ケフィアライフをお勧めする中垣技術士事務所




| 文献調査(アロニア:研究の最前線) |
シアニジン3-O-β-ガラクトシドは、 |
Zhuoyan Fan et al., |
|
|
概要 |
| 世界中の老人の間で一般的に起こる代謝障害は、老化の重要な兆候です。神経変性疾患の初期症状には、エネルギー代謝の低下とミトコンドリア機能障害が含まれます。同様に、早期の食事介入は、代謝の調節におけるその役割のために、脳の老化を予防または遅延させるのにより効果的である可能性があります。ポリフェノールの摂取は、アルツハイマー病の予防にその可能性を示しています。ただし、ポリフェノールと老化中の脳のエネルギー代謝との間に密接な関係があるかどうかは不明であります。この研究では、ブラックチョークベリー(Aronia melanocarpa(Michx.)Elliott)のシアニジン3-O-β-ガラクトシドが、老化したマウスの認知機能だけでなく、エネルギー代謝にもプラスの影響を与えるかどうかを評価しようとしました。シアニジン3-O-β-ガラクトシド(25および50mg / kg /日)を8週間胃内投与すると、老化したマウスの脳グルコース取り込みの低下(低下率18.29%対1.05%、7.63%)が効果的に緩和されました。 さらに、シアニジン3-O-β-ガラクトシドは、海馬(神経細胞の数212.33±16.19対285.33±29.53、301.67±10.07; p <0.05)および皮質(神経細胞の数82.00±4.58対111.67±6.51、112.00±1.00; p <0.05)の神経細胞の損傷も軽減しました。 さらに、シアニジン3-O-β-ガラクトシドは脳内のβ-アミロイド負荷を減少させ、モリス水迷路試験で交差プラットフォーム数を有意に増加させました(0.92±1.11対1.83±0.68、2.08±0.58; p <0.05)。さらに、プロテインキナーゼB(AKT)が、脳のエネルギー代謝を制御する上で有益な役割を果たしたシアニジン3-O-β-ガラクトシドの標的である可能性があることを確認しました。これらの結果は、アントシアニンの早期介入が脳のエネルギー代謝の課題の下で神経保護を促進する可能性があることを示唆しました。 |
はじめに |
|
アルツハイマー病(AD)は、世界で最も一般的な老人性神経変性疾患であり、高齢者の健康に対する大きな脅威です。過去10年間、アルツハイマー病の治療に多大な努力とリソースが費やされてきましたが、ほとんどの候補薬は不十分な効果を示したり、最終的に臨床試験に合格しなかったりすることがよくありました(1,2)。 米国食品医薬品局(FDA)によって承認された新しい抗アミロイド薬であるアデュヘルムは高価であり、その臨床的利点をさらに検証する必要があります(3)。さらに、β-アミロイド(Aβ)ベースの治療戦略の有効性は、脳領域の神経細胞がすでに不可逆的に損傷しているなど、アルツハイマー病のさらなる進行を防ぐことができるかどうかに依存します。アミロイドが不可逆的なカスケード反応を引き起こすと、アミロイド沈着物を除去した後でも、この病気は進行し続けます(4)。現在、アルツハイマー病に使用されている4つの主流薬であるリバスチグミン、ガランタミン、ドネペジル、メマンチンは、症状を改善することはできますが、病気の発症を効果的に予防したり、進行を逆転させたりすることはできず、副作用を引き起こす可能性さえあります(5)。 さらに、アルツハイマー病の治療費は2030年までに1兆ドルになると予想されており(6)、これは患者の家族に深刻な負担をもたらす可能性があります。 したがって、アルツハイマー病の予防と治療は世界的な課題となっています。 |
| 異常なエネルギー代謝は老化の重要な兆候です(7)。ブドウ糖は脳の主な燃料であるため、ブドウ糖の消費量の減少は、高齢者だけでなく認知症の患者にも起こります(8,9)。脳のエネルギー代謝は加齢とともに減少します、 これは通常、神経細胞のアポトーシス、神経細胞とシナプスの接続の喪失、ミトコンドリアの機能障害など、神経変性疾患の臨床症状と病理学的特徴の前に発生します(10)。 したがって、脳のエネルギー代謝の障害は、神経変性疾患の病理学において重要な役割を果たす可能性があります。 言い換えれば、それは、特にアルツハイマー病の発生と悪化のための神経変性疾患の開始因子である可能性があります(11,12)。 近年、脳のグルコース代謝は、多くの神経変性疾患の診断のための信頼できるバイオマーカーとなっています(13)。 |
| クリニックでの治療を除いて、食事療法の予防もアルツハイマー病介入の代替戦略でした。 近年、植物由来のポリフェノールは、アルツハイマー病の治療効果が非常に高いため、非常に好まれています。 たとえば、ライチ種子に由来するフラバノールは、タウの過剰リン酸化を有意に抑制し、ラットの認知機能を改善しました(14)。 フェノール酸はアセチルコリンエステラーゼ(AChE)とブチリルコリンエステラーゼ(BChE)を阻害し、β-アミロイド原線維の形成を阻害しました(15)。 認知機能と神経保護のためのスチルベンの利点も広く注目を集めています(16)。 アントシアニンの重要な抗酸化作用と抗炎症作用が広く報告され、確認されています。 最も注目に値するのは、アントシアニンが神経細胞とグリア細胞をβ-アミロイドフィブリル、グルタメート、リポ多糖の損傷から保護することでアルツハイマー病を改善できることです(17)。 以前の報告によると、invivoとinvitroの両方の証拠は、食事性アントシアニンが血液脳関門(BBB)に浸透して脳組織に入り、そこで神経保護活動を行い、認知機能を改善して神経変性疾患を予防できるという事実を裏付けています(.17,18)。 アントシアニンは神経組織に一定期間留まり、脳機能への直接的な影響を増大させる可能性があります(18,19)。 さらに、in vitro研究では、アントシアニンが脳内皮細胞に蓄積し、1時間以内に時間依存的に血液脳関門細胞モデルを通過することが示されました(20,21)。 多くの研究は、β-アミロイド、グルタメートおよびリポ多糖による損傷およびアポトーシスから神経細胞を保護すること、ならびに認知および記憶機能を強化するために酸化ストレスを低減することにおけるアントシアニンの役割に焦点を合わせています。 ただし、アントシアニンとアルツハイマー病の初期代謝との関係は不明なままであり、アルツハイマー病やその他の神経変性疾患の栄養介入としてのアントシアニンの可能性を明らかにするのに役立つ可能性があります。 |
| シアニジン3-O-β-ガラクトシド(Cy3Gal)は、以前の研究でブラックチョークベリー(Aronia. melanocarpa(Michx.)Elliott)から精製されたアントシアニンの一種です(22)。 他のアグリコンと比較して、ガラクトシドに結合したアントシアニンは体内でより速く吸収されます(23)。 さらに、シアニジン3-O-β-ガラクトシドには強力な抗酸化作用と神経保護効果があり、β-アミロイド誘発ラットの学習と記憶障害を改善することがわかりました(22)。 考えられるメカニズムを明らかにするために、シアニジン3-O-β-ガラクトシドが脳のエネルギー代謝を調節し、認知障害をさらに改善できると仮定しました。 この研究では、シアニジン3-O-β-ガラクトシドが脳のエネルギー代謝、臨床症状、および老化促進傾向マウス(SAMP8)における遅発性アルツハイマー病の病理学的特徴に及ぼす影響を調査しました。 シアニジン3-O-β-ガラクトシドはプロテインキナーゼB を標的としている可能性が高く、脳のエネルギー代謝の調節に有益な役割を果たしており、老化したマウスのアルツハイマー病症状を大幅に軽減することがわかりました。 私たちの結果は、シアニジン3-O-β-ガラクトシドの初期の栄養補助食品が、加齢中の脳のエネルギー代謝を改善し、アルツハイマー病の発生率を低下させる可能性があることを示唆しています。 |
材料と方法 |
化学薬品 |
| ブラックチョークベリー(A. melanocarpa(Michx)Elliott)からの純度95%のシアニジン3-O-β-ガラクトシドは、Anhui HaoChen Ecological Agriculture Development Co.,Ltd.(Anhui、China)から提供されました(18). 18F-フルオロデオキシグルコース(18F-FDG)放射性トレーサーおよび18F-フロルベタピル(18 F-AV45)放射性トレーサーは、安徽省馬鞍山バイオテクノロジー株式会社(中国安徽省)から購入しました。 Bax、Bcl-2、切断型カスパーゼ-3、切断型カスパーゼ-9、および西洋ワサビペルオキシダーゼ結合二次抗体は、Cell Signaling Technology(Danvers、MA)から購入しました。 β-アミロイド1‑42 およびHRPヤギ抗ウサギIgG二次抗体は、Abcam Company(Cambridge、UK)から購入しました。 BCAタンパク質濃度アッセイキットは、Shanghai BiYunTian Biotechnology Co.,Ltd.(Shanghai、China)から購入しました。 DABキットは、Dako Denmark A / S(Glostrup、デンマーク)から購入しました。 ヘマトキシリン-エオシン染色キットはBeyotime(北京、中国)から購入しました。 他のすべての化学物質は分析グレードでした。 |
| 実験動物 |
| 生後4ヶ月のオスの老化促進傾向マウス(SAMP8)および老化促進耐性マウス(SAMR1)(体重20±2 g)は、天津伝統中国医学大学(中国、天津)の第一教育病院の動物資源センターから購入しました。 すべてのマウスは、12時間の暗光サイクルの下で温度制御された(22±2℃)動物施設で飼育されました。 適応の1週間後、老化促進傾向マウスを次のようにランダムに3つのグループ(各グループn = 10)に分けました。 モデルグループとしてマウスに0.2mLの滅菌水を経口投与した老化促進傾向マウス。 高用量シアニジン3-O-β-ガラクトシド(Cy3Gal-High)は、0.2 mLの滅菌水(50 mg / kg / day b.w.)に溶解した高用量のシアニジン3-O-β-ガラクトシドをマウスに経口投与しました。 低用量シアニジン3-O-β-ガラクトシド(Cy3Gal-Low)は、0.2 mLの滅菌水(25 mg / kg / day b.w.)に溶解した低用量のシアニジン3-O-β-ガラクトシドをマウスに経口投与しました。 マウスに0.2mLの滅菌水を経口投与した老化促進耐性マウス(SAMR1)グループを対照群とした。 |
| 放射線イメージングは、投与の前後に実行されました。 すべてのマウスは、投与の8週間後に行動試験を受け、最後の行動試験の2週間後に麻酔をかけられました。 すべての実験は、動物倫理福祉委員会(No. NKYYDWLL-2019-105)によって承認されました。 |
| ロータロッドテスト |
| ロータロッドテストは前述のように実行されました(24,25)。 回転ロッドの直径と長さはそれぞれ30mmと60mmでした。 回転ロッドの速度範囲は5〜40rpmでした。 試験の前に、マウスを回転棒上に3分間置いて順応させ、1分間訓練した。 試験では、回転ロッドの速度を4から40rpmまで180秒間加速しました。 テストを4回繰り返し、落下するまでの待ち時間を記録しました。 |
| 放射線画像 |
| マウスを固定具で固定し、6時間の絶食後、尾静脈から約200μCiの18F-フルオロデオキシグルコース(18F-FDG)放射性トレーサーを注射しました。 初期投与量、測定時間、注射時間、注射後の注射器の残留量、および検出時間を正確に記録した。 注射後、マウスを麻酔ボックスに入れ、イソフルランガスを3L /分の流速で5〜10分間与えた。 マウスを固定し、InveonマルチモダリティマイクロPET / SPECT / CTスキャナー(Siemens Medical Solutions)を備えたマイクロポジトロン放出断層撮影/コンピューター断層撮影(PET / CT)に配置しました。 動的PETモニタリングとCTスキャンがマウスで実行されました。 スキャン中、イソフルランガスが継続的に注入されました。 脳内の放射性物質の取り込み値を記録し、標準取り込み値(SUV)をInveon Research Workplaceソフトウェア(Siemens)で分析しました。 PET / CT画像は、PMODソフトウェア(バージョン3.4、PMOD Technologies Ltd.,スイス、チューリッヒ)を使用して処理されました。 |
| β-アミロイドのモニタリング方法はブドウ糖のそれと類似しています。 つまり、約100μCiの18F-フロルベタピル(18F-AV45)を尾静脈からマウスに注射し、PET / CTでスキャンしました。 |
| ニッスル染色 |
| 脳組織のニッスル染色は以前に記載されたように実施された(25)。 マウスを安楽死させ、通常の生理食塩水で灌流した。 次に、脳が見つかり、4%パラホルムアルデヒドを入れました。 クリオスタット(LeicaMicrosystems、Wetzlar、Germany)を使用して、25μmの厚さの連続冠状切片を切断した。 切片を0.01M PBSで5分間2回洗浄し、1%トルイジンブルー染色液で20〜25°Cで5〜10分間インキュベートしました。 次に、切片を蒸留水ですすぎ、95%エタノールに30分間浸し、100%エタノールで脱水した。 脱水後、切片をキシレンに入れ、樹脂媒体を使用してカバースリップをかけた。 海馬と大脳皮質の神経細胞は、Image J-Pro Plusソフトウェア(NIHのWayne Rasbandによって開発された)を使用して定量化されました。 |
| ウエスタンブロット |
| マウスの脳はすぐに液体窒素に入れられた。 凍結した脳をタンパク質溶解物バッファーでホモジナイズし、4℃で15分間12000gで遠心分離しました。 Bio-Radタンパク質アッセイ(Beyotime、北京、中国)を使用して、タンパク質濃度を決定しました。 タンパク質サンプルをSDSPAGEで分離し、ポリ(フッ化ビニリデン)(PVDF)メンブレンに転写し、5%スキムミルクで2時間密封しました。 次に、PVDFメンブレンを特定の一次抗体とともに4℃でインキュベートしました。 西洋ワサビペルオキシダーゼ結合二次抗体をPVDFメンブレンとインキュベートしました。 タンパク質は化学発光試薬によって視覚化され、ゲル記録システム(GelDoc It 310イメージングシステム)によって分析されました。 |
| モリス水迷路テスト |
| シアニジン3-O-β-ガラクトシドを胃内投与して8週間後、前述のようにモリス水迷路(MWM)テストを実施しました(25)。 円形のプールは直径120cm、深さ40cmで、プラットフォームは見えません(北京中石技術開発有限会社、北京、中国)。 プールの水は黒インクと混合されました。 実験中、水の温度は22〜24°Cのままで、迷路の周りのすべてのランドマークは同じままでした。 モリス水迷路テストには、空間学習と習得の試行(隠しプラットフォームの試行)と空間プローブの試行が含まれていました。 空間取得試行の前に、各マウスを水中に入れて2分間順応させ、第1象限の中央に1日間配置されたプラットフォームが表示されました。 次に、水面から1cm下のプラットフォームでプールに水を追加しました。 隠されたプラットフォームの試行中、各マウスは、壁に面して、開始位置の1つから水中に配置されました。 空間取得試行は、1日4回の試行で5日間にわたって実施されました。 動物がプラットホームに到着した場合、タイマーは停止されました。 動物が90秒以内にプラットフォームを見つけられなかった場合、プラットフォームの位置を知るために15秒間プラットフォームに置かれました。 空間プローブテストは7日目に実施され、プラットフォームは取り外されました。 次に、マウスを、プラットフォームを備えた象限の反対側の象限から水中に入れた。 遊泳経路、対象象限での時間、およびマウスがプラットフォームを横切った時間を記録した。 試験後、マウスをタオルで乾かして保温した。 テストはビデオカメラで記録され、データは任意の迷路行動追跡ソフトウェア(Stoelting Co.、Wood Dale、IL)を使用して分析されました。 |
| 営巣動作 |
| 営巣テスト(ネスティング)は、Deacon etal.26によって以前に説明されたように実行されました。 手短に言えば、新しい木の削りくずをマウスケージに入れて、32枚の柔らかい紙(4.5cm×4.5cm)を置いて底を平らにしました。 24時間後、マウスの営巣能力を評価し、スコアを付けました。 |
| 免疫組織化学的およびヘマトキシリン-エオシン染色(HE染色) |
| マウスを安楽死させ、通常の生理食塩水で灌流した。 次に、脳を見つけて、4%中性パラホルムアルデヒドに24時間入れました。 パラフィン切片をキシレンで脱ロウし、分化するまでエタノールの異なる勾配で脱水しました。 合計0.01mol / Lのクエン酸バッファーをスライスに入れ、抗原修復のために電子レンジで加熱しました。 ヤギ血清ブロッキング溶液を20分間使用した。 β-アミロイド1-42抗体希釈液(1:100)を添加し、4℃で1晩インキュベートしました。 HRPヤギ抗ウサギIgG二次抗体(1:2000)を添加し、37°Cで30分間インキュベートしました。 37°Cで30分間インキュベートしました。 ADABキットを5〜20分間追加しました。 核はヘマトキシリンでわずかに染色された。 切片をスキャンし、アミロイドタンパク質の沈着が顕微鏡下で茶色であることが観察された。 統計分析には、Image J-Pro Plusソフトウェア(NIHのWayne Rasbandによって開発された)を使用しました。 組織学的検査のために、脳組織をパラフィンに包埋し、ヘマトキシリン-エオシン(HE)で染色しました。 |
| 分子動力学シミュレーション |
| プロテインキナーゼBタンパク質(PDB ID:4GV1)およびホスファチジルイノシトール3-キナーゼ(PI3K)タンパク質(PDB ID:3HHM)の構造は、PDBデータベースからのものです。 シアニジン3-O-β-ガラクトシドの構造はPubChem(CID:441699)からのものでした。 Autodock Vinaは、タンパク質とシアニジン3-0-β-ガラクトシド複合体のコンフォメーションの分子ドッキングに使用されました(http://autodock.scripps.edu)。 ボックスの分子特性は、HARvard Macromolecule Mechanics 36(CHARMM 36)のChemistryを使用して計算およびシミュレーションされました。 描画にはPyMOLを使用しました(http://www.pymol.org/)。 水素結合と疎水性相互作用はligplot +によって分析されました。 |
| 統計分析 |
| SUVmeanは、すべての関心領域(ROI)から自動的に抽出され、小脳を参照領域として使用して標準化取込値比(SUVR)を計算するために使用されました(SUVR = SUVmean /小脳SUVmean)。 |
| データは平均±標準偏差として表されます。 テストデータは、SPSSバージョン20.0ソフトウェアパッケージを使用した一元配置分散分析によって処理されました。 データの多重比較は、p <0.05とp <0.01で有意差を計算するために使用されたテューキーの検定を介して実行されました。 |
結果 |
シアニジン3-O-β-ガラクトシドは、老齢マウスの脳におけるエネルギー代謝を改善しました |
| 脳のエネルギー代謝におけるシアニジン3-O-β-ガラクトシドの効果を調査するために、老齢マウスの強制運動持久力を回転バーでテストして、エネルギー消費量を分析しました。 老化促進耐性マウス(SAMR1)と比較して、老化促進傾向マウス(SAMP8)の運動能力は著しく低下した。 8週間の介入後、高用量シアニジン3-O-β-ガラクトシド(Cy3Gal-High)による治療は、低用量シアニジン3-O-β-ガラクトシド(Cy3Gal-Low)による治療よりも、運動持久力の改善に有意な効果をもたらしました。 まとめると、シアニジン3-O-β-ガラクトシドはエネルギー代謝を改善し、老齢マウスの運動能力を促進しました(補足図1)。 |
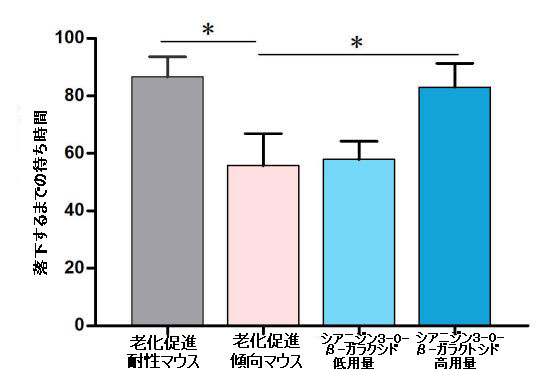 |
| 補足図1 シアニジン3-O-β-ガラクトシドは、老齢マウスの強制運動持久力を改善しました(n = 8)。 加速するロータロッドからのマウスの落下までの潜時を試験した。* p <0.05。 |
| ATP(訳者注:アデノシン三リン酸)は、その95%以上がグルコース代謝によって生成され、脳のエネルギー代謝の主要な通貨です(27)。上に示したシアニジン3-O-β-ガラクトシドの有益な効果を考慮して、我々は老齢マウスの脳におけるグルコース代謝を動的にモニターした。PET / CTテクノロジーは、悪性疾患の転移を特定して特定するために一般的に使用されています。PET / CTを適用すると、病変の機能や代謝などの詳細な分子情報を高感度、正確、具体的に提供できます(28)。 我々は、PET / CTによってマウス脳のさまざまな領域のグルコース代謝に対するシアニジン3-O-β-ガラクトシドの効果を調べ、18F-フルオロデオキシグルコースの標準化取込値比によってグルコース代謝を定量化しました(図1A)。 8週間後、標準化取込値比は老化促進耐性マウスで有意差はありませんでしたが(補足図2および3)、老化促進傾向マウスのほぼすべての脳領域で明らかに減少しました。これは、グルコース代謝が加齢とともに有意に減少したことを意味します(図1B、Cおよび補足図2および3)。 |
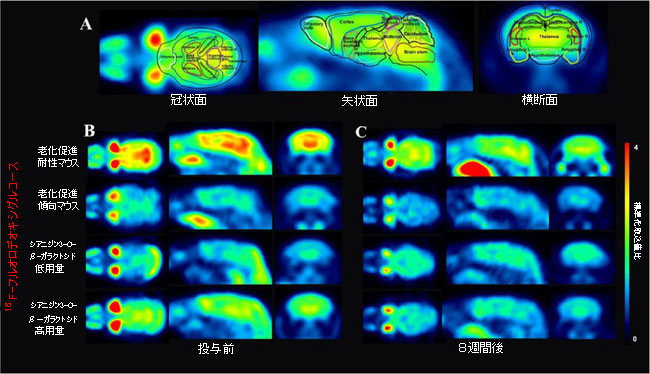 |
図1 シアニジン3-O-β-ガラクトシドは、老齢マウスの脳におけるグルコース取り込みを改善しました(n = 8)。 (A)18F-フルオロデオキシグルコースのPETスキャンによって視覚化されたマウス脳のさまざまな領域の画像(上のパネル)。 シアニジン3-O-β-ガラクトシドの投与前(B)および8週間後(C)の異なるグループのマウス脳におけるグルコース代謝の代表的な放射線画像。 さまざまなグループのマウスの脳における18F-フルオロデオキシグルコースの取り込みは、標準化取込値の比率(SUVR)を測定することによって評価され、比率が高いほど18F-フルオロデオキシグルコースの取り込みが多くなります。 |
 |
 |
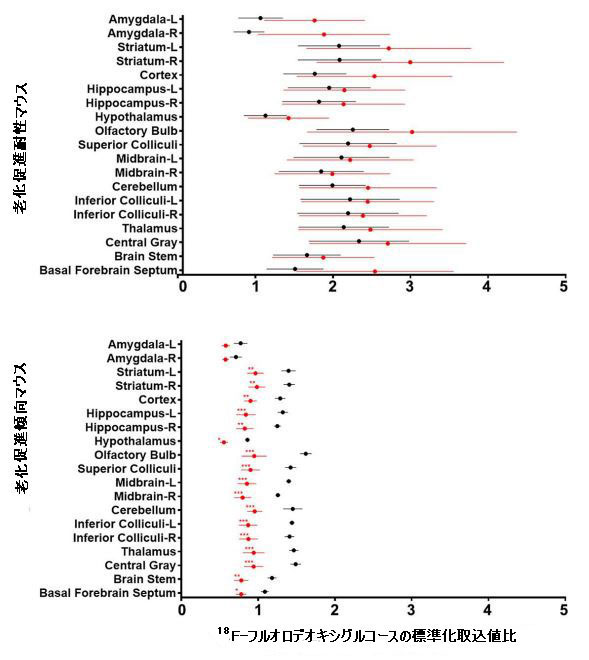
| 補足図2 18F-フルオロデオキシグルコースの標準化取込値の比率は、マウスの脳のさまざまな領域で定量化されました(n = 8)。 ベースライン値(シアニジン3-O-β-ガラクトシドの投与前)は黒で示され、8週間の投与後の値は赤で示されました。 * p <0.05、** p <0.01、*** p <0.001。 |
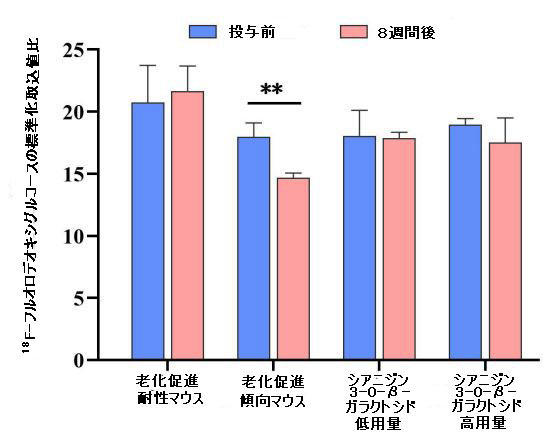 |
| 補足図3 異なるグループのマウスの脳における18F-フルオロデオキシグルコースの総取り込み(n = 8)。 * p <0.05、** p <0.01。 |
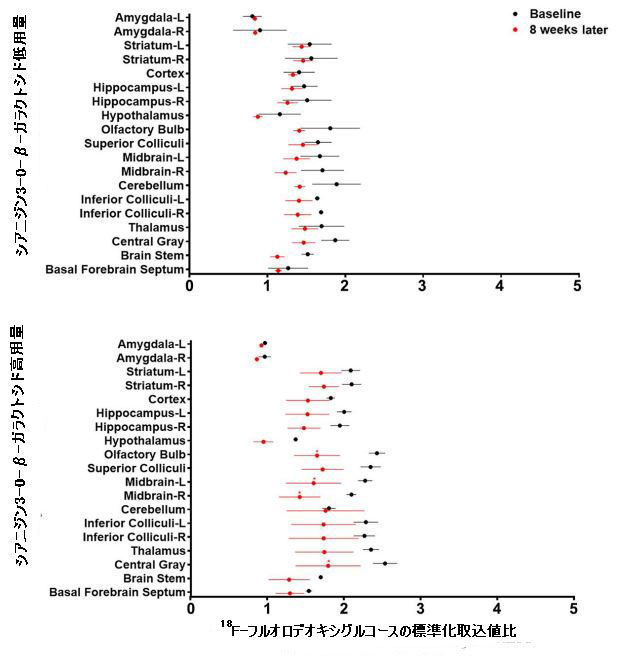
| 比較すると、低用量シアニジン3-O-β-ガラクトシドグループのマウスの脳における18F-フルオロデオキシグルコースの取り込みは、特に海馬、大脳皮質、嗅球、線条体、上丘、下丘、中脳、小脳、視床、中枢灰色物質および脳幹領域で減少しませんでした(補足図2および3)。 高用量のシアニジン3-O-β-ガラクトシドで治療されたマウスでは、脳の総グルコース取り込みに有意な増加は見られませんでした(補足図3)。 嗅球、中脳、および中央灰白質領域での18F-フルオロデオキシグルコースの取り込みが少ないと、この現象が発生する可能性があります(補足図2)。 これにより、高用量のシアニジン3-O-β-ガラクトシドで治療されたマウスの総脳グルコース摂取量に有意な改善が見られないという現象が生じる可能性があります(補足図3)。 |
| 全体として、シアニジン3-O-β-ガラクトシド介入と脳のグルコース取り込みの間には正の相関関係がありました(補足図4)。 シアニジン3-O-β-ガラクトシドの8週間の介入は、老化中のマウスの脳のエネルギー代謝に有益な効果をもたらしました。 |
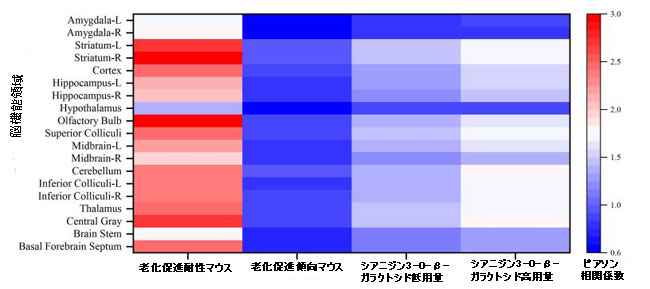 |
補足図4 マウス脳のさまざまな機能領域における18F-フルオロデオキシグルコースの標準化取込値の比率とさまざまな治療法との相関分析。 値が高いほど、相関が強くなります。 |
| シアニジン3-O-β-ガラクトシドは老齢マウスの神経損傷を軽減しました |
| アルツハイマー病などの神経変性疾患は、海馬や大脳皮質の神経損傷に直接関係していることが一般的に知られています(29)。 したがって、シアニジン3-O-β-ガラクトシドが老齢マウスの神経細胞に保護効果を及ぼすかどうかをさらに調査しました。 図2に示すように、ヘマトキシリン-エオシン染色の結果は、図2Aの赤い矢印で示されているように、老化促進耐性マウスの海馬の神経細胞が明確な核でしっかりと整然と配置されていることを示しました。 しかし、老化促進傾向マウスでは、神経細胞はより無秩序な配置を採用し、神経細胞核のクロマチンは高密度で凝縮しており、アポトーシスと多数の神経細胞の喪失を伴う核濃縮を示しています(赤い矢印で示されています)(図2Aおよび補足図5)。 シアニジン3-O-β-ガラクトシドを老化促進傾向マウスに投与した後、図2AのCA1およびDG領域で観察されるように、海馬の神経細胞は円形(または楕円形)で、水色の核でしっかりと配置されていました(赤い矢印で示されています)。 |
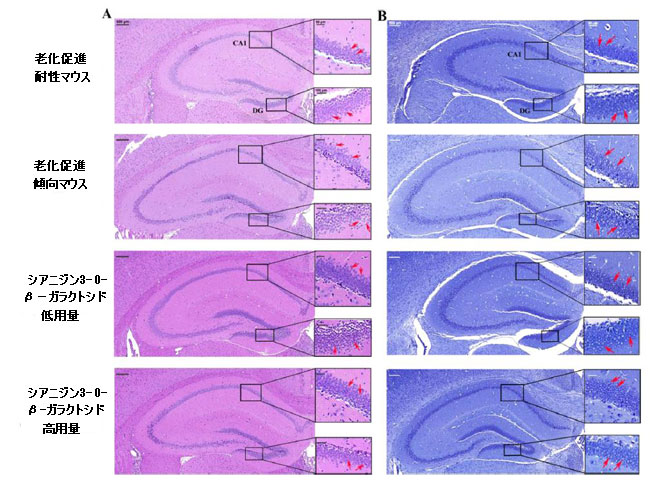 |
| 図2 マウスの脳の海馬領域の組織学的検査(n = 6)。 異なるグループのマウスにおける海馬領域のヘマトキシリン-エオシン染色(A)およびニッスル染色(B)の代表的な画像。海馬CA1(右上の長方形)および歯状回(DG、右下の長方形)領域を調べた。 赤い矢印はニューロンを示しています。 |
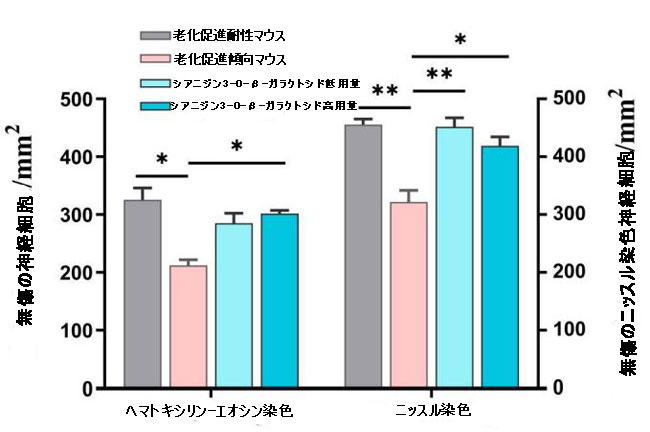 |
補足図5 海馬の無傷の神経細胞の数を示すヒストグラム(n = 3)。 * p <0.05、** p <0.01。 |
| シアニジン3-O-β-ガラクトシドは、老化促進傾向マウスの海馬における神経細胞喪失の顕著な減少を改善しました(補足図5)。 ニッスル染色の結果は、ヘマトキシリン-エオシン染色の結果と同様でした。神経細胞は、明確な円形または楕円形の核および核小体を伴う正常な形態を示し、老化促進耐性マウスの細胞質に透明なニッスル体を示した。 比較すると、老化促進傾向マウスでは、核が損傷し、細胞質が収縮した多数の萎縮性神経細胞が観察され、ほとんどの神経細胞がニッスル体の喪失を示しました(赤い矢印で示されています)。 シアニジン3-O-β-ガラクトシドは、神経細胞の形態(CA1およびDG領域に表示されたものなど)を改善し、加齢による神経細胞の喪失を著しく逆転させましたが、効果は用量に依存しませんでした(図2Bおよび補足図5)。 |
| ヘマトキシリン-エオシン染色およびニッスル染色により、老化促進傾向マウスの大脳皮質でも喪失を伴う神経変性に対するシアニジン3-O-β-ガラクトシドの改善が観察されました(補足図6および7)。 これらの結果は、シアニジン3-O-β-ガラクトシドが老齢マウスの神経細胞の損傷とアポトーシスを軽減することによって潜在的な神経保護効果を持っていることを示唆しました。 |
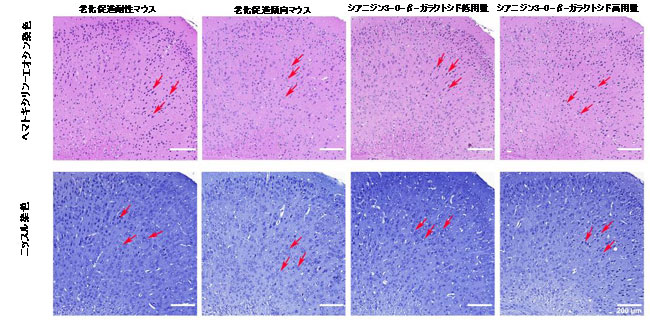 |
| 補足図6 マウスの脳における大脳皮質の組織学的検査(n = 6)。 異なるグループのマウスの大脳皮質領域のヘマトキシリン-エオシン染色(A)およびニッスル染色(B)の代表的な画像。赤い矢印は神経細胞を示しています。 |
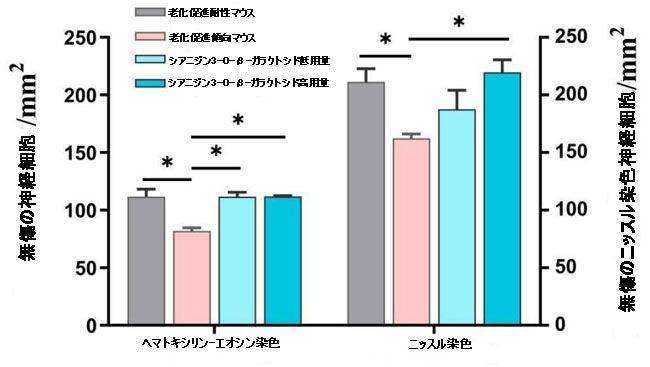 |
| 補足図7 皮質内の無傷の神経細胞の数を示すヒストグラム(n = 3)。 * p <0.05。 |
| シアニジン3-O-β-ガラクトシドはミトコンドリア機能と神経細胞アポトーシスの主要な調節因子を調節しました |
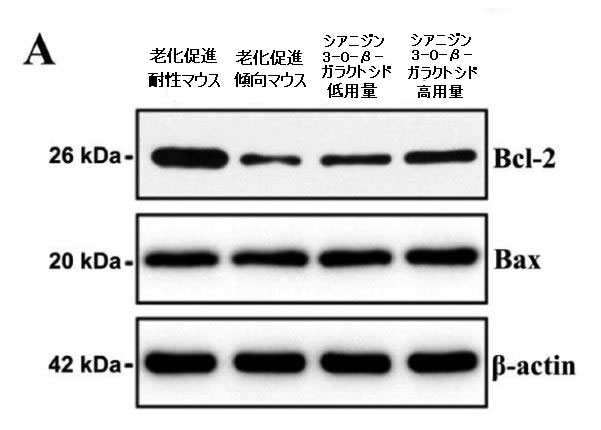
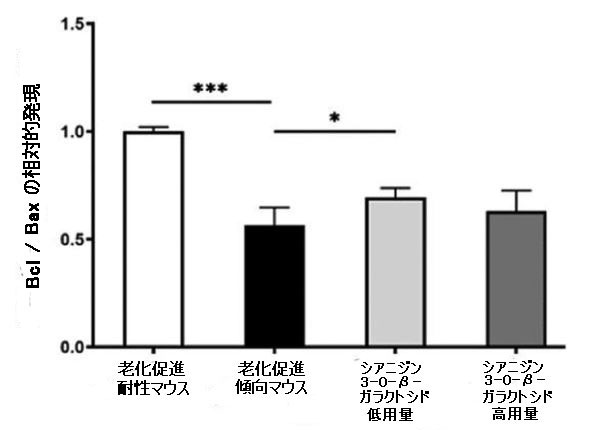
| ミトコンドリアは生命にエネルギーを供給する発電所であり、その構造と機能はエネルギー代謝と密接に関連しています。 同時に、ミトコンドリアは脊椎動物のアポトーシスの中心的な調節因子でもあります。 エネルギー代謝と神経細胞に対するシアニジン3-O-β-ガラクトシドの有益な効果を考慮して、最初にミトコンドリア膜タンパク質、B細胞リンパ腫-2(Bcl-2)およびBcl-2関連X(Bax)を調べました。 老化促進傾向マウスでの脳では、老化促進耐性マウスと比較してBcl-2とBaxの発現率が著しく低下しました。 低用量のシアニジン3-O-β-ガラクトシドを8週間投与した後、老化促進傾向マウスにおけるBcl-2とBaxの発現比はアップレギュレーションされましたが、高用量のシアニジン3-O-β-ガラクトシドで治療されたマウスでは有意な効果は観察されませんでした(図3A)。 このように、シアニジン3-O-β-ガラクトシドは老齢マウスの脳のミトコンドリア膜透過性をある程度改善しました。 |
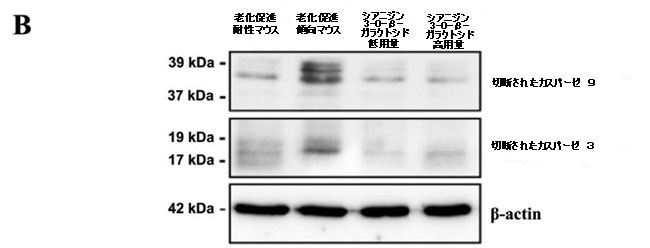
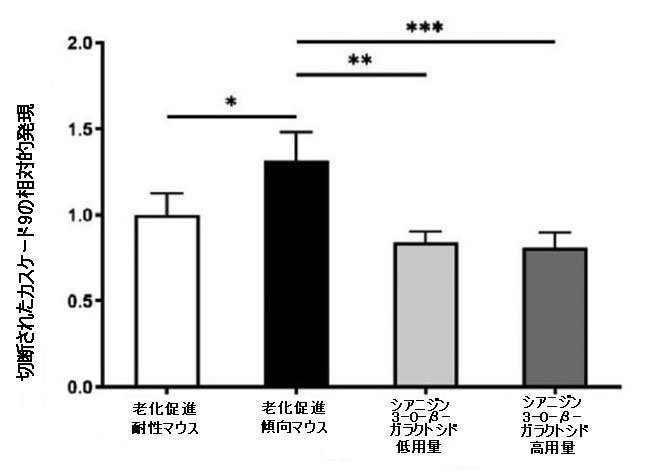
| カスパーゼは、ミトコンドリア依存性アポトーシス経路の重要な酵素であり、カスパーゼ-9とカスパーゼ-3がアポトーシスの過程における主要なエフェクターです(30)。 抗アポトーシスタンパク質Bcl-2とアポトーシス促進タンパク質Baxの間の相互作用は、ミトコンドリア外膜の透過性を調節し、したがって、カスパーゼ-9およびカスパーゼ-3依存性経路を介してアポトーシスを調節することができます。 シアニジン3-O-β-ガラクトシドはカスパーゼ-9とカスパーゼ-3の活性化を有意に阻害しましたが、老化促進傾向マウスの脳では明らかに活性化されていました。 図3Bに示すように、シアニジン3-O-β-ガラクトシドの投与後、切断されたカスパーゼ-9および切断されたカスパーゼ-3の発現は大幅に減少しました。 全体として、シアニジン3-O-β-ガラクトシドは、老齢マウスのミトコンドリア経路が関与する神経細胞のアポトーシスを阻害する可能性を示しました。 結果は、組織学的検査で示された神経保護効果と一致していました(図2および補足図6)。 |
 |
 |
 |
 |
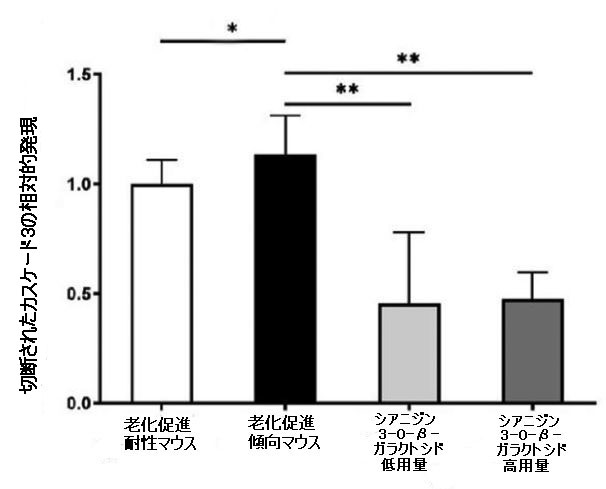 |
図3.老齢マウス(n = 4)のミトコンドリア機能と神経細胞アポトーシスに対するシアニジン3-O-β-ガラクトシドの効果。 代表的なウエスタンブロット画像とBcl-2およびBax(A)、切断されたカスパーゼ-9、および切断されたカスパーゼ-3(B)の分析。 * p <0.05、** p <0.01、および*** p <0.001。 |
| シアニジン3-O-β-ガラクトシドは、老齢マウスの認知能力と記憶能力を改善しました |
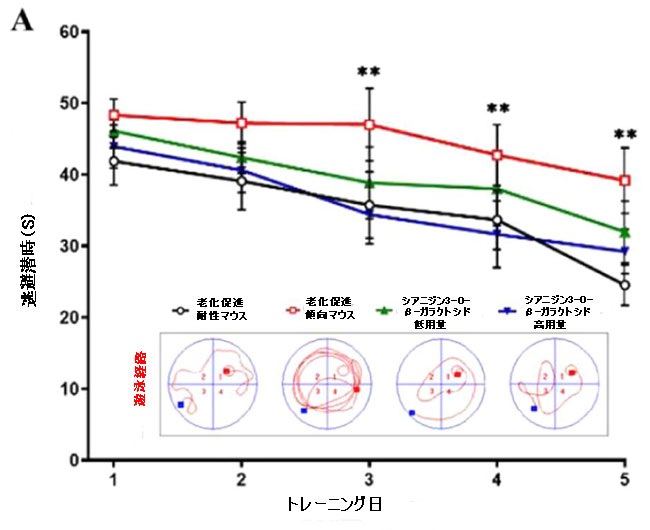
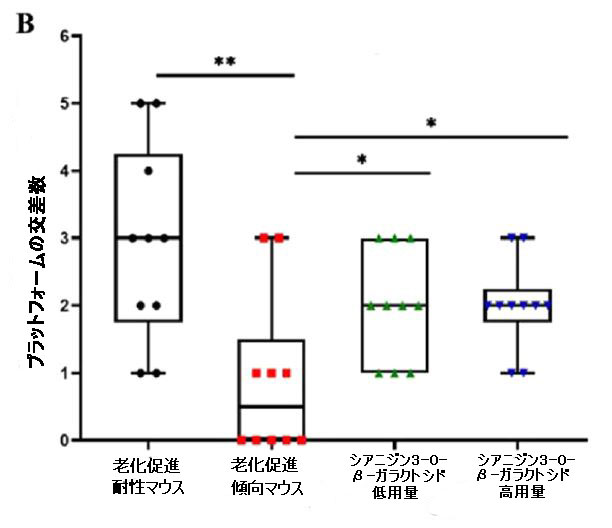
| シアニジン3-O-β-ガラクトシドによるエネルギー代謝の改善がアルツハイマー病の遅延に役立つかどうかを調べるために、モリス水迷路テストを実行して、老齢マウスの空間記憶と行動能力を評価しました。 空間取得テストでは、安全なプラットフォームを探すための逃避潜時は訓練によって徐々に減少しました。 明らかに、老化促進傾向マウスと老化促進耐性マウスの違いは、図4Aに示すように、時間の経過とともに増加しました。これは、代謝障害と神経損傷によって引き起こされる可能性があります。 老化促進傾向マウスは、老化促進耐性マウスと比較して加齢に伴う学習能力を示しました。 興味深いことに、シアニジン3-O-β-ガラクトシドの摂取により、老化促進耐性マウスと未処理の老化促進傾向マウスの間のギャップが狭まりました。 空間取得テストの結果と同様に、シアニジン3-O-β-ガラクトシドも老化促進傾向マウスが安全なプラットフォームを通過する時間を大幅に延長しました(図4B)。 結果は、シアニジン3-O-β-ガラクトシドが老齢マウスの空間学習と記憶障害に保護効果を持っていることを示しました。 |
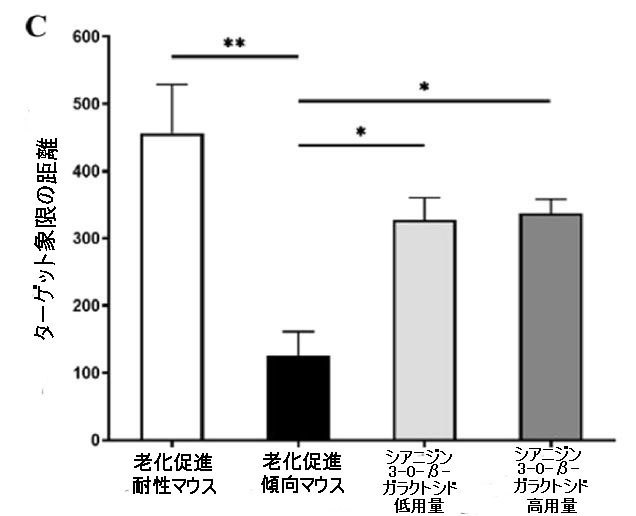
| 特に、シアニジン3-O-β-ガラクトシドは老化促進傾向マウスの総遊泳距離と速度を増加させ(図4C、D)、これは回転ロッドテストでの運動持久力の改善と一致していました(補足図1)。 営巣行動は、アルツハイマー病の病理学的過程における認知機能障害の指標として使用できます(32)。 営巣試験では、老化促進耐性マウスの営巣の平均スコアは3.83±0.40(合計スコアは5)であり、マウスは紙を使用して、巣の反対側に糞便が集中したきれいなボウル型の巣を構築した。対照的に、老化促進傾向マウスのスコアは1.83±0.75のみでした。老化促進傾向マウスは、シアニジン3-O-β-ガラクトシドの低用量と高用量の投与後に、紙が巣の周りに散らばったカップ型の巣を作ることができ、マウスのスコアはそれぞれ3.00±0.63と2.83±0.98でした(補足図8)。 これらの結果は、シアニジン3-O-β-ガラクトシドが老化したマウスの行動能力を効果的に改善し、認知機能障害を改善する可能性があることを示しています。 |
 |
 |
 |
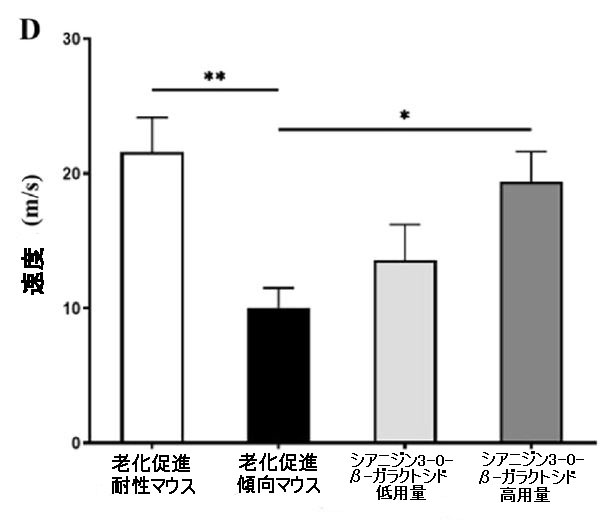 |
| 図4 老齢マウスの空間学習および記憶能力に対するシアニジン3-O-β-ガラクトシドの効果(n = 6) |
| 5日間にわたる空間取得試験で安全なプラットフォームを探しているマウスの逃避潜時。 (A)の挿入図は、モリス水迷路試験(A)におけるさまざまなグループのマウスの代表的なアクショントラックを示しています。 タイムズマウスは、空間プローブテストでプラットフォームを通過しました(B)。 空間プローブテスト(C)でのターゲット象限の合計移動距離。 空間プローブテスト(D)におけるマウスの遊泳速度(m / s)。 ** p <0.01および* p <0.05。 |
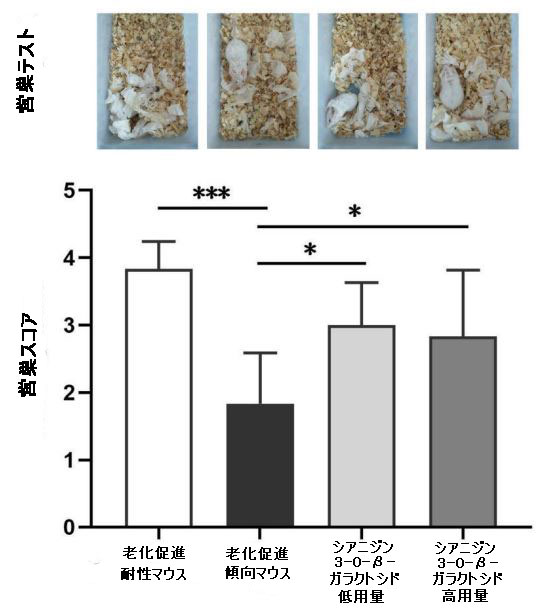 |
| 補足図8 老齢マウスの営巣行動能力に対するシアニジン3-O-β-ガラクトシドの効果(n = 6) |
| 異なるグループのマウスの営巣行動の代表的な写真が示され(上のパネル)、営巣スコアが分析されました(下のパネル)。 *** p <0.001。 |
| シアニジン3-O-β-ガラクトシドは老齢マウスの脳におけるβ-アミロイド蓄積を減少させました |
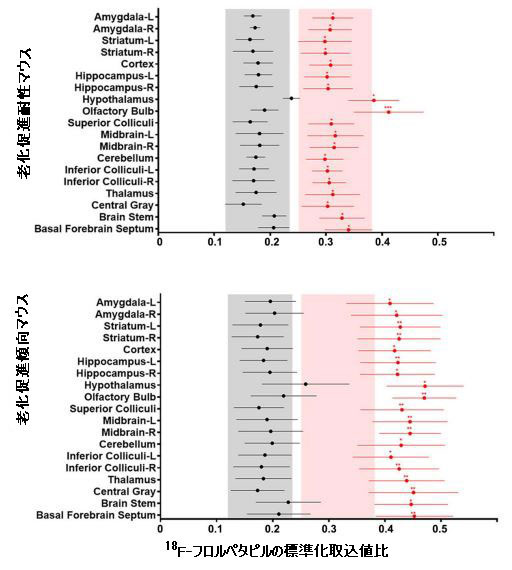
| 中枢神経系におけるβ-アミロイドの異常な蓄積は、アルツハイマー病の重要な兆候の1つでした(18)。 β-アミロイドとの親和性、感度、特異性が高い18F-フロルペタピル(18F-AV45) PETを使用して、β-アミロイド負荷をマークしました(33)。 脳画像解析により、PET画像を視覚的に分析してマウスの各脳領域における放射性18F-フロルペタピルの分布を評価し、18F-フロルペタピルの標準化取込値比を計算してβ-アミロイド負荷の値を評価しました。 18F-フロルペタピルの沈着が老化促進傾向マウスマウスの脳で著しく増加した8週間後、少量の18F-フロルペタピルが脳の老化促進耐性マウスに沈着しました(図5B)。 老化促進傾向マウスでは、β-アミロイド負荷は海馬、前脳基底部、嗅球、線条体、中脳、上丘、視床下部、視床、および中央灰白質で増加しましたが、β-アミロイドは主に老化促進耐性マウスの嗅球領域に蓄積しました(補足図9)。 補足図9に示すように、低用量および高用量のシアニジン3-O-β-ガラクトシドは、β-アミロイド負荷の有意な改善効果を示しました(図5B)。 シアニジン3-O-β-ガラクトシドは、老化促進傾向マウスの脳におけるβ-アミロイド負荷の範囲と程度を大幅に減少させました。 結果は、異なるグループのマウス脳組織の免疫組織化学分析によって検証されました。 老化促進傾向マウスの海馬(補足図10A)および大脳皮質(補足図10B)に大量のプラークが蓄積し、シアニジン3-O-β-ガラクトシド投与後にプラークのレベルが低下しました。 結果は、シアニジン3-O-β-ガラクトシドの早期介入が、加齢中のマウス脳におけるアルツハイマー病に関連する病理学的進行を遅らせることを示唆しました。 |
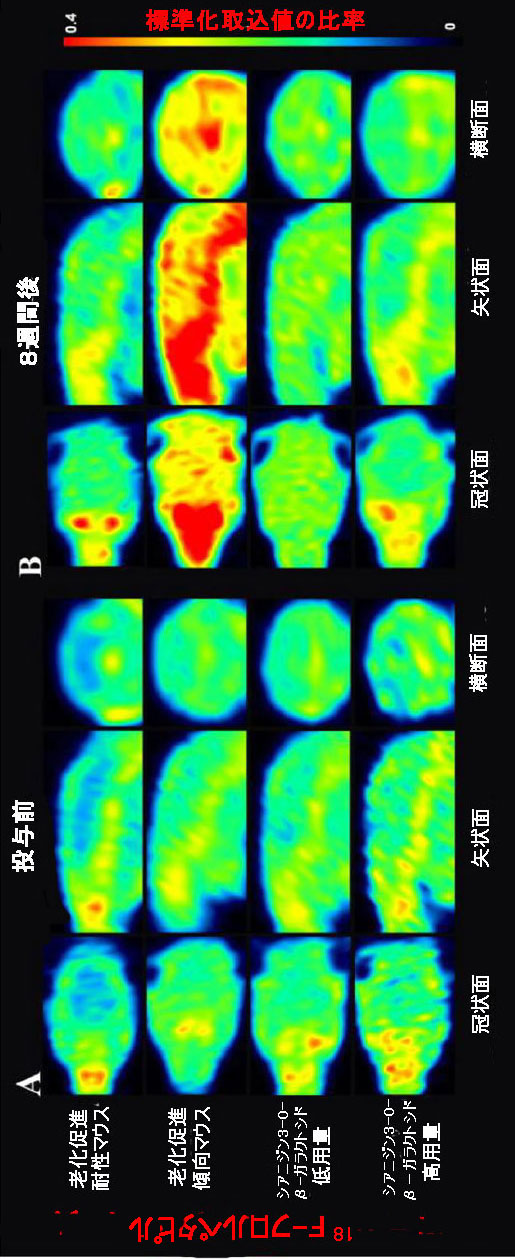 |
| 図5 老齢マウスの脳におけるβ-アミロイド蓄積に対するシアニジン3-O-β-ガラクトシドの効果 |
| シアニジン3-O-β-ガラクトシドの8週間投与前(A)および投与後(B)の異なるグループのマウス脳における18F-フロルペタピル(18F-AV45)取り込みの代表的な放射線画像。 異なるグループのマウス脳における18F-フロルペタピルの取り込みは、標準化取込値の比率を測定することによって評価されました。 比率が高いほど、18F-フロルペタピルの取り込みが高くなります。 |
 |
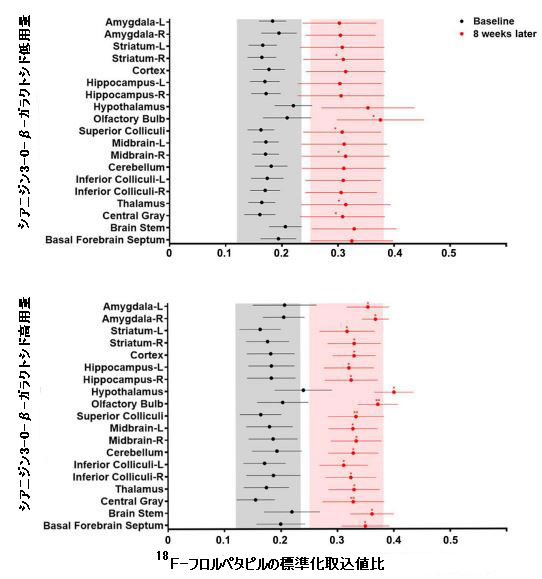 |
| 補足図9。マウス脳のさまざまな領域における18F-フロルペタピル標準化取込値比の定量化(n = 8) |
| ベースライン値(シアニジン3-0-β-ガラクトシドの投与前)は黒で示され、8週間の投与後の値は赤で示されました。* p <0.05、** p <0.01、*** p <0.001。 |
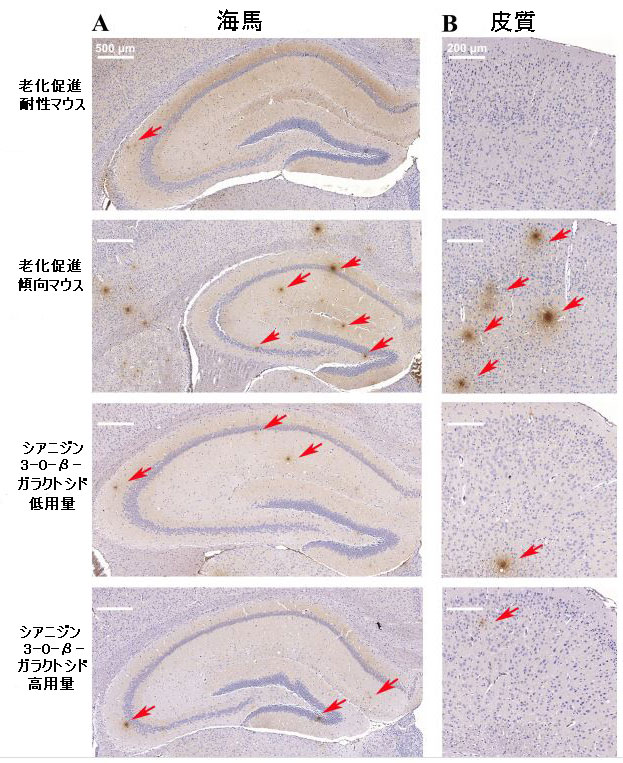 |
| 補足図10 マウス脳のβ-アミロイドを標的とする免疫組織化学的染色(n = 4) |
| シアニジン3-O-β-ガラクトシドを8週間投与した後のマウスの海馬(A)および皮質(B)領域におけるβ-アミロイドのレベル。赤い矢印はβ-アミロイド凝集体を示しています。 |
| シアニジン3-O-β-ガラクトシドはプロテインキナーゼBファクターをターゲットにしている可能性があります |
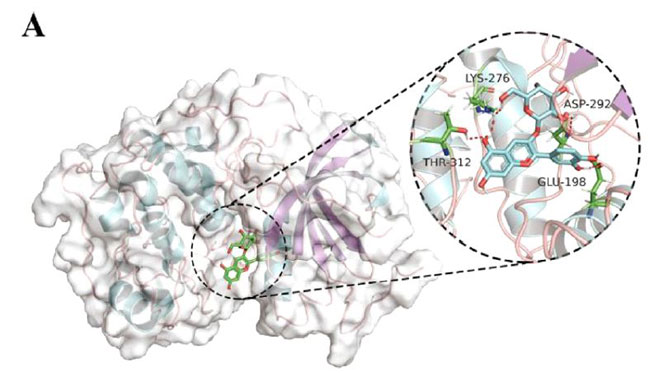
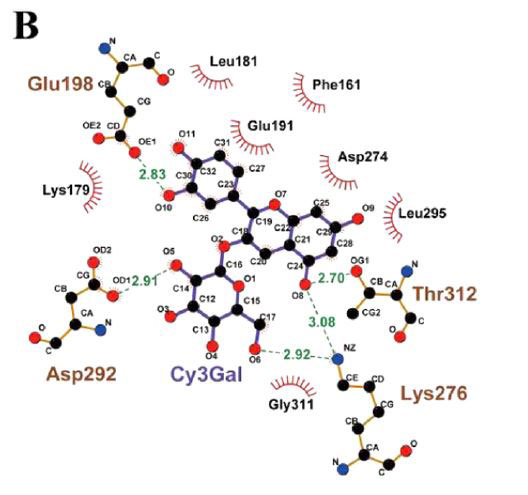
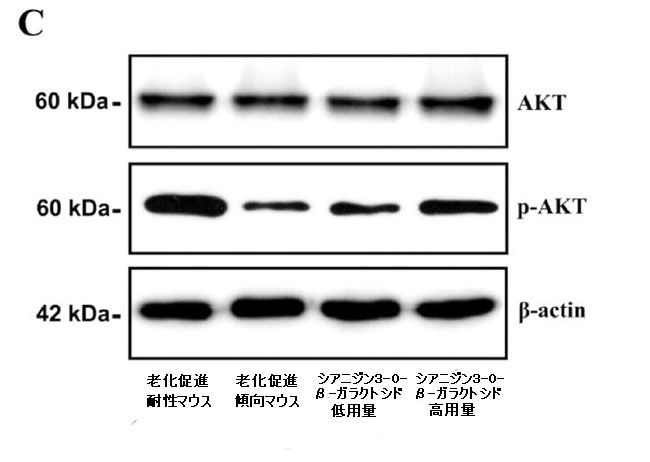
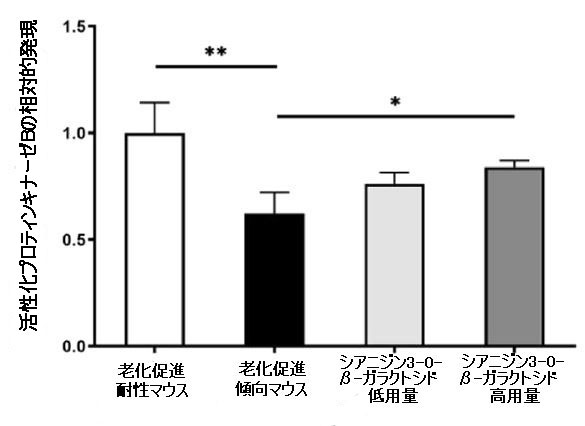
| 次に、脳内の認知障害とエネルギー代謝の改善に対するシアニジン3-O-β-ガラクトシドの影響の可能性を調査しました。 PI3K / Akt(訳者注:ホスファチジルイノシトール3-キナーゼ/プロテインキナーゼB)シグナル伝達は、エネルギー代謝とアポトーシスの調節に重要な役割を果たします。 同時に、PI3K / Akt軸の異常な制御は、アルツハイマー病の病原性ノードであると見なされ、異常なPI3K / Aktシグナル伝達は、アルツハイマー病の初期の特徴であると見なされます(34)。 したがって、シアニジン3-O-β-ガラクトシドがPI3K / Akt経路に影響を及ぼし、神経保護に有益な効果をもたらす可能性が最も高いです。 仮説を検証するために、最初に分子動力学シミュレーションを使用して、シアニジン3-O-β-ガラクトシドとPI3K / Aktの間の相互作用を予測しました。 分子ドッキングの結果は、シアニジン3-O-β-ガラクトシドとホスファチジルイノシトール3-キナーゼの結合が弱いことを示しました(補足図11A、Bおよび補足表2)。 ただし、シアニジン3-O-β-ガラクトシドとプロテインキナーゼBの組み合わせははるかに強力で、水素結合と疎水性の力の両方によって安定化されました。 図6A、Bに示すように、シアニジン3-O-β-ガラクトシドはプロテインキナーゼBの疎水性空洞に埋め込まれ、5つの水素結合(Glu198、Asp292、Thr312、およびLys276)と2つの疎水性相互作用(Asp274およびPhe161)に囲まれています。 特に、Asp274とPhe161はまさにプロテインキナーゼBの代謝部位です。 分子ドッキングの結果と一致して、シアニジン3-O-β-ガラクトシドはマウスの脳でプロテインキナーゼBの活性化を促進しましたが、ホスファチジルイノシトール3-キナーゼは促進しませんでした(図6Cおよび補足図11C)。 結果は、シアニジン3-O-β-ガラクトシドがプロテインキナーゼBの活性部位に結合し、下流のシグナル経路を活性化したことを示唆しました。 |
 |
 |
 |
 |
| 図6. シアニジン3-O-β-ガラクトシドとプロテインキナーゼBの間の相互作用 |
| プロテインキナーゼBとシアニジン3-O-β-ガラクトシド間の相互作用の分子ドッキング: (A)複雑なビュー (B)プロテインキナーゼBとシアニジン3-O-β-ガラクトシド間の2D相互作用の結合シミュレーション分析。 (C)代表的なウエスタンブロット画像およびマウス脳における活性化プロテインキナーゼBの分析(n = 4)。* p <0.05および** p <0.01。 |
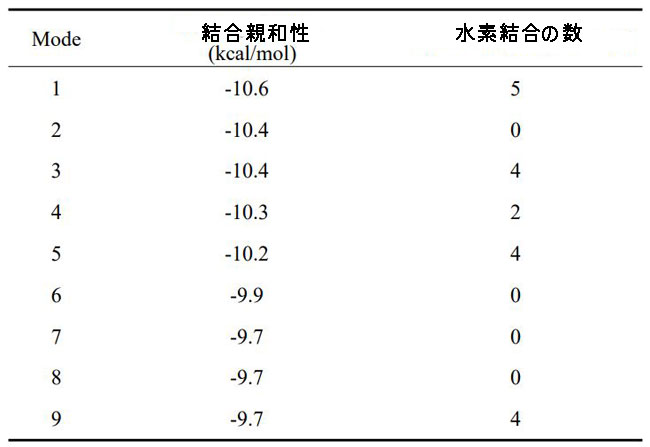 |
| 補足表1 シアニジン3-O-β-ガラクトシドとプロテインキナーゼBタンパク質の結合エネルギーと水素結合 |
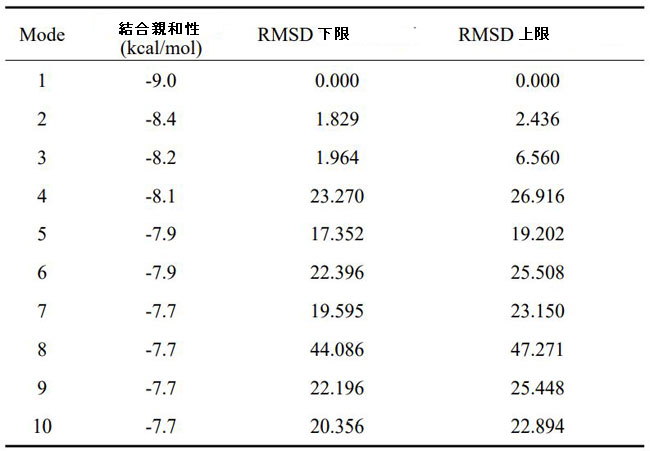 |
| 補足表2 シアニジン3-O-β-ガラクトシドおよびホスファチジルイノシトール3-キナーゼタンパク質の結合親和性、二乗平均平方根偏差(RMSD)の下限および上限 |
討論 |
アルツハイマー病の早期診断は、早期かつ効果的な治療的介入に不可欠であり、アルツハイマー病のリスクを軽減します。 PETは脳のブドウ糖利用を分析するために使用することができ、アルツハイマー病の早期診断のための最も信頼できるツールであると考えられています.(35)。 この研究では、PET / CTを使用して、加齢中のマウスの脳のエネルギー代謝の変化を監視し、アルツハイマー病関連の病状と症状に対する早期介入の効果をさらに評価しました。 脳のグルコース代謝は、神経変性疾患を含む多くの脳疾患の診断のための重要なツールになっています。 アルツハイマー病の患者の場合、脳(特に海馬と大脳皮質)でのブドウ糖利用の低下は、学習、記憶、および認知障害に関連しています(36)。 さらに、脳のグルコース取り込みの減少は、アルツハイマー病の神経萎縮および機能障害の早期かつ正確なバイオマーカーと見なされてきました(37)。 ますます多くの証拠が、脳のエネルギー代謝の障害が、病理学的変化および最終的な認知障害の前のアルツハイマー病の前臨床段階で発生することを示しています(38,39)。 脳内のエネルギー代謝の障害は、酸化ストレスを誘発し、神経伝達物質の生成をブロックすることにより、アルツハイマー病の病理学的プロセスをさらに促進します(40)。 一方、脳内のβ-アミロイド凝集体などの有毒タンパク質は、ミトコンドリアの機能障害や酸化ストレスを誘発し、エネルギー代謝の障害を加速させ、最終的に悪循環を形成する可能性があります(41)。 したがって、ここでは、エネルギー代謝の観点からアルツハイマー病の治療のための潜在的な戦略を提供し、有意な効果が観察されました。 |
| アルツハイマー病患者のほぼ95%は、家族歴のない散発性アルツハイマー病(sAD)でした。 散発性アルツハイマー病は加齢性認知症の最も一般的な形態です(42)。 したがって、適切な動物モデルは、散発性アルツハイマー病の病因と効果的な薬剤の開発をさらに解明するためにも重要です。 シアニジン3-O-β-ガラクトシド投与の認知効果を調査するためにアルツハイマー病マウスモデルとして老化促進傾向マウス(SAMP8)を使用しました。 老化促進傾向マウスは、遅発性アルツハイマー病の散発的な症例の研究に大きな利点があります。 これは、加齢に伴う認知障害と、老化防止因子の異常な発現、酸化ストレス、炎症、β-アミロイド沈着、タウの過剰リン酸化、小胞体ストレス、異常なオートファジー活動、腸内細菌叢の破壊など、アルツハイマー病の多くの病理学的特徴を示します(43)。 さらに、正常な老化表現型を持つ老化促進耐性マウス(SAMR1)は、老化促進傾向マウスのバックグラウンドコントロールとして一般的に使用されます。 したがって、老化促進傾向マウスは、散発性アルツハイマー病の薬剤を研究する上で適切な動物モデルでもありました。 |
| 栄養の観点から、アントシアニンはアルツハイマー病の大きな予防的および治療的可能性を示しています(44)。 さらに、天然化合物として、アントシアニンはより安全で入手が容易です。 マウスの老化の進行に介入するために体内でより速く吸収されるベリーアントシアニン、シアニジン3-O-β-ガラクトシドを選択します。 マウスの脳の糖代謝は加齢とともに低下するが、脳のさまざまな領域の糖代謝はシアニジン3-O-β-ガラクトシド投与後に有意な低下を示さなかったことがわかった。 脳のさまざまな領域の中で、嗅球が懸念の原因でした。 β-アミロイドが蓄積しやすい嗅球の糖代謝は、加齢とともに大幅に減少しました。 同様に、シアニジン3-O-β-ガラクトシドは、この領域でのグルコース代謝とβ-アミロイドの蓄積にあまり影響を与えませんでした(補足図2および7)。 以前の研究では、アルツハイマー病患者はβ-アミロイド沈着と認知障害の前に嗅覚機能障害を発症したことが示されています(45)。 今回の結果は、嗅球領域が加齢に敏感であり、この領域をグルコース代謝率を検出するための標的領域として使用できることを示唆しており、これを使用してアルツハイマー病を早期に予測または診断することができます。 |
| ミトコンドリアは真核生物のエネルギー代謝の部位であり、正常な機能を維持している場合にのみ、神経細胞の機能をサポートするのに十分な燃料を供給することができます。 ミトコンドリアのエネルギー代謝のレベルが低下すると、インスリンシグナル伝達、グルコース受容体の変化、および星状細胞の代謝表現型に影響を及ぼし、アルツハイマー病の発症を加速します(46)。 さらに、ミトコンドリアによって調節される神経細胞のアポトーシスも、アルツハイマー病の病因の重要な理由です(47)。 アポトーシスはアルツハイマー病の初期段階で発生します。 脳内の神経細胞の喪失は、アルツハイマー病の最も重要な病理学的特徴の1つです。 シアニジン3-O-β-ガラクトシドは、加齢に伴うマウスの脳のアポトーシス、特に海馬と大脳皮質の神経細胞の損傷を弱めることがわかりました(図2および補足図5)。 モリス水迷路テストで観察されたように、マウス海馬のCA1およびDG領域の神経細胞のアポトーシスは、神経細胞の障害、シグナル伝達の妨害、およびマウスの認知機能の低下につながる可能性があります(図4)。 特に、ミトコンドリアの品質管理の重要な要素であるBcl-2ファミリータンパク質は、ミトコンドリアのオートファジー、アポトーシス、有糸分裂、および融合の調節に関与しています(48,49)。 シアニジン3-O-β-ガラクトシドは、老齢マウスの脳組織におけるBcl-2発現を増加させ、カスパーゼアポトーシス経路の活性化を阻害し、大脳皮質と海馬の神経細胞を効果的に保護しました(図2と3および補足図5)。 さらに、老化促進傾向マウスではシアニジン3-O-β-ガラクトシドによって営巣行動が改善され(補足図8)、これは海馬機能の改善と密接に関連している可能性があります(26)。 分子標的を見つけ、エネルギー代謝とミトコンドリア機能を調節するシアニジン3-O-β-ガラクトシドの経路を説明するために、我々は主要な上流モジュレーターを専攻しました。 PI3K / Akt軸は、ほとんどすべての主要な老化疾患の病原性ノードであるため、広く懸念されています。 PI3K / Akt経路は、代謝制御、ストレス反応、シナプス可塑性、タンパク質の安定性など、アルツハイマー病の重要なイベントで重要な役割を果たします(34)。 さらに、PI3K / Akt経路は、細胞内のグルコース取り込みと代謝に関与する細胞エネルギー代謝の重要な調節因子です。 プロテインキナーゼB(Akt)は解糖系酵素の発現を誘導し、細胞膜上のグルコース受容体の発現と動員を促進することができます。 この研究では、プロテインキナーゼBはシアニジン3-O-β-ガラクトシドによって活性化されました(図6A)。 分子ドッキングは、シアニジン3-O-β-ガラクトシドがプロテインキナーゼBの疎水性空洞に水素結合と疎水性相互作用で結合していることを示しました(図6B、C)。 一方、シアニジン3-O-β-ガラクトシドには「ニューラルアベイラビリティ」があり、プロテインキナーゼBのターゲットに昇格し、ダウンストリームのシグナルカスケードを引き起こしました。 しかし、プロテインキナーゼBの下流因子がエネルギー代謝とニューロンアポトーシスを間接的に調節する方法にすぎないと結論付けるのは難しいです。 これは、哺乳類のラパマイシン標的であるグリコーゲンシンターゼキナーゼ-3β(GSK-3β)のように、経路で重要な役割を果たす他のシグナルタンパク質もいくつかあるためです(51,52)。 それらは常にアルツハイマー病薬の開発と研究のターゲットとして使用されます(53)。 |
| 私たちの研究の結果に基づいて、加齢によって引き起こされる認知障害の緩和に対するシアニジン3-O-β-ガラクトシドの効果を図7で結論付けることができます。 |
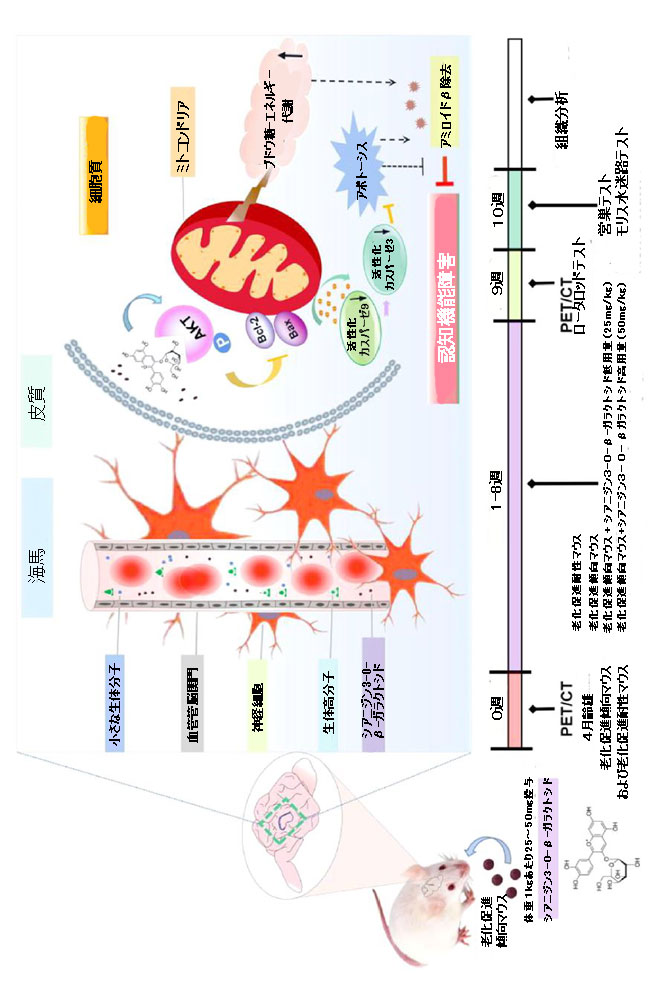 |
| 図7 アルツハイマー病モデル老化促進傾向マウス(上のパネル)および動物実験プロトコル(下のパネル)における認知障害を改善するシアニジン3-O-β-ガラクトシドのメカニズムを表す概略図 |
| シアニジン3-O-β-ガラクトシドによる8週間の介入は、加齢中のマウス脳のグルコース取り込みを改善し、神経損傷、β-アミロイド発現、ミトコンドリア機能障害などの病理学的変化を改善し、加齢マウスの空間学習と記憶障害に大きな影響を与えることを実証しました 。 |
| 結果は、加齢に伴うマウスの脳におけるエネルギー代謝の課題に対する認知と神経保護の改善におけるシアニジン3-O-β-ガラクトシドの有効性を確認しました。 プロテインキナーゼBは、シアニジン3-O-β-ガラクトシドがアルツハイマー病に対して早期の利益を発揮するためのターゲットである可能性があります。 私たちの研究は、健康な脳の老化のインターフェースで有望な栄養戦略を明らかにしています。 |
|
参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
この文献は、J. Agric. Food Chem. 2022, 70, 4, 1111–1121に掲載されたCyanidin 3‑O‑β-Galactoside Alleviated Cognitive Impairment in Mice by Regulating Brain Energy Metabolism During Aging.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |