少させました。 同様に、ピロキシカムによって悪化した慢性大腸炎では、最高用量のみが結腸萎縮を14.3%軽減しました。 それにもかかわらず、両方の投与量は結腸の組織学的損傷を最大85%減少させました。 |
| 急性大腸炎とは対照的に、細胞外小胞の摂取は慢性大腸炎の体重減少を防ぐのに効果がありませんでした[50]。したがって、3 mg / kg体重で得られたより広範な効果により、高用量は慢性および急性大腸炎の治療により効果的であるように思われた。 TNBS誘発性急性大腸炎に対する小胞の投与はまた、3×108および3×1010小胞/頭でマウスの体重減少を最大12.5%まで効果的に防止しました。さらに、投与により、直腸出血および下痢状態の重症度がそれぞれ75%および91%減少しました。次に、結腸組織への損傷は最大85%減少しました[53]。ケフィアのラクトバチルスからの小胞による治療は、体重減少、直腸出血の重症度、および下痢状態の予防、ならびに結腸の組織学的損傷の軽減において、プレドニゾロン薬(2 mg / kg)よりも効果的でした[53]。プレドニゾロンは、大腸炎やクローン病の炎症を治療するために使用される抗炎症ステロイドです。しかし、いくつかの副作用があることに加えて、病気の再発を防ぐことはできません[87、88]。したがって、ケフィアからの生物活性化合物による治療は、有効性と副作用の低減の両方に有望です。 |
| EPSプロデューサーLactobacillusparacasei(OD590 = 0.25)の懸濁液は、フラジェリン誘発性急性大腸炎モデルのCaco-2細胞におけるCCL20炎症性プロモーターの誘導を最大55%阻害しました。 Lactobacillus paracasei CIDCA 8339菌株は、他の試験菌株よりも劇的な抗炎症能を示しました[13]。これは、EPSの機能的ポテンシャルが菌株に依存していることを示しています。同様に、100 mMの乳酸は、Caco-2細胞において、フラジェリン、IL-1β、およびTNFαによるCCL20プロモーター誘導をそれぞれ78、80、および42%阻害しました。インラインで、ヒト腸上皮細胞は乳酸受容体を発現します。それでも、対応する乳酸濃度のケフィアからの乳酸溶液および上清は、Caco-2細胞に対して同様の阻害効果を示し[56]、ケフィアマトリックスがこの生物活性化合物の影響を低減しないことを示しています。In vivoでは、ケフィランに富むケフィア上清の経口投与(1 mL /日)が、ラットの綿誘発性腹部肉芽腫の重量を44%減少させる原因となった。ケフィアはこれらの肉芽腫の減少にデキサメタゾン(0.2 mg / kg)と同じくらい効果的でした[16]。デキサメタゾンは、肉芽腫の治療における主要な選択肢として使用されるコルチコステロイド薬です[89]。この証拠は、ケフィアからの生物活性化合物の抗炎症の可能性を強化します。 |
| したがって、一般に、生物活性剤は、炎症性サイトカインの発現およびCaco-2細胞を用いたinvitro炎症モデルのCCL20プロモーターの活性化を阻害した。 in vivo大腸炎モデルでは、生物活性化合物は、体重減少、萎縮、および結腸の組織学的損傷を軽減した。 |
| これらの化合物は、IκBα阻害剤の発現により、Caco-2細胞および結腸粘膜のNF-κB経路を阻害することにより、抗炎症作用メカニズムを示しました[50、53、56]。 さらに、生物活性剤は腸のバリアの完全性を促進し、オクルディン、ZO-1、およびクローディン-1閉塞タンパク質の発現を増加させました[50]。 EPSに対して提案された追加の抗炎症メカニズムは、一酸化窒素ラジカル捕捉能力[54]および無細胞invitroシステムにおけるヒアルロニダーゼ活性の阻害[55]でした。ヒアルロニダーゼによる細胞外マトリックスの加水分解は、炎症性病変全体にヒアルロナンのような化合物を放出します[90]。細胞外小胞については、マウス血漿中のミエロペルオキシダーゼ活性化の遮断も報告されています[53]。炎症性腸疾患における酸化ストレスは、好中球などの炎症性細胞を活性化し、そのミエロペルオキシダーゼは活性酸素種の産生を触媒します[91]。このシナリオでは、細胞外小胞はミエロペルオキシダーゼの阻害においてプレドニゾロン薬と同じくらい効果的でした[53]。したがって、証拠は、ケフィアからの生物活性化合物が決定的な抗炎症の役割を果たす可能性があることを示唆しています。 |
| しかし、ケフィランを含むEPSは、炎症性発作がない場合、免疫賦活剤として作用する逆の効果も持つ可能性があります(表2)。 この役割は、invivoモデルでも裏付けられました[58]。 ミルク中のそれらの前駆体はブドウ糖とガラクトースでした[51、52]。 少数派の方法で、生物活性ペプチドも免疫賦活剤として報告されています[57]。 Lactobacillus属は、これらの免疫賦活剤、特にL. helveticus、L。pentosus、およびL.kefiranofaciens種の産生に関連していました。 |
| EPSの場合、介入時間は炎症誘発性および抗炎症性効果アッセイの両方で同様でしたが、採用されたEPS濃度は劇的に高かった。 in vitroでは、免疫刺激の濃度は50から5000μg/ mLまで変化しましたが[4、46、51]、阻害の場合は最大100μg/ mLに達しました[50]。 In vivoでは、免疫刺激のために100 mg / kg体重が経口投与されたが[58]、免疫応答を阻害するために、濃度は0.03から3 mg / kg体重の範囲であった[50]。 この事実は、EPSの濃度が免疫系で果たす役割を決定する重要な要因であることを示唆しています。 並んで、EPSは、試験した濃度に応じて、in vitroのマウスマクロファージによるTNFα、IL-10、およびIL-6の分泌を刺激または阻害することが報告されています[51、52]。 したがって、EPSは、その濃度に応じて、さまざまな細胞シグナル伝達経路によって作用する可能性があるようです。 ただし、作用の免疫刺激メカニズムはまだ研究する必要があります。 |
| In vitroでは、EPSおよび生物活性ペプチドの免疫刺激の役割は、ヒト末梢血単核細胞(PBMC)に加えて、マクロファージの細胞株および初代培養で実証されています(表2)。 50-400μg/ mLのR-5-EPSおよびR-17-EPSは、増殖、食作用、ホスファターゼ活性、IL-6分泌、およびRAW264.7マウスマクロファージ細胞によるNO産生を刺激しました。それでも、100〜200μg / mLの濃度範囲で、TNFα、IL-1β、およびIL-10の分泌を刺激しました。 EPSは、マクロファージによる細胞増殖、食作用、およびサイトカイン分泌の促進において、リポ多糖と同じくらい効率的でした[51、52]。 1000〜5000μg / mLのケフィランはIL-6分泌を増加させ、2000〜4000μg / mLの濃度は24時間後にヒトPBMC培養物の細胞増殖を最大200%刺激しました[46]。 L. kefiranofaciens由来の生物活性ペプチドは、マウス腹腔マクロファージ培養により、TNFα、IL-1β、IL-6、およびIL-12の分泌を1000、700、1300、および3000%改善しました。しかし、異なる微生物株からのペプチドは免疫刺激能力の違いを示し[57]、ペプチドの機能的可能性が株に依存していることを示唆しています。さらに、生物活性ペプチドはTLR2受容体を介して作用しました[57]。トール様受容体2は、マクロファージが微生物のリガンドを認識できるようにし、それによって炎症を促進します[92]。 |
| 一貫して、ケフィラン(100 mg / kg体重)を健康なマウスに最大7日間経口投与すると、小腸の粘膜のIgA、IL-10、IL-6、IL-12、およびIL-4が増強されました。 小腸の液体中のIL-12。 血清中、ケフィランはIL-4、IL-6、IL-10、およびIFNを増加させました。 しかし、より広範な免疫刺激が大腸で起こり、IgA、IgG、IL-4、IL-10、IL-6、INF、およびTNFの含有量が増加しました。 大腸で最も明白な刺激活性は、腸内細菌叢によるケフィラン発酵に起因しています[58]。 したがって、生物活性は、これらの分子が消化過程全体で受ける生化学的変換に応じて変化する可能性があるようです。 |
| 最後に、免疫刺激が感染症のより良い予後[93]およびワクチン接種後の免疫グロブリン産生の刺激[94]にとって興味深い可能性があることを強調することが不可欠です。 したがって、濃度および環境状況(炎症性発作の有無)は、生物活性化合物を投与する目的に応じて考慮すべき関連要因です。 |
| メタアナリシスの結果は、治療が対照と比較して有意な免疫調節活性を有することを示したため、免疫調節に対するケフィア生物活性化合物の利点を裏付けました(補足図S4)。 推定(SMD)は(SMD(-1.17)(95%CI:-1.47–-0.87、およびP≤.001))でした。 この分析では、不均一性は統計的に有意に中程度でした(I2 = 67%、P≤.001)。 |
| 3.4.5 抗がん効果 |
| 癌は、転移する可能性のある典型的な無秩序な細胞増殖に見られる100種類以上の悪性腫瘍を含む用語です[95]。 ただし、ヒトのin vitroモデルでは、ケフィア由来の生物活性化合物の乳がん、結腸がん、子宮頸がん、および肝細胞がんに対する抗がん効果が報告されていますが、in vivo研究がないため、生物におけるそれらの抗がん効果の程度の理解が制限されています。 生物活性のあるものの中で、ケフィランを含むEPSは、乳がん、結腸がん、子宮頸がん、および肝細胞がんなどのさまざまな種類のがんをカバーする、より広範な影響を及ぼしました(表2)。 ミルク中のEPSの主な前駆体はブドウ糖とガラクトースでした[47]。 ラクトバチルス属は、ケフィアに対する抗癌効果を伴うEPS産生に関連しているようでした(表2)。 |
| ケフィランは、介入の48時間後にMCF7乳がん細胞の生存率を最大45%低下させ、4 mg / mLの最高濃度で効果を失いました。ただし、効率は72時間後に15.6%に低下し、テストされた最低濃度の0.5 mg / mLでは効果がありませんでした[46]。 HepG2肝細胞癌細胞の場合、ケフィランは最小濃度の250μg/ mLから最大82%まで生存率を低下させました。対照的に、HeLa子宮頸がん細胞の場合、ケフィランは用量依存的に生存率を最大72%低下させました[48]。同様に、EPS MRS101は、HT-29結腸癌細胞の生存率に用量依存的な効果を示し、24時間後に生存細胞数を最大55.9%減少させました[47]。したがって、EPS濃度と介入時間は、抗発癌効果を得るかどうかを決定するように思われます。ただし、ゼブラフィッシュの胚では、ケフィラン濃度が100μg/ mLを超えると発生毒性が観察され、1000μg/ mLのケフィランが80%の死亡率に達することに注意する必要があります[48]。インラインでは、10-3から3μg/ mLのEPSは、典型的なベロ細胞培養に対して毒性を示しませんでした[55]。したがって、EPSは使用する濃度によっては正常組織の成長に影響を与える可能性があるため、この要因は将来の臨床応用で考慮する必要があります。 |
| EPSに対して提案された作用機序は、アポトーシス遺伝子Cyto-c(シトクロムc)、BAD(BCL2関連細胞死アゴニスト)、BAX(BCL2関連Xタンパク質)、およびカスパーゼ3、8、および9のアップレギュレーションでした。 ; これらの遺伝子の発現の増加は15から120%の範囲でした。 対照的に、EPSはミトコンドリアの酸化安定性に関与するBCL2遺伝子を70%ダウンレギュレーションしました[47]。 遺伝子発現のそのような変化は、癌細胞の形態への悪影響に関連している可能性があります。 付着能力の喪失と細胞内液胞の形成は、細胞死をもたらすより一般的な変化でした[48]。 |
| 一方、生物活性ペプチドは、エストロゲン感受性乳がんモデルでのみ報告されています。さらに、それらは記載されている他の生物活性化合物よりも長い介入時間を示しました(表2)。ただし、ケフィア由来のペプチドは、0.31〜10%(v / v)の用量で正常なヒト乳腺上皮細胞に対して抗増殖効果を発揮しなかったため、ペプチドはEPSに対する利点として毒性が低いようです。一方、これらの正確な投与量では、ペプチドは用量依存的にMCF7-E3ヒト乳がんエストロゲン感受性細胞数を最大88%減少させました[15]。これはこの生物活性化合物の特異性を示しています。それでも、ヨーグルト抽出物は、5%(v / v)濃度から正常なヒト上皮細胞に毒性を示すことに加えて、ケフィア抽出物よりも劇的に低い効率を示しました。非発酵乳は、とは異なり、MCF7-E3癌細胞の増殖を刺激する悪影響を及ぼしました[15]。これらの発見は、ケフィアからのペプチドの抗発癌性特異性を裏付けています。 |
| 有機酸の抗癌効果は直接テストされていませんが、ケフィアに存在する乳酸と酢酸は、HT-29細胞の糞便水誘発性DNA損傷を減少させました。 糞便水で30分間培養すると、HT-29細胞のDNA損傷が36%増加しました。これは、過酸化水素による影響よりも毒性が強い効果です[49]。 しかし、糞便の水によるDNA損傷は、ケフィアの上清で20%抑制されました。 したがって、有機酸は、それらの抗酸化活性に起因して、結腸癌に対する予防効果を有する可能性があります。 インラインでは、ケフィアは未発酵のミルクよりも78.6%高い抗酸化能力(トロロックス等価)を示し[49]、DNAの保護における発酵中に生成された有機酸の役割を裏付けています。 |
| 最後に、HT-29結腸がん細胞株[47、49]は、MCF7乳がん細胞株[46]よりもEPSに対して感受性が高いようでした。 後者の場合、アッセイで報告された濃度は10倍高かった(表2)。 したがって、ケフィアからの生物活性化合物に対する感受性は、細胞型に依存しているように見えます。 |
| 私たちの発見を検証すると、全体的な抗癌効果の推定(SMD)は(SMD(-2.44)(95%CI:-3.41–-1.47、P≤.001))であったため、生物活性化合物による治療は有意な抗発癌効果を示しました。 図S5)。 データは均一であり(I2 = 46%、P≤0.0006)、これは研究間の一貫性が良好であることを意味します。 |
| 3.4.6 血漿グルコース |
| 糖尿病は、網膜症、腎症、脳卒中などの重篤な臨床的影響を伴う慢性的な糖代謝障害です[96]。 2型糖尿病は、膵臓のβ細胞によるインスリン分泌の低下とインスリン抵抗性による血糖値の上昇を特徴とする代謝障害です。 さらに、ここ数十年の世界的な糖尿病の有病率の増加は、人口における肥満の有病率の増加と関連しています[96]。 |
| このシナリオでは、ケフィアの生物活性化合物を、invivoでの肥満および糖尿病マウスモデルのグルコース代謝についてテストしました(表2)。 EPSプロデューサーのLeuconostoc mesenteroidesLMDH4とLactobacillus kefiri LKDH5の混合物を8週間経口投与(10 mL / kg体重)しても、食餌誘発性肥満マウスの血糖値には影響しませんでした。 しかし、この治療は、脂肪細胞における炎症誘発性および脂肪酸合成遺伝子の発現をダウンレギュレートすることに加えて、脂肪細胞組織の重量を効果的に36%減少させました[76]。 この事実は、脂肪組織における脂肪酸代謝の調節が、血漿グルコースの有意な変化を促進する決定的な要因ではないことを示しています。 |
| 対照的に、5〜20 mLのケフィアと黒米抽出物の混合物(1:1 / kg体重)を4週間投与すると、膵臓のランゲルハンス島の最大199%および2330%とインスリン陽性β-糖尿病ラットの細胞が増強されました。 10 mL / kg体重からの濃度は、STZ-NA(ストレプトゾトシン-ニコチンアミド)によって誘発された膵臓の損傷を完全に逆転させ、グリベンクラミドと同様の効果を達成しました[78]。これは、膵臓のβ細胞からのインスリン分泌を増加させることによって血糖値を低下させる、第2世代のスルホニル尿素クラスの抗糖尿病薬です[97]。この効果は、飲料に含まれるアルコールおよびフェノール化合物からのプロトンラジカル捕捉活性に起因していました[78]。この発見は、抗酸化能力が膵臓によるインスリン産生の恒常性を促進する重要な要因でなければならないことを示しています。しかし、この研究では黒米抽出物を添加せずに製剤をテストしなかったため、β細胞に対するこのプラスの効果のどれだけがケフィアだけに起因するのかを判断することは不可能です。実際、黒米抽出物の添加は、DPPH [78]で測定した場合、ケフィア単独と比較して、飲料の抗酸化能力が56.8%増加しました。 |
| したがって、グルコース代謝に対するケフィアの生物活性化合物の効果に関する文献の現在の発見は決定的ではなく、純粋なケフィア飲料またはそれから単離された生物活性化合物を用いたさらなる前臨床研究が緊急に必要とされています。 |
| 3.4.7 脂肪組織における血清コレステロールと脂肪の蓄積への影響 |
| 心血管疾患(CVD)は、世界中の罹患率と死亡率の主要な主要な原因の1つです。 しかし、CVDによる死亡は管理可能な危険因子に起因し、主なものは高い総血清コレステロール、高血圧、および喫煙です[98]。 さらに、高コレステロール血症は肥満に関連するリスク状態の1つです[99]。 このシナリオでは、脂肪細胞の総血清コレステロールと脂肪の蓄積を減らすことは、CVDを防ぐために使用される戦略の1つです。 しかし、肥満を治療するための薬剤は、血圧、肝不全、膵炎、および頭痛に関連するいくつかの副作用を引き起こすと報告されています[100]。 したがって、肥満の治療のための天然化合物の研究が奨励されています。 |
| ケフィアからの生物活性化合物の中で、EPSだけが脂肪細胞脂肪の低下とinvitroコレステロールの低下に関連しています。 Lactococcus lactis WH-C1、Lactobacillus kefiri、およびLeuconostoc mesenteroidesは、ケフィアでこれらのEPSの生産者として同定された微生物でした[77]。 しかし、ミルク中のそれらの前駆体の調査の欠如は、文献にギャップがあることを示しています(表2)。 |
| Tiberケフィア粒から分離されたEPSプロデューサーLactococcus lactis WH-C1(4%、v / v)は、培地から最大31.23%のコレステロールを除去でき、in vitroでの潜在的なコレステロール低下特性を示しています[77]。対照的に、EPSはin vivo実験モデルでは効果がありませんでした。ケフィアから分離されたEPSプロデューサーの熱殺菌乳酸菌(HLAB)の混合物を8週間補給しても、肥満マウスの脂質プロファイルは変化しませんでした。したがって、血清HDL、LDL、総コレステロール、およびトリグリセリドは、食餌誘発性肥満マウスでは影響を受けませんでした[76]。血清コレステロールへの影響がないことは、使用したEPS産生微生物の濃度(1.1×1010 CFU / mL)が、in vivoでコレステロールレベルの有意な変化を達成するには不十分であった可能性があることを示しています。微生物が加熱により死んだときにEPS生産を継続できなくなったことを考慮に入れる必要があります。実際、EPS産生微生物の増殖が大きければ大きいほど、培地からのコレステロール除去率がin vitroで実証されました[77]。 |
| 同様に、腸内細菌叢の調節をもたらさなかったケフィアの補給は、結腸でのプロピオン酸産生の低下と関連しており、血清コレステロールレベルの変化を促進するには不十分です[101]。 また、ポリフェノールが豊富な抽出物(ワイン用ブドウ種子粉-2.5%、v / v)を食事に含めることで、血清コレステロール値は変化しませんでした[76]。 したがって、抗酸化能力は血中コレステロール値を下げる重要な要因ではないかもしれません。 |
| 一方、脂肪細胞における脂肪蓄積を減少させるEPS能力は、in vitroおよびin vivoの動物モデルで確認されました(表2)。 Lactobacillus kefiriおよびLeuconostoc mesenteroidesの菌株から分離されたEPSは、in vitroで3T3-L1脂肪細胞における脂質蓄積の阻害(最大28%)を示しました。 この阻害は、一般に、用量依存的に起こった[76]。 さらに、L.mesenteroidesLMDH4のEPSのみが0.01mg / mLで効果を示したため、効果を得るためのEPSの最小濃度は菌株に依存するようでした。 同時に、L.kefiriLKDH3およびL.mesenteroides LMDH6からのものは、テストされた最高濃度(0.2 mg / mL)でのみ脂肪蓄積に影響を及ぼしました。 対照的に、L.mesenteroidesLMDH8およびLMDH9からのEPSは、試行された最高濃度でも作用しませんでした(表2)。 この発見は、脂肪蓄積に対するEPSの機能的可能性が、その微生物起源によって異なることを意味します。 |
| in vivo動物実験について、EPSプロデューサーHLABを8週間補給した食餌は、高脂肪および高果糖食餌を与えたC57BL / 6Jマウスの脂肪組織重量を36%減少させました[76]。このように、ケフィアの微生物からのEPSは、肥満の場合に使用できる機能性成分である可能性があることを示唆しています。 EPSのパフォーマンスは、脂肪組織への作用によって発生しました。 Wdfc21およびHp炎症性遺伝子の発現は、対照群と比較して、脂肪組織でそれぞれ56%および57%低下しました。また、脂肪酸の合成に関連する遺伝子(Fabp4とFsan)の発現は、HLABグループの脂肪組織でそれぞれ55%と43%ダウンレギュレーションされました。 Fabp4は脂肪酸結合タンパク質4(Fabp4)をコードし、Fsanは脂肪酸シンターゼをコードします。これは、EPSが脂肪細胞の抗炎症と脂肪酸合成の阻害に役割を果たしていることを示しています。ただし、Wdfc21、Hp、Fabp4、およびFsan遺伝子に対するこれらの影響は、HLABでの処理によってレベルが変化していないため、血清コレステロールの変化を引き起こしませんでした。 |
| 興味深いことに、ポリフェノールが豊富なワイン用ブドウ種子粉(2.5%)と組み合わせてHLABを食事に補給すると、脂肪の蓄積を25%減らす効果が高まりました[76]。 したがって、抗酸化活性は脂肪蓄積を減らすための関連要因である可能性があり、脂肪細胞に対するEPSの影響を高めることができます。 その結果、相乗的治療は、血清トリグリセリドを有意に減少させることができる唯一のものでした。 |
| 3.4.8 降圧効果 |
| 高血圧は、血圧の上昇につながる慢性疾患です[102]。 アンジオテンシン変換酵素(ACE)は、哺乳類の適切な血圧レベルを維持するための恒常性メカニズムの一部として、レニン-アンジオテンシンシステムの重要なコンポーネントです。 ACE1からACE2(血管収縮作用を持つ酵素)への変換は、血圧を調節する一般的なメカニズムです[103]。 しかし、異常な状態では、この収縮は高血圧を引き起こし、心臓が体の主要な動脈に血液を送り込むために必要な仕事を増やす可能性があります[104]。 このように、ACE1からACE2への変換の阻害は、高血圧の管理に役割を果たします。 |
| 系統的レビューでは1つの記事しか選択されなかったため、ケフィアからの生理活性化合物が血圧に及ぼす影響を一貫して理解する上での大きな制限でした。 それでも、この研究では、生物活性化合物を生成する微生物、その作用メカニズム、またはミルク中のこの生物活性化合物の前駆体については調査していません。 さらに、研究されたモデルはその場でした。 in vivoでの調査がないため、生理学的システムの血圧に対するケフィアからの生物活性化合物の機能的可能性の理解が制限されます(表2)。 |
| その場で、ACE活性は、ケフィア粒で24時間発酵させた後、ミルクで98.4%減少しました。 観察された効果は、発酵中にケフィアの微生物によって放出された生物活性ペプチドに起因していました[14]。 これは、ケフィア由来の生理活性ペプチドが高血圧の治療に使用される可能性があることを示唆しています。 ただし、将来、より一貫性のある結論に到達するには、さらなる調査が緊急に必要です。 |
| 3.4.9 神経変性疾患に関連する老化モデルにおける抗酸化効果 |
| アルツハイマー病、パーキンソン病、筋萎縮性側索硬化症は、ニューロン細胞の進行性の喪失、運動機能または認知機能の低下、および異常なタンパク質凝集体の蓄積を特徴とする神経変性疾患です[105]。 老化は、アルツハイマー病やパーキンソン病を含むほとんどの神経変性疾患の主要な危険因子です。 65歳以上の10人に1人がアルツハイマー病を患っており、年齢が上がるにつれてこの病気の有病率は増加し続けています[106]。 ミトコンドリアの機能障害とその結果としての老化中の酸化ストレスは、神経変性疾患の進行に大きく影響する確立された要因です[106]。 老化したミトコンドリアでの活性酸素種(ROS)の生成が増加し、膜電位が小さくなり、ATP合成が減少し、呼吸酵素複合体の活性が低下します。 |
| その結果、ミトコンドリアDNAの酸化に加えて、タンパク質の酸化と凝集が起こり、突然変異と欠失が現れ始めます[105]。 加齢に伴う神経変性疾患の効果的な治療法はほとんどまたはまったくないため、不可逆的に進行する傾向があります。 この文脈において、幹細胞治療は、神経細胞集団を再生するための神経変性疾患の治療のために研究された代替方法の1つです[107]。 |
| 現在の系統的レビューでは、神経変性疾患の治療戦略として、ケフィアからの生物活性化合物をテストする2つの適切に管理された研究のみを選択しました。 両方で、EPSの可能性が評価されました(表2)。 神経幹細胞培養(PC12細胞培養)の増殖に対するケフィランの効果は、ナノファイバーを形成するための純粋なポリアクリロニトリル(PAN)に関連する5%および10%のケフィラン濃度についてJenab et al [46]によって調査されました。 ただし、コントロール(PANのみ)と比較した場合、テストした両方のケフィラン濃度は、1日のインキュベーション後にPC12の生存率を最大26.7%低下させ、2日後に有意な効果を示しませんでした。 4日目と6日目に、10%ケフィランはPC12細胞株の生存率をそれぞれ15.4%と21.2%減少させました。 したがって、ケフィランはニューロンの再生を促進するのに適した化合物ではありませんでした。 それとは異なり、10%の濃度から毒性を示しました。 |
| 一方、12週間のEPS(20mLEPS溶液/ kg体重)の経口投与は、EPSが結果として生じる酸化ストレスを軽減できるため、D-ガラクトースで誘発された老化マウスモデルに有益な効果をもたらしました。低用量(1 mg / mL)では、EPSは血清中の総抗酸化能を27.7%向上させました。ただし、総抗酸化能の増加に加えて、EPSは血清グルタチオンペルオキシダーゼ、スーパーオキシドジスムターゼ、およびカタラーゼをそれぞれ21.55、33.14、および61.09%増加させました。報告されている抗酸化酵素の活性の増加に伴い、血清マロンジアルデヒドが49.6%減少しました[83]。次に、マロンジアルデヒドは、パーキンソン病、筋萎縮性側索硬化症、およびアルツハイマー病を含む神経変性疾患の進行の生物学的マーカーとして提案されてきた。これらの疾患では末梢マロンジアルデヒドレベルの上昇が起こるためです[108]。したがって、血清マロンジアルデヒドの減少は、EPSが神経変性疾患の進行を緩和できることを示唆しています。 |
| Lactobacillus plantarum YW11は、抗酸化活性を持つEPS産生微生物として同定されました(表2)。 末梢レベルでのEPSの抗酸化作用は、腸内細菌叢を積極的に調節する能力に起因していました。 この調節効果は、腸管の酸化ストレスの減少と関連していた。 糞便中の酢酸と酪酸の同時増加に伴うNOxの減少は、宿主の腸管の酸化状態を改善するEPSの可能性を示しています(図2)。 興味深いことに、EPSはNOxのレベルを下げるのにアスコルビン酸よりも効率的であり、腸内の短鎖脂肪酸の含有量を上げるのにも同じくらい効率的でした。 EPSの抗酸化能力は、Fe2 +キレート能力に加えて、ヒドロキシルラジカル、DPPHラジカル、およびスーパーオキシドアニオンに対するその捕捉能力に起因する可能性があります[83]。 |
| したがって、ケフィアからのEPSは、治療目的よりも予防効果の点でより有望であるように思われます。 確かに、老化したマウスモデルは、酸化ストレスなどの神経変性疾患の初期段階で提示されたものと同様の特性を示します。 したがって、このモデルは、これらの疾患に関連する最も初期の神経変性変化を研究するために十分に確立されています[109]。 ただし、この仮説を裏付けるには、人間の臨床試験とより前臨床試験を実施する必要があります。 |
| 3.4.10 骨粗鬆症への影響 |
| ここでの系統的レビューは、骨粗鬆症に対する生物活性化合物の効果に関する1つの研究のみを選択しました[80]。 ただし、in vivoの動物実験では、ケフィアで生物活性化合物を生成する微生物や、ミルクでその前駆体を生成する微生物については調査していません。 作用機序も記載されていません。 したがって、骨粗鬆症に対するケフィア生物活性化合物の効果をよりよく理解するために、さらなる研究が緊急に必要とされています。 |
| 雌マウスの卵巣切除処置は、卵巣の除去によるエストロゲンホルモンの低下に関連する骨粗鬆症をもたらした。これらは、骨の骨芽細胞と破骨細胞の活動の不均衡を引き起こしたエストロゲンを生成する主な腺です。その結果、マウスの骨塩密度、海綿骨量、および骨梁数が約16%、70%、および70%減少しました。ただし、ケフィア(100 mg / kg体重)から卵巣切除マウスへの生理活性ペプチドの補給は、骨量減少を抑制する可能性があります。したがって、ペプチドは、骨塩密度、骨梁体積、骨梁数、および骨の弾性率と硬度の機械的特性を、それぞれ41%、264%、235%、42%、および36%増加させました。その結果、大腿骨の小柱分離およびナノインデンテーション領域で36.5%および33%の減少が得られました。骨塩密度、骨梁数、骨梁分離、弾性率、硬度、およびナノインデンテーション領域が陰性対照(非静脈瘤切除女性)のものと類似していたため、ペプチド補給が最終的に骨粗鬆症の状態を逆転させる可能性があることに注意することが重要です。 |
| 骨粗鬆症治療の標準的な手順である炭酸カルシウムの投与も、海綿骨の体積と骨梁の数を増加させる可能性があります。 しかし、これらの値は、生物活性ペプチドの補給によって得られた値よりも約0.7倍低く、骨粗鬆症の治療薬としてのケフィアからのペプチドの可能性を示しています。 最後に、単離された生物活性ペプチドに関するカルシウムと生物活性ペプチドの複合作用の効果に有意差はなく、これら2つの治療の間に相乗効果は見られませんでした[80]。 |
3.5 職人技と工業用ケフィアの生物活性化合物は、機能的可能性の点で異なりますか? |
| 2つのグループが、職人技(ケフィア粒)と工業(スターターカルチャー)のケフィア試験をそれらの機能特性に関して比較するために評価されました(図3)。 まず、抗菌/抗真菌の可能性に関連する効果をターゲットにして、推定ORが2.96(95%CI:0.87–10.05、P = 0.07)になります。 したがって、ケフィアの種類は、その抗菌力と有意に関連していませんでした。 この結果は、両方のタイプのケフィアの抗菌効果の主な原因となる生物活性化合物の1つとして有機酸(特に乳酸)を報告した我々の発見(表2)と一致しています。 |
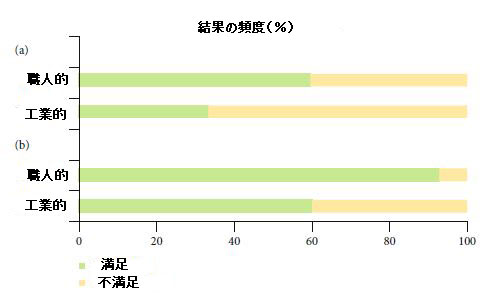 |
| 図3 メタアナリシスに含まれる職人的および工業的ケフィア試験からの有意な満足および不満足な結果の頻度。 (a)抗菌薬の潜在的な結果。 (b)抗酸化剤の潜在的な結果。 |
| 抗酸化能に関する職人的および工業的ケフィアの結果を評価することを目的とした後、別のグループ分析が実行され、8.56の推定ORが得られました(95%CI:2.27–32.21、P≤.001)。したがって、職人技のケフィアを利用すると、工業プロセスのケフィアよりも抗酸化作用を持つ可能性が756%高くなる可能性があります。前に述べたように、フェノール化合物は、工業用ケフィアの抗酸化活性に関与する生物活性化合物でした。対照的に、職人技のケフィアは、生物活性のある抗酸化化合物の多様性を示しました。フェノール化合物に加えて、職人ケフィアには、ケフィラン(職人ケフィア専用構造)を含む生物活性ペプチドとEPSも含まれていました(表2)。工業用ケフィアに含まれるフェノール化合物は還元力がありますが、職人技のケフィアに含まれるフェノール化合物[75]とは異なり、ラジカル捕捉能[85]の点では効果がありませんでした。これらの発見は、抗酸化能に影響を与える可能性のある両方のケフィア間のフェノール化合物の異なるプロファイルを示唆しています。さらに、職人技のケフィアに含まれる生物活性化合物の多様性が、このタイプのケフィアの抗酸化能力を強化するのに貢献した可能性があります(表2)。 |
| 職人によるケフィアに存在する抗酸化化合物の生産者として報告されている微生物[28、73]は工業用ケフィアには一般的に見られないため、抗酸化能の違いはケフィアの種類間の微生物プロファイルの違いに関連している可能性があります(表1)。 確かに、職人のケフィアのより多様な微生物プロファイルは、工業用ケフィアよりも優れた機能的利益を与えることに責任があると示唆されています[20]。 一貫して、前述のように、職人のケフィアで劇的に大きな真菌の多様性を観察しました。 さらに、両方のケフィア間の細菌組成は特異性を示しました(表1)。 したがって、職人のケフィアの微生物プロファイルは、抗酸化能を持つ生物活性化合物のより多様性を生み出すことに関連しているようです。 |
| ただし、職人によるケフィアは主にケフィア研究に選択されることが多いため、産業用ケフィアの所見を報告する研究はメタアナリシスに含まれているだけであり、ケフィアの種類を比較する際の制限要因であったことを報告する必要があります。 |
4.結論と展望 |
| このレビューは、予防および治療薬としてのケフィアからの生物活性化合物の可能性を強調しています。ケフィア投与の健康上の利点に取り組む研究はたくさんあります。ただし、文献の制限は、制御されていない実験であるか、観察された健康上の利点の原因となる生物活性化合物を特定していないことです。さらに、生物活性化合物を調査する研究の相対的な不足-ミルク中の生産微生物とその基質は、文献のギャップを構成します。メタアナリシスは、抗菌、抗癌、および免疫調節活性を裏付けました。ケフィランと乳酸は、世界的な抗菌作用を持つ主成分でした。しかし、ケフィランは、静菌効果ではなく殺菌効果を発揮する濃度閾値を示しました。同時に、乳酸は用量依存的に細菌胞子の発芽を抑制しました。それでも、ケフィランを含むEPSは、主に結腸がんと乳がんに対する活動に関与していました。それにもかかわらず、EPS濃度と介入時間は、抗がん効果を得るかどうかを決定するものであり、EPSは子宮頸がんと結腸がんに対して用量依存的な阻害効果を示しました。 |
| 抗炎症作用に関しては、EPS、細胞外小胞、および乳酸が関与する主な生物活性剤でした。ただし、EPSは、その濃度と環境条件(炎症性発作の有無)に応じて、免疫賦活効果も持つ可能性があります。免疫刺激を行うには、抗炎症効果に使用される介入時間と同様の介入時間にもかかわらず、EPS濃度を劇的に高くする必要がありました。細胞外小胞の高用量は、低用量よりも大腸炎に対してより効果的であるように思われました。したがって、これらの要因は、将来の臨床応用で慎重に検討する必要があります。研究がinvitroおよびinvivo実験に限定されたことは注目に値するので、ケフィアからの生物活性化合物の実用化を進めるために臨床的証拠が緊急に必要とされています。生物活性化合物の作用機序は多様であり、異なるシグナル伝達経路によって作用できることを示しています。抗酸化活性と腸の調節はケフィアの有効性を支持する傾向を示していますが、おそらくデータの不均一性が高いため、統計的有意性には達しませんでした。したがって、これらの効果を評価するために使用される方法論の標準化は、将来のさまざまな研究を比較するのに役立ちます。血漿グルコースレベル、神経変性疾患、脂質プロファイル、血圧、および骨粗鬆症に関する生物活性化合物は決定的ではなく、一貫した結論を得るにはさらなる研究が必要です。最後に、職人技と工業用ケフィアは、健康への影響に応じて異なる機能的可能性を持っている可能性があり、これは両方のタイプのケフィア間の微生物組成の違いに関連している可能性があります。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
この文献は、Oxidative Medicine and Cellular Longevity Volume 2021, Article ID 9081738, 34 pages に掲載されたBioactive Compounds from Kefir and Their Potential Benefits on Health : A Systematic Review and Meta-Analysis を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |