ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(発酵乳、腸内細菌の科学:研究の最前線) |
胆道がんにおける胆汁酸代謝と腸内細菌叢の相互作用 |
Ifeoma Ike et al., |
|
|
| 要約 |
腸内細菌叢と胆汁酸(BA)は、宿主の代謝、免疫機能、および疾患に影響を及ぼす、厳密に制御された双方向的な関係にあります。肝臓で合成された一次胆汁酸は、腸内細菌によって化学的に変換され、多様な二次胆汁酸のプールを形成します。これらは抗菌作用を発揮するとともに、ファルネソイドX受容体(FXR)(訳者注:ファルネソイドX受容体(FXR)は、主に肝臓や小腸などに存在する核内受容体(転写因子)の一つです。胆汁酸を主なリガンド(結合物質)とし、脂質や糖の代謝、胆汁酸の合成・輸送などを調節する重要な役割を担っています)、Takeda Gタンパク質共役受容体5(TGR5)(訳者注:Takeda Gタンパク質共役受容体5(TGR5)は、主に胆汁酸をリガンド(結合物質)とする膜受容体です。武田薬品工業によって発見されたこの受容体は、代謝や免疫系の調節に深く関わっており、肥満や糖尿病などの治療標的として注目されています)、スフィンゴシン-1-リン酸受容体2(S1PR2)(訳者注:スフィンゴシン-1-リン酸受容体2(S1PR2)は、生理活性脂質S1Pのシグナルを伝達するGタンパク質共役受容体 です。血管バリアの破綻、アレルギー反応、がん細胞の増殖 に深く関わるほか、心筋梗塞後の細胞修復 や骨形成 においても重要な役割を果たす注目すべき分子標的です)などの宿主シグナル伝達経路を活性化します。これらの経路は、胆汁酸の恒常性、上皮バリア機能の維持、炎症、および発がんを制御しています。この胆汁酸-腸内細菌叢軸の破綻は、世界的に発生率が増加しており治療選択肢も限られている悪性度の高い腫瘍群である胆道がん(BTC)に関与していることが示唆されています。二次胆汁酸や胆汁酸受容体シグナル伝達は、NF-κB(訳者注:NF-κB(核内因子κB)は、ほぼすべての動物細胞に存在する重要なタンパク質複合体(転写因子)です。免疫や炎症反応の中心的役割を担い、ストレスやウイルスなどの刺激に応答して特定の遺伝子の発現を制御します)の活性化、酸化ストレス、細胞生存の変化を介して腫瘍の発生と進行に寄与する一方、ファルネソイドX受容体シグナルの低下や腸肝循環の阻害は、炎症の調節不全をさらに助長します。新たな知見により、腸内細菌叢のディスバイオシス(腸内菌叢バランスの乱れ)や胆汁酸代謝の変化が、胆道がん特有の細菌叢プロファイルと関連していることが明らかになっています。このプロファイルは、フソバクテリウム属、サルモネラ属、プレボテラ属、アクチノマイセス属などの分類群の増加と、ラクトバチルス属 などの常在菌の減少を特徴とします。これらの分類群は、炎症シグナル伝達、胆汁酸変換、および上皮損傷に影響を及ぼし、発がんに寄与します。腸内細菌叢と胆汁酸の相互作用は、抗腫瘍免疫や免疫チェックポイント阻害薬(ICI)への応答にも影響を与えます。特定の細菌叢シグネチャー(特に ラクノスピラ科、エリシペロトリクス科、バクテロイデス門、アリスティペス属の増加)は、肝胆道がんにおける免疫活性化の増強や臨床転帰の改善と相関しています。抗生物質、プロバイオティクス、または糞便微生物叢移植による腸内細菌叢の調節は、胆汁酸組成、免疫監視機構、および治療効果に影響を及ぼし得ます。以上のデータは、胆道がんの病因および治療反応において、胆汁酸・細菌叢軸が中心的な役割を果たしていることを浮き彫りにしています。腸内細菌叢および胆汁酸代謝物のプロファイリングはバイオマーカー開発の有望な手段であり、一方で胆汁酸シグナル伝達や微生物生態系の標的を絞った制御は、胆道がんの転帰を改善するための潜在的な治療戦略となります。 |
| 目次(クリックして記事にアクセスできます) |
| 1.はじめに |
| 2.胆汁酸の概要 |
| 3.胆汁酸-腸内細菌叢軸 |
| 4.腫瘍生物学における胆汁酸 |
| 5.胆汁酸、腸内細菌叢、および胆道がんの相互作用 |
| 6.免疫調節および耐性メカニズムにおける役割 |
| 7.免疫療法、腸内細菌叢、および胆道がんの相互作用 |
| 8.臨床的に一般的な交絡因子 |
| 9.今後の展望と結論 |
本文 |
| 1.はじめに |
| 腸内細菌叢と胆汁酸は、厳密に制御された双方向的な関係にあり、宿主の代謝、免疫、および疾患において根本的な役割を果たしています。腸内細菌叢とは、細菌、古細菌、ウイルス、真核生物など、消化管内に存在する微生物の集合体を指します。胆汁酸は肝臓でコレステロールから合成され、その後、腸内微生物によって修飾を受け、それぞれ異なる生物学的特性を持つ多様な二次胆汁酸へと変換されます。こうした微生物による変換は、胆汁酸プールの化学的多様性を大幅に拡大させるとともに、抗菌防御、代謝恒常性、上皮バリア機能、さらには炎症-免疫-発がんを制御する受容体介在性シグナル伝達経路を調節することで、宿主の生理機能に影響を及ぼします(Guo et al., 2022)。一方で、胆汁酸はシグナル伝達分子、抗菌物質、代謝調節因子として機能し、微生物叢の構成を形成するとともに宿主の遺伝子発現を調節することで、「胆汁酸-微生物叢軸」を形成しています(Kim et al., 2007; Fiorucci et al., 2009; Vavassori et al., 2009; Guo et al., 2022)。 |
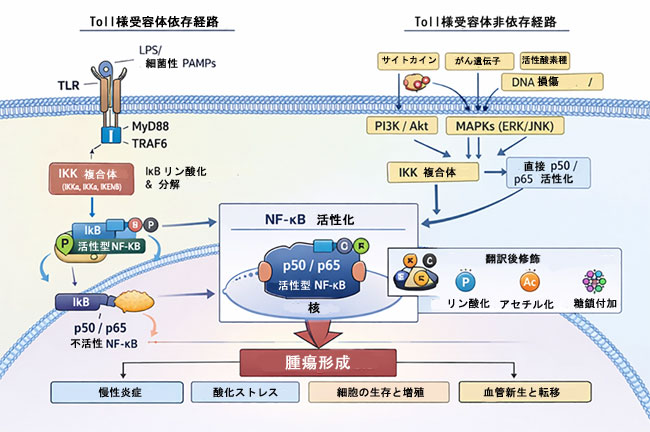 |
図1. 腫瘍形成における標準的NF-κBシグナル伝達経路:TLR依存性およびTLR非依存性の活性化機構。 本図は、TLR依存性シグナル伝達(LPS/PAMP–TLR–MyD88/TRAF6–IKK複合体–IκB分解)を介したNF-κBの活性化と、サイトカイン、がん遺伝子、活性酸素種、DNA損傷、およびPI3K/AktやMAPK経路が関与し、IKKまたはp50/p65に直接収束するTLR非依存性機構による活性化を示している。いずれの経路も、最終的には標準的NF-κBヘテロ二量体(p50/p65)の核内移行に至る。この二量体の転写活性は、リン酸化、アセチル化、糖鎖付加などの翻訳後修飾によって精密に制御されている。活性化したNF-κBは、慢性炎症、酸化ストレス応答、細胞生存、増殖、血管新生、転移に関連する遺伝子プログラムを駆動し、それによって腫瘍形成を促進する。 略語:IKK、IκBキナーゼ;LPS、リポ多糖;NF-κB、核内因子κB;PAMPs、病原体関連分子パターン;ROS、活性酸素種;TLR、Toll様受容体。 |
| 2.胆汁酸の概要 |
| 胆汁酸はコレステロール代謝の最終産物であり、ヒトにおけるコレステロール排泄の主要な経路を担っています。その合成は、胆汁の形成を促進するだけでなく、リン脂質、コレステロール、外因性物質、および内因性老廃物の排泄も可能にします(Ridlon et al., 2016; Guo et al., 2022)。 |
| 胆汁酸は肝臓において、コレステロールのステロール環の修飾および側鎖切断反応を経て、主にコール酸やケノデオキシコール酸といった一次胆汁酸として合成されます。その後、これらはグリシンやタウリンと抱合して胆汁酸塩を形成します(Rembacz et al., 2010; Ridlon et al., 2016)。これらの胆汁酸塩は、リン脂質やコレステロールと集合して混合ミセルを形成し、胆嚢に貯蔵された後、小腸へと放出されます。小腸では、食事由来の脂質や脂溶性栄養素の乳化および吸収を助ける役割を果たします。小腸での機能を終えた後、胆汁酸の大部分は回腸で能動的に再吸収され、門脈を介して肝臓へと戻ります(Ridlon et al., 2016)。この極めて効率的な腸肝循環は、消化や解毒だけでなく、代謝バランスの維持においても中心的な役割を担っています(Chiang, 2013; Sannasiddappa et al., 2017)。 |
| 腸管内において、胆汁酸塩加水分解酵素は一次胆汁酸を脱抱合し、その疎水性と上皮透過性を増大させる(Ridlon et al., 2016)。胆管系内におけるこれら非抱合型胆汁酸の蓄積は、胆管上皮細胞の膜損傷、ミトコンドリア機能障害、およびNF-κBを含む炎症シグナル伝達経路の活性化を促進する。その後、コール酸やケノデオキシコール酸といった非抱合型胆汁酸は、胆汁酸誘導性オペロン(*bai*)によってコードされる酵素の作用により、二次胆汁酸であるデオキシコール酸およびリトコール酸へと変換される(Ridlon et al., 2016; Song et al., 2018; Guo et al., 2022)。これら二次胆汁酸はより高い細胞毒性および遺伝毒性を有し、酸化的DNA損傷を誘発するとともに、Takeda Gタンパク質共役受容体5などの増殖シグナル伝達経路を活性化させることで、胆管上皮細胞の増殖およびアポトーシスへの抵抗性を亢進させる。さらに、微生物由来のヒドロキシステロイド脱水素酵素が介在する酸化反応やエピマー化反応は、ファルネソイドX受容体シグナル伝達を含む胆汁酸受容体の活性を修飾し、炎症状態や胆汁酸の恒常性を変化させる。総じて、こうした微生物による変換作用は、胆汁酸プールをより疎水性が高く炎症惹起性の強い成分へとシフトさせ、上皮損傷、慢性炎症、および胆管がんの発生を助長する増殖シグナル伝達を促進する(Ridlon et al., 2016)。 |
| 3.胆汁酸-腸内細菌叢軸 |
| 微生物群集は胆汁酸プールを形成します。胆汁酸の組成変化や特定の濃度レベルは、細菌や潜在的な病原体の過剰増殖を誘発する可能性があります(di Gregorio et al., 2021)。二次胆汁酸は細菌の細胞膜に対して強力な界面活性剤様作用を示し、濃度依存的に静菌作用や殺菌作用を発揮し得ます。一次胆汁酸の蓄積は、特定の細菌種において細胞内pHを低下させ、膜電位の著しい乱れとそれに続く細菌増殖の阻害を引き起こす可能性があります(di Gregorio et al., 2021; Wu et al., 2025)。 |
| 胆汁酸は、抗菌作用に加え、ファルネソイドX受容体やTakeda Gタンパク質共役受容体5といった宿主受容体のリガンド(結合物質)としても機能します(Wu et al., 2025)。健全な腸管上皮バリアは、腸管内腔の微生物産物への全身的な曝露を制限すると同時に、制御性T細胞の誘導やIL-10、TGF-βといった抗炎症性サイトカインの産生を介して免疫寛容を促進します。これらの過程は、短鎖脂肪酸や、ファルネソイドX受容体やTakeda Gタンパク質共役受容体5を含む胆汁酸活性化受容体によって部分的に制御されています(Rooks and Garrett, 2016)。このバリアの維持は、粘膜および肝胆道系の炎症を抑制する上で極めて重要です。一方、バリアが破綻すると、微生物のトランスロケーションやエンドトキシンに起因するサイトカインシグナル伝達が生じ、これらが胆道内における慢性炎症や腫瘍形成を促進する環境の一因となる可能性があります。 |
| ファルネソイドX受容体の活性化は、胆汁酸の合成と輸送を制御し、腸管上皮バリア機能を維持します(Erice et al., 2018; Cossiga et al., 2023)。ファルネソイドX受容体は、胆汁酸合成の律速酵素であるコレステロール7α-水酸化酵素を抑制することで、胆汁酸産生に対する負のフィードバック制御を行います。さらに、ファルネソイドX受容体の活性化は胆汁酸の排出を促進するトランスポーターの発現を高め、それによって肝細胞毒性を軽減します(Wu et al., 2025; Yan et al., 2025)。胆管上皮細胞を含む非実質細胞に発現するTakeda Gタンパク質共役受容体5は、状況に応じて炎症、代謝、細胞増殖を調節します(Fiorucci et al., 2009; Wu et al., 2025)。重要な点として、抱合型および非抱合型胆汁酸はこれらの受容体を異なる様式で活性化し得ます。具体的には、非抱合型胆汁酸はファルネソイドX受容体をより強く活性化する一方、抱合型胆汁酸はTakeda Gタンパク質共役受容体5や、スフィンゴシン-1-リン酸受容体2(S1PR2)を介した炎症促進シグナル伝達を優先的に活性化する傾向があります(Fiorucci et al., 2009; Song et al., 2018; Wu et al., 2025)。スフィンゴシン-1-リン酸受容体2は、特に胆管上皮細胞や肝星細胞において、炎症反応や線維化反応の促進に関与していることが示唆されています(Wu et al., 2025)。そのシグナル伝達活性は、肝障害や胆管癌の発生に寄与している可能性があります(Thibaut and Bindels, 2022)。 |
| 4.腫瘍生物学における胆汁酸 |
| 胆汁酸は、消化や腸内細菌叢の制御といった役割に加え、宿主のシグナル伝達経路への作用を通じて腫瘍生物学に影響を及ぼします。中でも特によく研究されているのが、ファルネソイドX受容体やTakeda Gタンパク質共役受容体5が関与するシグナル伝達経路です。これらはいずれも胆汁酸(特に二次胆汁酸)によって直接活性化され、がんの病態形成に関与しています(Cossiga et al., 2023; Wu et al., 2025)。 |
| ファルネソイドX受容体は肝細胞および胆管上皮細胞において腫瘍抑制因子として機能する(Kim et al., 2007)。抱合を受けていない胆汁酸(主にケノデオキシコール酸およびコール酸)によるファルネソイドX受容体の活性化は、胆汁酸合成の抑制、脂質合成の低下、および抗炎症性の遺伝子発現誘導をもたらす(Kim et al., 2007)。肝細胞において、ファルネソイドX受容体の活性化はCYP7A1(訳者注:CYP7A1(コレステロール7α-水酸化酵素)は、肝臓でコレステロールから胆汁酸を合成する過程における「律速酵素」です。体内のコレステロール恒常性や脂質代謝をコントロールする上で非常に重要な役割を果たしています)の発現抑制を介して胆汁酸合成を抑制する(Kim et al., 2007)。胆管がんにおいては、隣接する正常肝組織と比較して腫瘍組織でのファルネソイドX受容体発現が著しく低下しており、ファルネソイドX受容体発現の低さは、分化度の低下、腫瘍病期の進行、および血管浸潤の増大と相関している(Bernstein et al., 2009; Dai et al., 2013)。ファルネソイドX受容体の薬理学的活性化(特にオベチコール酸によるもの)は、胆管がん細胞株の増殖および遊走を阻害し、マウス異種移植モデルにおける腫瘍増殖を抑制することが示されている(Dai et al., 2013)。これらの胆管がんに特異的な知見は、胆管がんにおけるファルネソイドX受容体の腫瘍抑制的役割が状況依存的であることを裏付けている。しかしながら、ファルネソイドX受容体シグナル伝達は肝細胞と胆管上皮細胞の間やリガンド(結合物質)の種類によって異なる可能性があり、その腫瘍抑制効果をすべての胆汁酸に一律に当てはめるべきではない。 |
| 抱合型胆汁酸によって活性化されるTakeda Gタンパク質共役受容体5は、状況に応じて異なる役割を果たすと考えられています(Song et al., 2018; Jia et al., 2020)。非悪性の胆管上皮細胞において、Takeda Gタンパク質共役受容体5の活性化は細胞内cAMP濃度を上昇させ、細胞生存を促進するとともに、胆汁酸による傷害から細胞を保護します。この保護作用は、「重炭酸塩アンブレラ(bicarbonate umbrella)」の増強を介して発揮されます。重炭酸塩アンブレラとは、塩化物イオン/重炭酸イオン交換機構によって細胞頂端膜表面にアルカリ性の微小環境を維持し、胆汁酸の毒性から胆管上皮細胞を保護する仕組みのことです(Beuers et al., 2010)。Takeda Gタンパク質共役受容体5は主に二次胆汁酸、特にリトコール酸やデオキシコール酸によって活性化されますが、その抱合体による活性化も認められています(Fiorucci et al., 2009)。一方、悪性細胞においては、Takeda Gタンパク質共役受容体5の活性化は細胞増殖、アポトーシス抵抗性、および活性酸素種の産生増加に寄与し、腫瘍形成を促進する環境を作り出します。 |
| タウロコール酸などの抱合型胆汁酸もまたスフィンゴシン-1-リン酸受容体2を活性化し、COX-2のアップレギュレーションやプロスタグランジンE2産生の増加を介して、炎症促進および発がん性のシグナル伝達経路をさらに増幅させることが示されている(Wang et al., 2017; Song et al., 2018)。マウスの胆汁うっ滞モデル(胆管癌モデルではないが、関連する知見として)において、スフィンゴシン-1-リン酸受容体2の活性化は、ERK1/2(訳者注:ERK1/2(細胞外シグナル調節キナーゼ)は、細胞の増殖、生存、分化をコントロールする重要なシグナル伝達経路(MAPK経路)の中心的な酵素です。外部からの刺激を細胞の核に伝え、適切な細胞応答を引き起こします)およびAKTシグナル(訳者注:AKT (プロテインキナーゼB) シグナル伝達は、細胞の生存、増殖、成長、代謝を調節する中心的な経路です。成長因子などの細胞外刺激により活性化され、アポトーシス(細胞死)の抑制や糖代謝の促進を行いますが、経路の異常はがんや糖尿病などの疾患に関与します)伝達やCOX-2(訳者注:COX-2(シクロオキシゲナーゼ-2)は、炎症や痛みの原因となる物質(プロスタグランジン)を作り出す酵素です。通常は体内に存在しませんが、炎症や組織の損傷が起きた際に急激に増加するため、炎症と痛みのメディエーターとして知られています)発現の増加を介して、胆管上皮細胞の増殖および肝障害を促進した。胆管がんを直接対象としたモデルでの研究は限られているものの、これらの経路は胆管がんに関与する発がん性シグナル伝達ネットワークと重複しており、腫瘍に伴う胆管増殖に対しても同様の機序が関与している可能性が示唆される。 |
| 腫瘍生物学に対する胆汁酸の影響に関与するもう一つのシグナル伝達経路として、炎症および細胞生存の主要な制御因子である核内因子カッパB(NF-κB)が挙げられます(Wang et al., 2017)。in vitro研究により、非抱合型胆汁酸は高濃度においてNF-κBの活性化を阻害し、胆管癌細胞の増殖を抑制する一方、抱合型胆汁酸はNF-κBを活性化させ、IL-6およびCOX-2の発現を亢進し、腫瘍の増殖を促進することが示されています(Wang et al., 2017)。図1には、関与するシグナル伝達経路がより詳細に示されています(Zhang et al., 2021)。 |
| これらの経路のいずれかに調節不全や炎症状態が生じると、免疫・炎症細胞の動員や、サイトカインおよび急性期反応物質の異常産生が引き起こされます(Zhao et al., 2025)。がんなどの病態においては、腫瘍が胆汁の流れを阻害し、それによって胆汁酸やその他の胆汁代謝産物の腸肝循環が遅延する可能性があります(Staley et al., 2017; Pezzino et al., 2022)。 |
| 5.胆汁酸、腸内細菌叢、および胆道がんの相互作用 |
| 胆道がんと腸内細菌叢のディスバイオシス(腸内細菌叢のバランスの乱れ)との関連性が近年指摘されています(Sharma et al., 2007; Ye et al., 2023)。微生物群集およびその代謝産物、とりわけ胆汁酸の変化は、現在、胆道がんの発症に関与する要因の一つとして認識されています(Song et al., 2018; Jia et al., 2020)。胆道系や消化管内における慢性的ディスバイオシスや持続的な感染は、持続的な炎症、発がん性代謝産物の産生、宿主のエピジェネティックな制御の変化など、いくつかのメカニズムを介して発がんに寄与します。分子生物学的解析により、胆道悪性腫瘍患者の胆汁中には、非悪性疾患患者とは異なる微生物プロファイルが存在することが明らかになっています(Sharma et al., 2007)。これらの微生物プロファイルにおいて頻繁に検出される細菌には、放線菌、腸チフス菌、フソバクテリウム、カンピロバクター、ヘリコバクター・ピロリ、プレボテラなどが含まれます(Endt et al., 2010; Kamada et al., 2013; Saab et al., 2021; Ye et al., 2023)。 |
| いくつかの研究において、病原性細菌の高レベルな存在と胆道がんとの関連が示唆されています。肝外胆管癌患者100名を対象にDNAを抽出し、微生物叢の特性を解析したAvilés-Jiménezらによる論文では、フソバクテリウム属、プレボテラ属、ヘリコバクター・ピロリ、およびカンピロバクター属が高頻度で検出されました。これらの菌は、短鎖脂肪酸産生菌の減少や上皮バリア機能の障害を伴う制御不能な炎症反応を介して、胆道がんの発症に関与している可能性が示唆されています(Tan et al., 2014; Avilés-Jiménez et al., 2016; Saab et al., 2021)。また、胆管内の微生物叢においてプロテオバクテリア門:カンピロバクター属を含む)が約60%を占めることが明らかになりました。この構成は小腸の微生物叢と酷似しており、腸内細菌叢と胆道系における病原性環境との間に関連がある可能性を示唆しています(White et al., 2006; Kamada et al., 2013; Wu et al., 2013)。本研究では胆汁酸の直接的な測定は行われていませんが、プロテオバクテリア門の増殖は、胆汁酸の脱抱合能の亢進や炎症シグナルの活性化と広く関連していることが知られています(これらの現象は肝胆道系悪性腫瘍に共通して認められます)(Avilés-Jiménez et al., 2016; Jia et al., 2020)。ただし、これらの知見は肝外胆管がんに特有のものであり、肝内胆管がんに対して慎重な検討なしにそのまま適用すべきではありません。 |
| 対照的に、肝内胆管がんは、局所的な胆管内への細菌定着よりも、むしろ腸内細菌叢や胆汁酸代謝の変化とより強く関連していることが示唆されています。Jiaらは、16S rRNAシーケンシングを用いて、肝内胆管がん患者、肝細胞がん患者、および健常者の間で腸内細菌叢のプロファイルが異なることを明らかにしました。84名の患者を対象とした解析において、肝内胆管がん患者(24名)では、ラクトバチルス属、放線菌属、およびペプトストレプトコッカス科の細菌の増加(濃縮)が認められました。特にラクトバチルス属と放線菌属の増加は、肝内胆管がんを肝細胞がんや肝硬変と鑑別する上で、診断に役立つ可能性が示されています。また、肝細胞がん患者や健常者と比較して、肝内胆管がん患者では抱合型胆汁酸であるタウロウルソデオキシコール酸(TUDCA)の増加が認められ、この増加はラクトバチルス属および放線菌属の菌量の増加と正の相関を示しました(Jia et al., 2020)。これらの知見は、肝内胆管がん(iCCA)における「胆汁酸-腸内細菌叢」軸の存在を裏付けるものですが、解析対象のコホートが肝疾患と胆道疾患の混合集団であったため、この関連性が肝内胆管がんに特異的なものであると断定することはできません。 |
| 対照的に、肝内胆管がんは、局所的な胆管内への細菌定着よりも、むしろ腸内細菌叢や胆汁酸代謝の変化とより強く関連していることが示唆されています。Jiaらは、16S rRNAシーケンシングを用いて、肝内胆管がん患者、肝細胞がん患者、および健常者の間で腸内細菌叢のプロファイルが異なることを明らかにしました。84名の患者を対象とした解析において、肝内胆管がん患者(24名)では、ラクトバチルス属、放線菌属、およびペプトストレプトコッカス科の細菌の増加(濃縮)が認められました。特にラクトバチルス属と放線菌属の増加は、肝内胆管がんを肝細胞がんや肝硬変と鑑別する上で、診断に役立つ可能性が示されています。また、肝細胞がん患者や健常者と比較して、肝内胆管がん患者では抱合型胆汁酸であるタウロウルソデオキシコール酸(TUDCA)の増加が認められ、この増加はラクトバチルス属および放線菌属の菌量の増加と正の相関を示しました(Jia et al., 2020)。これらの知見は、肝内胆管がん(iCCA)における「胆汁酸-腸内細菌叢」軸の存在を裏付けるものですが、解析対象のコホートが肝疾患と胆道疾患の混合集団であったため、この関連性が肝内胆管がんに特異的なものであると断定することはできません。 |
| 表1 言及された細菌/分類群およびそれらと胆道がんとの関連の概要 |
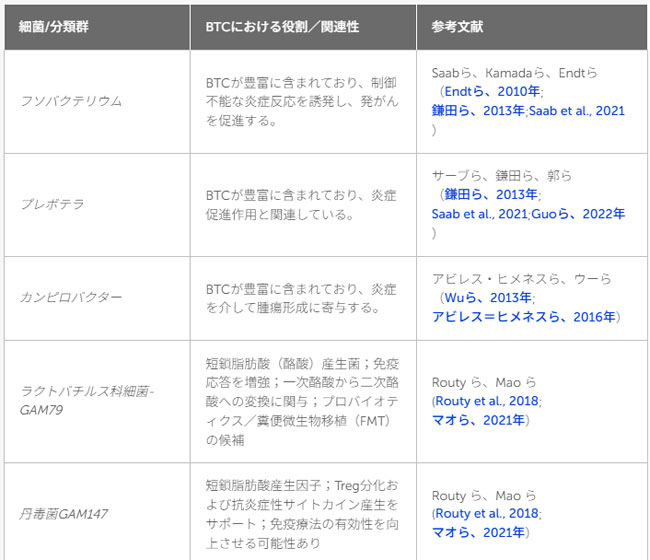 |
 |
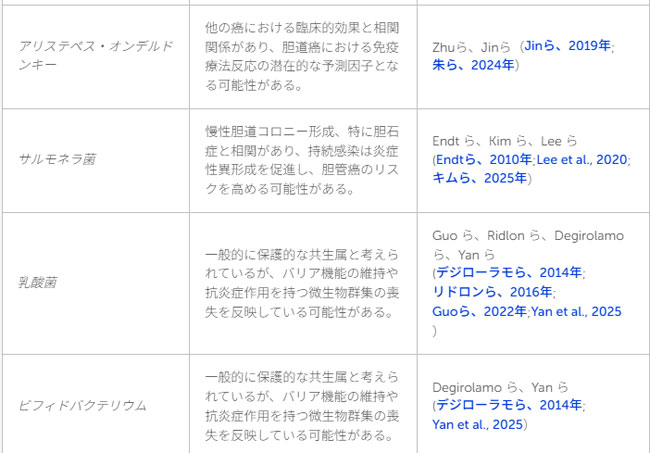 |
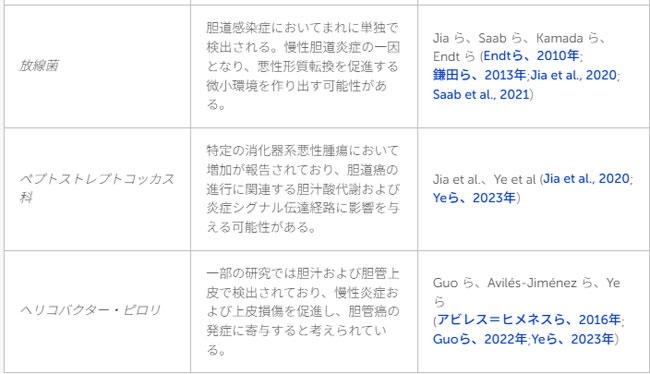 |
| BTC:胆道がん、FMT:糞便微生物叢移植、SCFA:短鎖脂肪酸、T-reg:制御性T細胞。 |
| 6.免疫調節および耐性メカニズムにおける役割 |
| 近年の研究により、肝・胆道系悪性腫瘍における腸内細菌叢、胆汁酸代謝、および免疫調節の間の重要な関連性が明らかになっています。特筆すべき発見として、CXCL16-CXCR6軸(訳者注:CXCL16-CXCR6軸は、免疫細胞(主にT細胞や自然リンパ球など)の表面にある受容体「CXCR6」と、その特異的リガンドである「CXCL16」が結合するシグナル伝達経路です。免疫細胞を特定の組織や炎症部位へ誘導・定着させる役割を持ち、がん治療、アルツハイマー病、アレルギー性疾患など幅広い分野で研究されています)を介した一次胆汁酸と肝臓のナチュラルキラーT(NKT)細胞との相互作用が挙げられます(Ma et al., 2018; Wu et al., 2025)。マウスモデルにおいて、一次胆汁酸は肝類洞内皮細胞におけるCXCL16(訳者注:CXCL16(C-X-C motif chemokine ligand 16)は、免疫細胞の遊走や活性化、接着を制御する独自の構造を持つ膜結合型および可溶型のケモカインです。主に免疫応答、炎症、癌の進行、脂質代謝などの多様な生命現象に関与しています)の発現を促進し、それによって抗腫瘍効果を発揮するCXCR6(訳者注:CXCR6(C-X-Cモチーフ ケモカイン受容体6)は、免疫細胞の表面に存在するGタンパク質共役受容体の一種です。リガンドであるケモカイン「CXCL16」と結合することで、免疫細胞を炎症部位や特定の臓器へ遊走・配置させる重要な役割を担っています)陽性ナチュラルキラーT細胞の動員と活性化を誘導します(Wu et al., 2013)。この免疫監視機構は、デオキシコール酸やリトコール酸といった二次胆汁酸によって抑制されます(Wu et al., 2013)。 |
| 抗生物質を用いてグラム陽性の胆汁酸変換細菌を減少させるなど、一次胆汁酸の蓄積を促すように微生物叢を操作することは、ナチュラルキラーT細胞を介した腫瘍抑制を増強し得ます。こうした知見は、TIGER-LC試験を含むヒトコホート研究でも裏付けられており、そこではケノデオキシコール酸(一次胆汁酸の一種)の高値が、CXCL16発現の増大およびグリコリトコール酸濃度の低下と相関することが示されています(Chaisaingmongkol et al., 2017; Ma et al., 2018; Wu et al., 2025)。 |
| Yanらによる、核内受容体シグナル伝達および胆汁酸-マイクロバイオーム軸におけるその役割に関する総説では、ラクトバチルス属やビフィドバクテリウム属といった特定の微生物叢の減少がファルネソイドX受容体の枯渇を招き、それによって一次胆汁酸(具体的にはコール酸およびケノデオキシコール酸)の合成が抑制されることが示されました。この過程は炎症の調節不全を促進し、結果として肝胆道系疾患や腸管疾患を引き起こす可能性があります(Yan et al., 2025)。 |
| メカニズムに関する研究の多くは前臨床モデルや胆管系以外の腫瘍モデルに基づいたものですが、脂質シグナル伝達、微生物代謝、免疫調節の相互作用を考慮すると、脂肪酸アミド加水分解酵素(FAAH)および関連するエンドカンナビノイド・システムの構成要素が、炎症を基盤とするがんにおける腫瘍の挙動や治療反応性を調節する因子であることが示唆されます。脂肪酸アミド加水分解酵素は膜結合型セリン加水分解酵素であり、エンドカンナビノイドであるアナンダミドや、それに関連するN-アシルエタノールアミン類(パルミトイルエタノールアミドやオレオイルエタノールアミンなど)といった内因性アミド化脂質のシグナル伝達を終結させることで、エンドカンナビノイド・システムのトーン(活性レベル)を調節しています(Cravatt et al., 1996; V et al., 2004)。これらの脂質メディエーターは、カンナビノイド受容体1(CB1)、カンナビノイド受容体2(CB2)、およびペルオキシソーム増殖剤活性化受容体α(PPAR-α)のシグナル伝達を調節することを通じて、代謝恒常性、神経炎症、痛覚、細胞増殖に影響を及ぼします(D, 2003)。がん生物学において、FAAH脂肪酸アミド加水分解酵素の発現や活性の変化はアミド化脂質レベルの異常と関連しており、その影響(腫瘍増殖、アポトーシス、血管新生、免疫細胞の動員など)は状況によって異なります。例えば、一部の悪性腫瘍では、脂肪酸アミド加水分解酵素活性の低下がエンドカンナビノイド・シグナルを増強し、カンナビノイド受容体1(CB1)/カンナビノイド受容体2(CB2)を介した経路を通じて増殖を抑制する可能性がありますが、別の腫瘍では、エンドカンナビノイド・システムの活性化が腫瘍の進行や免疫回避に関連していることが報告されています(Maccarrone et al., 2015; Morena et al., 2016)。さらに、近年の研究により、腸内常在細菌叢とエンドカンナビノイド・システムとの間の双方向的なクロストークが明らかになりつつあります。短鎖脂肪酸や食事由来の多価不飽和脂肪酸などの微生物代謝産物は、N-アシルエタノールアミンの合成・分解、腸管透過性、全身性の炎症シグナル伝達に影響を及ぼすことで、エンドカンナビノイド・システムのトーンを調節し得ます(M et al., 2010; S and V, 2013; Cani et al., 2014)。逆に、宿主のエンドカンナビノイド・システムの活性は、腸管バリアの完全性や微生物叢の構成を形成し、代謝や免疫に関するシグナルを統合するフィードバックループを構築しています。腫瘍領域において、このマイクロバイオーム・エンドカンナビノイド軸は、炎症性サイトカイン・ネットワーク、骨髄系細胞の分極、酸化ストレス、ならびに化学療法や免疫療法に対する抵抗性を変化させることにより、がんの進行や治療反応性に影響を及ぼす可能性がある(Sido et al., 2016)。 |
| 7.免疫療法、腸内細菌叢、および胆道がんの相互作用 |
| 胆汁酸-腸内細菌叢軸の破綻は、免疫チェックポイント阻害薬(ICI)への反応性に影響を及ぼす可能性があり(Routy et al., 2018)、その概要を図2に示します。腸内細菌叢は、胆汁酸の組成を形成するとともに、抗腫瘍免疫を促進または抑制する免疫経路を活性化させることができます(Jia et al., 2018)。その結果、胆汁酸は、T細胞の活性化や制御性T細胞の増殖といった免疫細胞の機能を調節し得ます。このネットワークが(例えば、ディスバイオシスや胆汁酸プールの変化によって)破綻すると、免疫チェックポイント阻害薬が効果的に作用するための免疫環境が損なわれる可能性があります(Jia et al., 2018)。 |
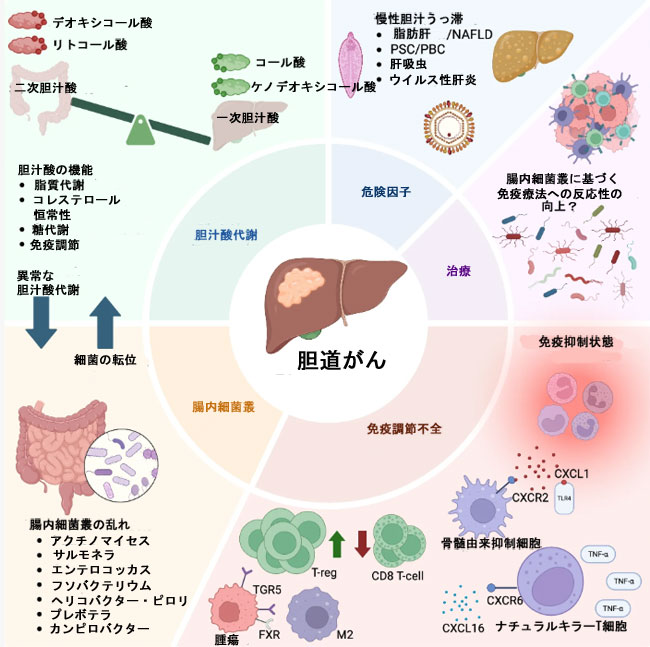 |
図2. 胆道がんにおける胆汁酸代謝、腸内細菌叢、および免疫調節不全の相互作用の概要。 略語:BA、胆汁酸;BTC、胆道がん;CDCA、ケノデオキシコール酸;CA、コール酸;DCA、デオキシコール酸;FXR、ファルネソイドX受容体;LCA、リトコール酸;MDSC、骨髄由来抑制細胞;NK T細胞、ナチュラルキラーT細胞;NAFLD、非アルコール性脂肪肝疾患;PBC、原発性胆汁性胆管炎;PSC、原発性硬化性胆管炎;TNF-α、腫瘍壊死因子α;T-reg、制御性T細胞。 |
| 抗生物質による治療は、腸内細菌叢の多様性や構成を乱し(腸内細菌叢の不均衡:ディスバイオーシス)、がんに対する免疫チェックポイント阻害薬の有効性を低下させる可能性があります(Routy et al., 2018; Pinato et al., 2019)。広域スペクトルの抗生物質は、一次胆汁酸から二次胆汁酸への変換や、免疫調節作用を持つ代謝産物の産生など、重要な代謝機能を変化させることがあります。抗生物質は胆汁酸の生体内変換を担う腸内細菌に影響を及ぼし、一次・二次胆汁酸プールのバランスを変化させるとともに、肝臓における免疫監視機構(例えばCXCL16-NKT細胞シグナル伝達を介したもの)を改変します。こうした変化は、動物モデルにおいて肝腫瘍の生物学的特性と直接関連していることが示されています(Wang et al., 2017)。プロバイオティクスは、胆汁酸組成に影響を与える微生物機能を回復させるのに役立ちます。これには、一次胆汁酸から二次胆汁酸への変換や、肝臓の炎症を制御するファルネソイドX受容体やTakeda Gタンパク質共役受容体5といった受容体の調節などが含まれます(Degirolamo et al., 2014)。これらの経路のバランスを整えることで、特定のプロバイオティクス株(ラクトバチルス属やビフィドバクテリウム属を豊富に含むものなど)は、腸-肝免疫シグナル伝達を改善し、胆道疾患に伴うディスバイオーシス関連の変化の一部を是正できる可能性があります(Degirolamo et al., 2014)。胆汁酸の調節以外にも、プロバイオティクスは腫瘍を促進する炎症を抑制することが示されています。具体的には、IL-6、TNF-α、IL-17などの炎症性サイトカインを減少させると同時に、IL-10やTGF-βなどの抗炎症性メディエーターを増強し、それによってNF-κBを介した発がんシグナル伝達を抑制します(O'Callaghan and Sinderen, 2016)。前臨床がんモデルにおいて、特定のプロバイオティクス株は、樹状細胞の成熟促進、CD8陽性T細胞の浸潤増加、制御性T細胞のバランス調節などを通じて抗腫瘍免疫を増強し、腫瘍の進行を許容しにくい腫瘍微小環境の形成に寄与します(Routy et al., 2018)。さらに、プロバイオティクス関連の細菌群による短鎖脂肪酸(特に酪酸)の産生は、上皮バリア機能を強化し、内毒素血症を軽減し、骨髄由来抑制細胞やマクロファージの分極状態を変化させます。これにより、腫瘍微小環境は免疫寛容ではなく免疫監視が優位な状態へと再構築されます(Louis and Flint, 2017)。メカニズムに関するデータの大部分は結腸直腸がんや肝細胞がんのモデルに由来するものですが、慢性炎症、胆汁酸代謝調節の異常、および免疫回避といった共通の関与が認められることから、これらの経路は胆道がんにおいても生物学的に重要な意義を持っています。 |
| 免疫チェックポイント阻害薬に反応した非小細胞肺がんまたは腎細胞がんの患者から無菌マウスへの糞便微生物叢移植は、マウスにおける抗PD-1抗体の有効性を回復させることが示されていますが、非反応者からの糞便微生物叢移植ではそのような効果は認められません(Routy et al., 2018)。(訳者注:PD-1は、活性化T細胞の表面に存在する免疫抑制受容体です。がん細胞はこの分子を利用して免疫細胞の攻撃から逃れますが、これを阻害する「PD-1抗体薬」ががん免疫療法として広く活用されています)このことは、免疫チェックポイント阻害薬への反応性を媒介する上で特定の微生物成分が極めて重要であることを示唆しており、反応者においてアッカーマンシア・ムシニフィラ、ルミノコッカス・カリダス、ラクノスピラ-GAM79といった分類群が豊富に存在し、それらが転帰の改善や無増悪生存期間の延長と関連しているという臨床的知見とも整合しています(Geva-Zatorsky et al., 2017; Routy et al., 2018)。以下の表2では、ここで取り上げた主要な研究とその主な結果をまとめています。 |
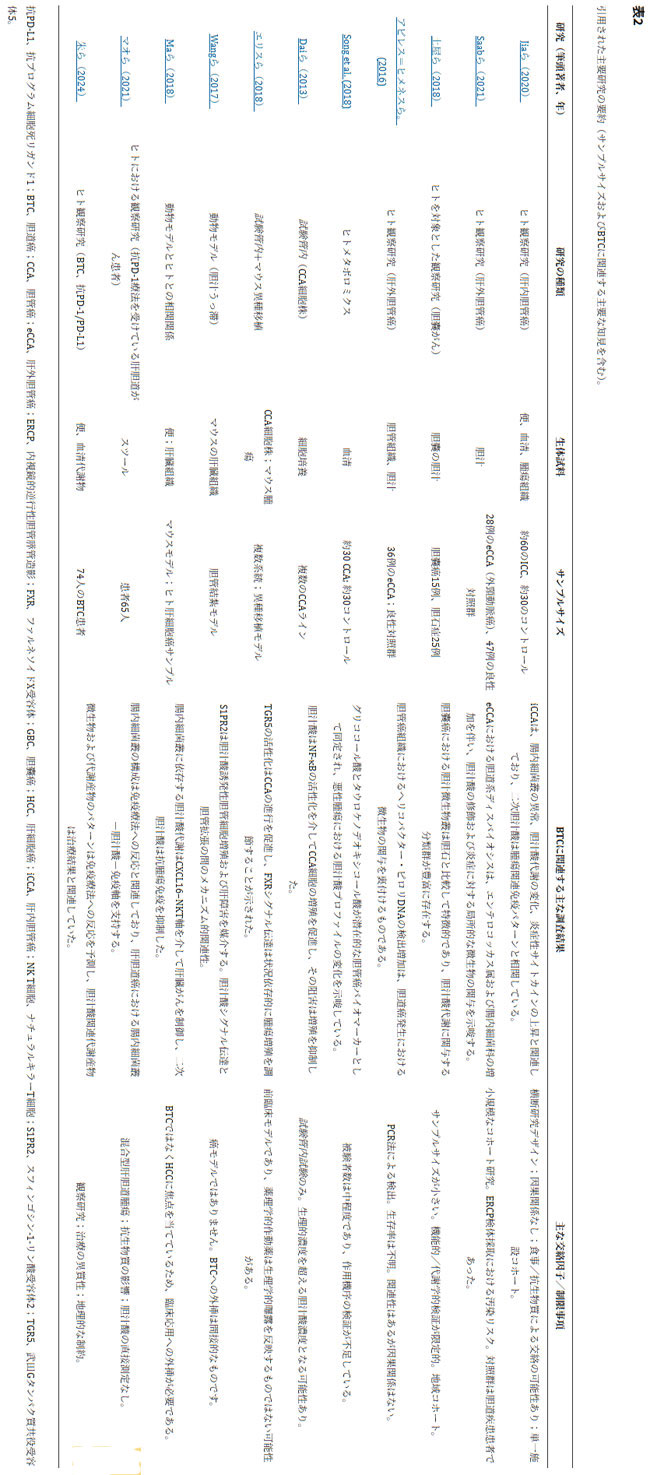 |
| 進行肝胆道がん患者65名を対象とした最近の研究において、抗PD-1療法に対する奏効群と非奏効群の間で、特徴的な微生物叢のパターン(シグネチャー)が認められました。奏効群では、胆汁酸(BA)の調節や免疫活性化に関連する特定の細菌群の増加(濃縮)が確認されました(Lee et al., 2022)。RoutyらやMaoらの研究と同様に、ラクノスピラ科、ルミノコッカス属、エリシペロトリクス科といった細菌群は、肝胆道がんに対する抗PD-1療法の臨床的奏効の向上や抗腫瘍免疫の増強と関連していました(Routy et al., 2018; Mao et al., 2021)。特筆すべき点として、ラクノスピラ科やエリシペロトリクス科は短鎖脂肪酸を産生する細菌であり、これらは抗炎症性サイトカイン(TGF-βやIL-10など)の産生促進や制御性T細胞の分化誘導を介して、粘膜免疫に寄与しています(Rooks and Garrett, 2016; Mao et al., 2021)。こうした粘膜免疫を介した機序は、大腸がんにおいてPD-1阻害療法の有効性を高めることが示されています。また、ラクノスピラ科は一次胆汁酸から二次胆汁酸への変換にも関与しており、腸内恒常性や定着抵抗性を回復させるための糞便微生物叢移植などの微生物叢を標的とした治療において、有益な菌群であると考えられています(Mao et al., 2021)。 |
| 胆道がんにおける免疫療法への反応には、アルファ多様性およびベータ多様性も寄与していると考えられています。アルファ多様性はサンプル内の種の豊富さや多様性を表すのに対し、ベータ多様性は2つ以上の群集間の類似性を比較するものです(Cassol et al., 2025)。PD-1/PD-L1阻害薬の投与を受けた胆道がん患者88名を対象とした前向きコホート研究において、治療開始時のアルファ多様性には、持続的な臨床的ベネフィットが得られた患者群と得られなかった患者群との間で有意差は認められませんでしたが、ベータ多様性においては両群間で明確な分離が示されました。この知見は、免疫チェックポイント阻害薬の有効性を予測する上で、単なる多様性よりも微生物群集の構成の方がより重要である可能性を示唆する、より広範なエビデンスと整合するものです(Fessler et al., 2019; Pinato et al., 2019; Zhu et al., 2024)。 |
| 特筆すべき点として、前述のZhuらによる研究において、持続的な臨床的ベネフィットが認められた群では、バクテロイデーテス門/ファーミキューテス門比が高いことが示されました。このプロファイルは、腎細胞がん、非小細胞肺がん、悪性黒色腫(メラノーマ)など、他の腫瘍型における免疫療法への良好な反応と関連することが以前から報告されています(Fesslerら、2019年;Zhuら、2024年)。この群では、バクテロイデーテス門、とりわけアリスティペス属(A.プトレディニス、A.ディスパー)の菌種が豊富に存在していました。アリスティペス属の菌量が多いことは、全生存期間および無増悪生存期間の延長と相関しており、これは他の悪性腫瘍における知見とも一致しています(Jinら、2019年;Zhuら、2024年)。メカニズムの観点からは、アリスティペス属は、短鎖脂肪酸の産生、炎症経路の調節、および免疫細胞の動員への関与を通じて、抗腫瘍免疫を増強している可能性があります。表3には、ここで取り上げた臨床コホートと、それらと微生物分類群および代謝産物プロファイルとの関連性がまとめられています。 |
| 表3.胆道癌およびその他の固形癌における、腸内細菌叢の分類群、代謝産物プロファイル、およびPD-1/PD-L1標的免疫療法への反応性の間の関連を評価した臨床コホートおよび前臨床モデルの概要 |
 |
| 略語:anti-PD-L1、抗PD-L1(抗プログラム死リガンド1);BA、胆汁酸;DCB、持続的臨床的ベネフィット;FMT、糞便微生物叢移植;NSCLC、非小細胞肺癌;OS、全生存期間;PFS、無増悪生存期間;RCC、腎細胞癌;SCFA、短鎖脂肪酸 |
| これらの結果は、特定のマイクロバイオームの特長(特にアリスティペス属の濃縮やバクテロイデーテス門/ファーミキューテス門比の高値)が、胆道がんにおける免疫チェックポイント阻害薬への反応性を予測し、さらにはその反応性を増強させる可能性があることを示唆しています。プロバイオティクスや糞便微生物叢移植といった介入による腸内細菌叢の調節は、免疫療法の有効性を向上させるための将来的な戦略となり得るでしょう。 |
| 近年の研究により、腸内細菌がフェニルアラニン、チロシン、ロイシンといった標準アミノ酸を胆汁酸に結合させた、新たな種類の微生物修飾胆汁酸の存在が明らかになりました(Garcia et al., 2022)。こうした微生物修飾胆汁酸は、宿主と微生物叢間の相互作用、免疫シグナル伝達、および胆汁酸受容体との相互作用を、未だ完全には解明されていない様式で変化させる可能性があります(Garcia et al., 2022)。 |
| 8.臨床的に一般的な交絡因子 |
| 胆道がんにおける胆汁酸と微生物叢の関連を解釈する際には、胆汁酸組成と微生物群集構造の双方を大きく変化させうる臨床的要因を考慮しなければならない。肝内および肝外の疾患で一般的に見られる胆道閉塞や胆汁うっ滞は、胆管内の胆汁酸濃度を変化させ、疎水性胆汁酸の貯留を増大させ、保護的な「重炭酸塩の傘」を破壊する。これにより、腫瘍の生物学的特性とは無関係に、上皮の損傷や炎症が促進される(Beuers et al., 2010; Chiang, 2013)。内視鏡的または経皮的な胆道ドレナージは、局所の酸素分圧を変化させるとともに手技に伴う汚染を招き、胆道内微生物叢のプロファイルを変動させる可能性がある。胆管炎の発作は、プロテオバクテリアやその他の病原性共生細菌を急激に増殖させる可能性があり、ディスバイオシスが原因なのか、それとも反応性の変化なのかという解釈を複雑にする。プロトンポンプ阻害薬(PPI)やメトホルミンは、それぞれ独立して腸内微生物叢の構成や胆汁酸プールを変化させる一方、ウルソデオキシコール酸は胆汁酸プールの疎水性指数や受容体シグナル伝達環境を直接的に修飾する(Fiorucci et al., 2009; Chiang, 2013)。食事内容、入院の有無、直近の入院歴などもさらなる変動要因となり、微生物多様性の指標や胆汁酸代謝に影響を及ぼす(Rooks and Garrett, 2016; Cassol et al., 2025)。したがって、今後の研究では、胆道への器具操作、直近の胆管炎、抗菌薬やプロトンポンプ阻害薬の使用、血糖降下療法、ウルソデオキシコール酸の使用、腫瘍に対する治療、食事パターン、入院歴について詳細に報告する必要がある。腫瘍に関連するシグナルと治療に伴う変動を切り分けるためには、可能な限り胆道への介入前に行う前向きな検体採取、閉塞状態による層別化、抗菌薬投与との関係における標準化されたタイミングの設定、並行した胆汁酸メタボロミクス解析、そしてα/β多様性解析手法の透明性のある報告が不可欠である。こうした研究デザイン上の配慮を行うことで、再現性が向上し、微生物叢や胆汁酸の変化が臨床的な交絡因子によるものではなく、胆道がんの病態生物学的な変化に起因するものであることをより明確に特定できるようになるだろう。 |
| 9.今後の展望と結論 |
| 腸内細菌叢と胆汁酸の相互作用は、動的かつ双方向的なシステムを形成しており、胆道がんの病因、診断、および治療反応性に多大な影響を及ぼしています。細菌による胆汁酸の変換は、その物理化学的特性やシグナル伝達能を変化させ、抗菌活性、上皮バリア機能、炎症、および免疫監視機構に対して下流の作用を及ぼします。しかし、現在のエビデンスの多くは、横断的研究や不均一な肝胆道系疾患コホートに基づいたものであり、より厳密かつメカニズムに基づいた研究デザインの必要性が浮き彫りになっています。 |
| 胆汁酸の調節不全とがんの進行や治療抵抗性との関連を示す証拠が増えるにつれ、新たな治療の可能性が浮上しています。今後の研究では、処置や閉塞に起因する影響と腫瘍関連のシグナルを切り分けるため、胆道への介入(ステント留置、ドレナージ、抗菌薬投与、化学療法など)の前後における経時的なサンプリングを優先すべきです。各コンパートメント(胆汁・糞便など)に特異的な微生物および胆汁酸のプロファイルと、それらが受容体シグナル伝達経路に及ぼす統合的な影響を明らかにするには、胆汁と糞便のペアサンプルを用いた統合的マルチオミクス解析(メタゲノム解析、標的および非標的メタボロミクス、トランスクリプトミクス)が不可欠となります。抱合状態や一次・二次胆汁酸の種類に基づいて分類されたBAパネルのデータは、胆管上皮細胞、肝細胞、および免疫細胞におけるファルネソイドX受容体、Takeda Gタンパク質共役受容体5、スフィンゴシン-1-リン酸受容体2や関連受容体の活性を示す機能的指標と関連付けられる必要があります。単なる相関関係を超えて因果関係を確立するためには、患者由来オルガノイド、初代胆管上皮細胞モデル、あるいは特定の微生物叢を定着させたノトバイオート(無菌)マウスやヒト化マウスを用いたメカニズムの検証が極めて重要です。 |
| マイクロバイオータ標的療法(プロバイオティクス、選択的抗菌薬、糞便微生物叢移植など)や胆汁酸標的薬(ファルネソイドX受容体作動薬など)は、概念的には魅力的な治療法ですが、胆道がんへの臨床応用にあたっては、その実現可能性とリスクを慎重に評価する必要があります。胆道閉塞、胆管炎、あるいは免疫抑制状態にある患者では、微生物叢への介入が菌血症や敗血症のリスクを高める可能性があります。胆汁酸受容体作動薬は、胆汁酸の種類や腫瘍のサブタイプによって異なる、状況依存的かつ細胞型特異的な作用を及ぼす可能性があります。したがって、初期段階の臨床試験には、安全性の評価項目、胆道感染性合併症の標準化された定義、および作用機序に基づいたバイオマーカー(受容体活性化シグネチャー、胆汁酸組成の変化、免疫プロファイリングなど)を組み込むべきです。さらなる臨床開発を正当化しうる臨床的に意義のある評価項目としては、無増悪生存期間の改善、免疫療法の奏効率向上、胆管炎発症頻度の低下、あるいは検証済みの胆汁酸受容体シグナル伝達経路の再現性ある調節などが挙げられます。 |
| 胆道がんにおいて、微生物叢-胆汁酸軸の調節を、生物学的な妥当性の段階から臨床的に実施可能な治療法へと発展させられるかどうかを判断するには、縦断的なヒトコホート、コンパートメント別のマルチオミクス解析、および機能的モデリングを統合した協調的な戦略が必要となるだろう。 |
参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
この文献は、Front Microbiomes. 2026 May 28;5:1774429.に掲載されたThe interplay between bile acid metabolism and gut microbiome in biliary tract cancers.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |