Chia-Chia Lee et al., |
要約 |
| ヒトの老化は、生物学的、生理学的、および心理学的機能の進行性の低下を特徴とし、慢性疾患の発症における主要なリスク要因です。したがって、老化関連障害を軽減し、健康的な老化を促進することを目的とした戦略の開発が重要です。以前の研究では、台湾の漬物キャベツから分離されたプロバイオティクス株であるLactobacillus plantarum TWK10(L.plantarumTWK10)が、健康なヒトの筋力、運動持久力、および全体的な体組成を改善することを実証しました。この研究では、マウスでL.plantarumTWK10の加齢関連障害の進行に対する効果を調査しました。L.plantarumTWK10は若いマウスの筋力を強化するだけでなく、筋肉グリコーゲンレベルの上昇を伴う高齢マウスの加齢関連の筋力低下を防ぐこともわかりました。さらに、L.plantarumTWK10は、学習能力と記憶能力、および骨量の加齢関連低下を軽減しました。 16S rRNA 遺伝子の次世代シーケンシング (NGS) を使用した腸内細菌叢のさらなる分析により、L.plantarumTWK10を 8 週間投与した後、腸内微生物組成のパターンが明らかに変化したことが示されました。L.plantarumTWK10を投与されたマウスでは、短鎖脂肪酸 (SCFA) 産生細菌が増加し、腸内 短鎖脂肪酸の全体的なレベルも高くなりました。さらに、L.plantarumTWK10投与により、老化に伴う病原性細菌の蓄積がある程度逆転しました。結論として、L.plantarumTWK10は、腸内細菌叢の不均衡を調整することで、老化関連疾患を軽減し、健康上の利点をもたらす潜在的な治療薬と見なすことができます。 |
| 目次(クリックして記事にアクセスできます) |
| 1.はじめに |
| 2.材料と方法 |
| 2.1.動物、プロバイオティクス、研究デザイン |
| 2.2.前肢の握力 |
| 2.3.モリス水迷路 |
| 2.4.体組成 |
| 2.5.骨質と血清ビタミン D の測定 |
| 2.6.筋肉グリコーゲン濃度 |
| 2.7.組織学的分析 |
| 2.8.DNA 抽出および 16S rRNA 遺伝子配列分析 |
| 2.9.短鎖脂肪酸(SCFA)レベルの測定 |
| 2.10.統計分析 |
| 3.結果 |
| 3.1.L.plantarumTWK10 が加齢に伴う骨格筋力の低下に及ぼす影響 |
| 3.2.L.plantarumTWK10の加齢に伴う骨量減少に対する効果 |
| 3.3.L.plantarumTWK10が加齢に伴う空間学習および記憶障害に与える影響 |
| 3.4.L.plantarumTWK10の体脂肪調節効果 |
| 3.5.L.plantarumTWK10の腸内細菌叢への影響 |
| 3.6.L.plantarumTWK10の腸内細菌ネットワークへの影響 |
| 3.7.L.plantarumTWK10が 短鎖脂肪酸産生腸内細菌の豊富さと腸内短鎖脂肪酸レベルに与える影響 |
| 3.8.腸内細菌叢の構成とL.plantarumTWK10による健康効果の相関関係 |
| 4.考察 |
| 4.1.L.plantarumTWK10の筋力に対する効果 |
| 4.2.L.plantarumTWK10の骨質への影響 |
| 4.3.L.plantarumTWK10の体脂肪に対する効果 |
| 4.4.L.plantarumTWK10の学習と記憶に対する効果 |
| 4.5.L.plantarumTWK10の腸内細菌叢に対する効果 |
| 5.結論 |
| 本文 |
| 1.はじめに |
| 老化は、身体能力、身体組成、学習と記憶、社会的および心理的反応、関節、代謝調節の悪影響を伴う進行性のプロセスです。組織と臓器の機能の老化に伴う低下は、慢性疾患の発症における主要なリスク要因です (1)。老化は、除脂肪体重と骨密度の減少、および脂肪量の増加を伴います。サルコペニアは、一般的に、加齢に伴って骨格筋量と筋力が徐々に低下することと定義されます (2、3)。筋肉量は 30 歳を過ぎると 10 年ごとに約 3~8% 減少し、60 歳を過ぎると減少率が加速します (4)。筋肉量の減少は、転倒や骨折、身体障害、生活の質の低下、および死亡のリスク増加と密接に関連しています。さらに、脂肪量は通常、加齢とともに徐々に増加し、特に腹部に局在します。腹部の脂肪量の蓄積は、心血管疾患や糖尿病などの代謝障害の発症と密接に関連しています (5)。 |
| 腸内細菌叢は健康維持に重要な役割を果たしており、腸内細菌叢の不均衡は老化と密接に関係しています (9、10)。高齢者の腸内細菌叢は若年成人とは異なることがわかっています。加齢とともに、腸内細菌叢の多様性は低下する一方で、Enterobacteria やClostridium属などの日和細菌のレベルは高くなるようです (10、11)。短鎖脂肪酸は、食物繊維発酵の最終産物として嫌気性腸内細菌によって生成されます。結腸に存在する総短鎖脂肪酸の 90% は、酪酸、プロピオン酸、酢酸で占められています。これらの代謝物は、代謝、腸管透過性、炎症反応、免疫機能を調節することで、宿主生理学において重要な役割を果たすことが示されています (12–15)。 短鎖脂肪酸は、大腸がん、炎症性腸疾患、糖尿病など、いくつかの疾患から宿主を保護する役割も果たしています (16–18)。Bacteroidetes 門とFirmicutes 門は、ヒトの腸内で最も多く存在する門で、細菌組成の 85% 以上を占めています (19)。ヒトの腸内では、Bacteroidetes門のメンバーは主に酢酸とプロピオン酸を産生し、Firmicutes 門の集団は主に酪酸を産生します (20)。老化マウスモデルにLactobacillus acidophilus DDS-1 を投与すると、腸内細菌組成の変化が引き起こされ、代謝表現型の改善につながり、盲腸と粘膜の短鎖脂肪酸産生が促進されることが示されています (21、22)。したがって、短鎖脂肪酸産生の刺激と腸内の短鎖脂肪酸産生細菌の濃縮は、全体的な健康の改善に不可欠です。 |
| 人口の高齢化は、すべての人類が直面する深刻な問題になりつつあります。2050年までに、65歳以上の人口は世界で15億人に達すると予想されています(World Population Aging 2019 Highlights、https://digitallibrary.un.org/record/3846855)。高齢者人口が急速に増加する中、老化関連疾患を軽減し、健康的な老化を促進するための戦略の開発が重要な問題となっています。最近の報告では、老化と戦うための潜在的な治療薬としてプロバイオティクスサプリメントの使用が強調されています。たとえば、Lactobacillus salivarius FDB89は、Caenorhabditis elegans(23)の成長を促進し、寿命を延ばすことが示されています。Leeら(24)は、Lactobacillus plantarum HY7714のヒトでの皮膚の抗老化特性も実証しました。Lactococcus lactis subsp. lactis Plasma菌株の摂取は、筋肉の変性に関連する遺伝子の発現を減少させ、老化を遅らせました (25)。さらに、Lactobacillus paracasei PS23 の摂取は、加齢に伴う認知機能の低下と筋肉の減少を緩和しました (26、27)。他の研究群は、Lactobacillus属の補給が、卵巣摘出 (OVX) 誘発性骨粗鬆症マウスモデルにおける骨量減少を予防することを実証しました (28、29)。しかし、これらのプロバイオティクスが腸内細菌叢に与える影響だけでなく、その根底にあるメカニズムは不明のままです。 |
| これまでの研究で、Lactobacillus plantarum TWK10(L.plantarumTWK10)はマウスとヒトの体組成、特に筋肉量、筋肉の質、脂肪量に有益な効果を発揮することを実証しました(30、31)。本研究では、(i)老化の進行、すなわち筋肉の減少、筋力の低下、認知障害、体脂肪の蓄積、骨密度の低下を緩和するL.plantarumTWK10の潜在的な役割を分析し、(ii)L.plantarumTWK10が腸内細菌叢に与える影響を調査することを目的としました。 |
| 2.材料と方法 |
| 2.1.動物、プロバイオティクス、研究デザイン |
| 雄の ICR マウスは BioLASCO (Charles River Licensee Corp.、Yi-Lan, Taiwan) から購入しました。すべてのマウスは湿度管理 (65 ± 5%)、温度管理 (24 ± 2°C) された環境で飼育され、12:12 の明暗サイクルで飼育され、標準的な実験用飼料 (No. 5001、PMI Nutrition International、Brentwood, MO, USA) と蒸留水が自由に摂取できました。すべての実験手順は、台湾桃園市にある国立台湾体育大学の動物実験委員会 (IACUC) によって承認されました (IACUC-10712)。 L.plantarumTWK10は、台湾の漬物キャベツ(33)から分離された植物由来のL. plantarum株(最近Lactiplantibacillus plantarum(32)として再分類された)であり、SYNBIO TECH INC.カルチャーコレクション(Kaohsiung, Taiwan)から入手されました。マウスに投与するために、L.plantarumTWK10の濃度を200μlリン酸緩衝生理食塩水(PBS)で1×109 CFUに調整しました。すべてのマウスは、まず年齢によって若齢群(4ヶ月齢、n = 17)と高齢群(19〜22ヶ月齢、n = 16)に分けられました。次に、これら 2 群のマウスをそれぞれ 2 つの実験群にランダムに割り当てました。Y- Control (リン酸緩衝生理食塩水、200 μl/マウス/日を投与された若いマウス) と Y-TWK10 (L.plantarumTWK10、1 × 109 CFU/マウス/日を投与された若いマウス)、および A-Control (リン酸緩衝生理食塩水、200 μl/マウス/日を投与された老齢マウス) と A-TWK10 (L.plantarumTWK10、1 × 109 CFU/マウス/日を投与された老齢マウス)。実験中、体重、食物摂取量、水分摂取量は毎週記録されました。また、腸内細菌叢の分析のために、ベースラインと治療開始後 8 週間で排泄されたばかりの糞便サンプルも収集されました。前肢の握力は、ベースライン (L.plantarumTWK10投与前)、4 週間、および 8 週間 (L.plantarumTWK10 投与後) にモニタリングされました。L.plantarumTWK10 投与から 8 週間後、すべてのマウスの体組成と認知機能を評価しました。その後、マウスは殺され、組織の重量、骨の質、筋肉のグリコーゲンに関する測定値を取得し、組織学的分析を実施しました。 |
| 2.2.前肢の握力 |
| 低力試験システム(Model-RX-5、アイコーエンジニアリング、Nagoya, Japan)を使用して、前述のように(34)、治療を受けたマウスの前肢の絶対握力を測定した。簡単に説明すると、マウスの尾の付け根をつかみ、バーに向かって垂直に降ろした。マウスは、2本の足(前肢)でバーをつかんだ状態で尾でわずかに後方に引っ張られ、「逆引き」が引き起こされた。10回の試行での最大握力が、絶対前肢握力として記録された。前肢の握力は、L.plantarumTWK10/リン酸緩衝生理食塩水投与後、ベースライン、4週間、8週間でモニタリングされた。 |
| 2.3.モリス水迷路 |
| モリス水迷路 (MWM) は、以前説明したように (35)、マウスの空間学習と記憶を評価するために使用されました。簡単に言うと、モリス水迷路テストは、毒性のないテンペラ ペイントで不透明に着色された水 (26 ± 1°C) が入った円形のプール (直径 100 cm、深さ 30 cm) で実施されました。プラットフォームとマザール外の手がかりは、学習試験中 (1 日目と 2 日目) を通じて同じ位置に留まり、マウスは 1 日に 3 回、水面下のプラットフォームを見つけるように訓練されました。水迷路には 3 つの開始位置があり、マウスは毎日異なる開始位置から運動を開始するようにしました。最大水泳時間は 90 秒に設定されました。3 日目のマウスの逃走潜時を使用して、空間学習と記憶を調べました。 |
| 2.4.体組成 |
| 生きたマウスの脂肪組織と除脂肪組織の測定には、マイクロ CT スキャン (Bioscan、Washington, DC, USA) を使用しました。ハウンスフィールド単位 (HU) の較正のために、マイクロ CT 空気/水ファントム スキャン (CT QC Phantom、Mediso、Hungary) を実施しました。すべてのマウスは、足を伸ばしたうつ伏せの姿勢で 1% イソフルラン吸入により麻酔されました。CT イメージングは、65 kVp、1,000 ms、0.123 mAs の露出、1:1 ビニング係数で、1 回転あたり 180 投影を使用して実施しました。下半身 (L1 レベルの大腿骨と腹部の間) の連続的な横断画像が得られました。CT 投影は、ボクセル サイズ 0.1475 × 0.1475 × 0.1477 mm3 で再構成されました。画像解析は、PMOD(バージョン 3.7 PMOD Technologies LTD、Zurich, Switzerland)解析ソフトウェアを使用して実施しました。画像は、まずガウス 3D フィルターを使用して平滑化され、次に組織密度に従ってセグメント化され、最初に総体積、次に脂肪体積がセグメント化されました (36)。L1 から L5 までの腹部の関心体積のヒストグラムが生成され、分布は二峰性で、1 つのモードは脂肪組織ボクセルを表し、他のモードは除脂肪組織ボクセルを表します (37)。本研究では、すべてのマウスについて、既知の脂肪組織領域と除脂肪組織領域に最適な分離方法に従って、総体積には -300 ~ +3,500 HU、脂肪体積には -200 ~ -50 HU の固定しきい値を使用しました。 |
| 2.5.骨質と血清ビタミン D の測定 |
| マウスから大腿骨を採取し、分析までメタノールで保存しました。各標本のマイクロ CT 画像は、SkyScan 1076 マイクロ CT (Bruker、Kontich, Belgium) を使用して取得しました。スキャン パラメータは、50 kV、200 μA、回転ステップ 0.5°、露出時間 2.1 秒、0.5 mm Al フィルター、および 9 μm/ピクセルのスキャン解像度に設定されました。骨質パラメータは、CT-Analyser ソフトウェア (Bruker) を使用して計算されました。以下の3次元(3-D)パラメータを評価しました:組織容積(TV)、骨容積(BV)、骨容積率(BV/TV)、骨表面(BS)、骨表面密度(BS/BV)、骨梁厚(Tb.Th)、骨梁間隔(Tb.Sp)、骨梁数(Tb.N)、骨梁パターン係数(Tb.Pf)、構造モデル指数(SMI)、および骨密度(BMD)。血清ビタミンD(1,25-(OH)2ビタミンD)は自動分析装置(Hitachi 7060, Hitachi, Tokyo, Japan)を使用して測定しました。 |
| 2.6.筋肉グリコーゲン濃度 |
| マウスの腓腹筋を分離し、その後の分析まで -80°C で保存しました。グリコーゲン含有量は、前述のように測定しました (38)。簡単に説明すると、100 mg の筋肉組織を 0.5 ml の冷過塩素酸でホモジェナイズしました。ホモジェネートを 15,000 × g で 15 分間 4°C で遠心分離し、上清を採取してからグリコーゲン濃度を測定しました。グリコーゲン濃度 (mg/g 筋肉) は、市販のアッセイキット (Sigma-Aldrich、St. Louis, MO, USA) を使用し、製造元の指示に従って測定しました。 |
| 2.7.組織学的分析 |
| 8 週間の治療後、精巣上体脂肪体 (EFP)、肩甲骨間褐色脂肪組織 (BAT)、腓腹筋を慎重に除去し、10% ホルマリンで一晩固定した後、パラフィンに包埋しました。パラフィン包埋組織を 4 μm で切片化し、ヘマトキシリンおよびエオシン (H&E) で染色して、光学顕微鏡 (BX-51、Olympus, Tokyo, Japan) を使用して精巣上体脂肪体 (EFP)と肩甲骨間褐色脂肪組織 (BAT)の組織学的検査を行いました。脂肪細胞の断面積 (CSA、μm2)、高倍率視野 (HPF) あたりに観察された肩甲骨間褐色脂肪組織 (BAT)脂肪細胞の平均数、および H&E 染色後の肩甲骨間褐色脂肪組織 (BAT)の平均相対褐色面積 (%) を、ImageJ ソフトウェア (NIH, MD, USA) を使用して測定し、分析しました。 |
| 2.8.DNA 抽出および 16S rRNA 遺伝子配列分析 |
| 8 週間の治療後、マウスから排泄されたばかりの糞便サンプルを採取しました。細菌ゲノム DNA を抽出するために、糞便サンプルを 1.0 ml のリン酸緩衝生理食塩水で 3 回洗浄し、14,000 × g で 5 分間遠心分離しました。糞便ペレットをリゾチーム (最終濃度 10 mg/ml) を含む 180 μl TE緩衝液に再懸濁し、ガラスビーズ (300 mg、直径 0.1 mm、Biospec、Bartesville, OK, USA) を含む懸濁液を FastPrep 24 ホモジナイザー (MP Biomedicals、USA) を使用して 30 秒間ホモジナイズし、細胞壁が完全に破壊され、DNA 分子が溶液に放出されるようにしました。次に、細菌ゲノム DNA を、製造元の指示に従って、ゲノム DNA ミニキット (Geneaid、Taipei, Taiwan) を使用して抽出しました。 DNA濃度は、BioDrop装置(Biochrom、Biochrom Ltd.、Cambridge, UK)を用いた分光光度計で測定した。DNAサンプルは、さらなる処理まで-20°Cで保存した。16S rRNA遺伝子のV3-V4領域は、16Sメタゲノムシーケンシングライブラリ調製手順(Illumina)に従って、特異的プライマー(319F:5′-CCTACGGGNGGCWGCAG-3′および806R:5′-GACTACHVGGGTATCTAATCC-3′)(39)を使用して増幅した。 |
| アンプリコンプールは、Illumina MiSeq™ シーケンシングプラットフォーム (Illumina、San Diego, CA, USA) でシーケンスされました。生のシーケンスデータは、q2-demux プラグインを使用してデマルチプレックス化され、QIIME 2 2019.7 (41) を使用して DADA2 (40) (q2-dada2 経由) で最小限の品質フィルタリングが行われ、結果としてアンプリコンシーケンスバリアント (ASV) が得られました。分類は、q2-feature-classifier (42)、SILVA データベース (リリース 132) に対する classify-sklearn ナイーブベイズ分類分類器、および 99% の類似性を持つ代表シーケンス (43) を使用して ASV に割り当てられました。アルファ多様性 (シャノン指数とリッチネス指数) とベータ多様性の両方が、30,000 シーケンスの希薄化を使用して QIIME2 で推定されました。ベータ多様性分析は、R ソフトウェアの「vegan」パッケージ (Vegan: Community Ecology Package。R パッケージ バージョン 2.5-6) の metaMDS 関数による重み付き UniFrac 距離または重みなし UniFrac 距離に基づく非計量多次元尺度法 (NMDS) 順序プロットを使用して実行されました。R ソフトウェアの vegan パッケージを使用して、999 の順列に基づいて有意差を調べるために、順列多変量分散分析 (PERMANOVA)/Adonis テストが適用されました。この記事の調査結果を裏付ける生のシーケンス ファイルは、プロジェクト アクセス番号 PRJNA726848 で NCBI Sequence Read Archive (SRA) データベースに保存されています。 |
| 2.9.短鎖脂肪酸レベルの測定 |
| マウスの盲腸を採取し、直ちに液体窒素で凍結し、次の処理まで-80°Cで保存した。盲腸内容物1mgを70%エタノール溶液10μlと混合し、適量のガラスビーズ(直径1.0mm、Biospec Products)とともに3,000rpmで10分間ボルテックスしてホモジナイズした。ホモジナイズしたサンプルを14,000×gで10分間遠心分離し、上清を収集して、前述の方法(44)に従って脂肪酸誘導体化処理した。誘導体化した上清は、0.22μmポリカーボネートシリンジフィルター(Millipore、St. Charles, MO, USA)を使用して濾過した。短鎖脂肪酸の分析は、高速液体クロマトグラフィー(HPLC)(HITACHI、Tokyo, Japan)を使用して実施した。 短鎖脂肪酸は、C18 HTec カラム (NUCLEODUR、Macherey-Nagel、Düren, Germany) を使用して、カラム温度 40°C、流速 1 ml/分、検出波長 400 nm に設定して分離されました。 |
| 2.10.統計分析 |
| データは平均±SDとして表されます。統計分析はGraphPad Prism 7.04(GraphPad Software、San Diego, CA)を使用して実施しました。複数の群の比較については、データは事後Tukey-Kramer検定を伴う2元配置分散分析によって分析されました。事後Dunn検定を伴うKruskal-Wallis検定は、複数のノンパラメトリック比較に使用されました。2群の比較については、パラメトリックまたはノンパラメトリックデータ分析にそれぞれStudentの無対t検定とMann-Whitney U検定を使用しました。前肢の握力と筋グリコーゲンレベル、および腸内細菌叢の豊富さと加齢に関連する表現型特性との相関分析には、それぞれピアソンの相関係数とスピアマンの相関係数を使用しました。P < 0.05の場合、差異は統計的に有意であるとみなされました。 |
| 3.結果 |
| 3.1.L.plantarumTWK10 が加齢に伴う骨格筋力の低下に及ぼす影響 |
| 4 つの実験群(Y-Control、Y-TWK10、A-Control、A- TWK10 のすべてのマウスに 8 週間にわたって同一の食事を与え、その平均体重、食物摂取量、水分摂取量を表 1 にまとめました。4 つの群間で体重、食物摂取量、水分摂取量に有意差は認められませんでした。 |
| 表1.動物の一般的な特徴 |
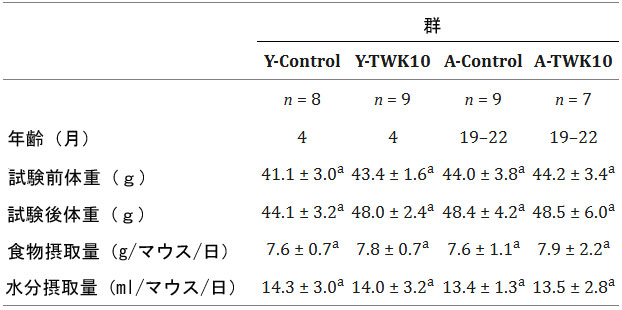 |
| データは平均±SDとして表されます。群間の統計的差異はTukey-Kramer検定によって分析されました。行内で、同じ上付き文字(a)は有意差がないことを示します(P> 0.05)。 |
| 若年マウス群と高齢マウス群の筋力のベースラインレベルを評価するために、L.plantarumTWK10投与前に全マウスの前肢握力測定を行った。高齢マウス群(A-ControlおよびA-TWK10、106.1 ± 3.7 g、n = 16)の前肢握力は、若年マウス群(Y-ControlおよびY-TWK10、144.7 ± 20.7 g、n = 17)よりも有意に(P < 0.001)低かった(図1A)。0週目では、Y-Control群とY-TWK10群、およびA-Control群とA-TWK10群の間に握力に有意差は認められなかったが、若年マウス群の握力レベルは高齢マウス群よりも有意に高かった(P < 0.001)。二元配置分散分析の結果、因子「加齢(若年 vs. 高齢)」および「治療(対照 vs. TWK10投与)」が8週目の握力に有意(P < 0.05)に影響することが示された。TWK10投与4週後および8週後、高齢マウス群の握力はそれぞれ若年マウス群と比較して有意に低かった[Y-TWK10では4週目と8週目(P < 0.001)、A-TWK10では4週目(P < 0.001)と8週目(P < 0.01)]。一方、TWK10投与群の握力は、それぞれY対照群およびA対照群と比較して有意に高かった[Y-TWK10では4週目と8週目(P < 0.05)、A-TWK10では4週目(P < 0.05)と8週目(P < 0.01)]。対照老齢マウス(A-Control)では、0週目と比較して4週目と8週目の両方で握力の有意な低下(P < 0.001)が観察されました(図1B)。さらに、老齢マウス群の腓腹筋重量は、若年マウス群と比較して有意に低かった(P < 0.05)。しかし、L.plantarumTWK10投与後、若年マウスと老年マウスの両方で腓腹筋重量に有意差は観察されませんでした(図1C)。老年マウス(A-Control)の腓腹筋グリコーゲンレベルは、若年マウス(Y-Control)と比較して有意に低かった。しかし、L.plantarumTWK10投与により、若年マウス群のグリコーゲンレベルは有意に増加し(P < 0.05)、老年マウスではわずかに増加しました(図1D)。握力と筋グリコーゲンレベルの間には正の相関関係が、若いマウス(ピアソンの相関係数、r = 0.630、P = 0.007)と老齢マウス(ピアソンの相関係数、r = 0.578、P = 0.019)の両方で観察されました(図1E)。 |
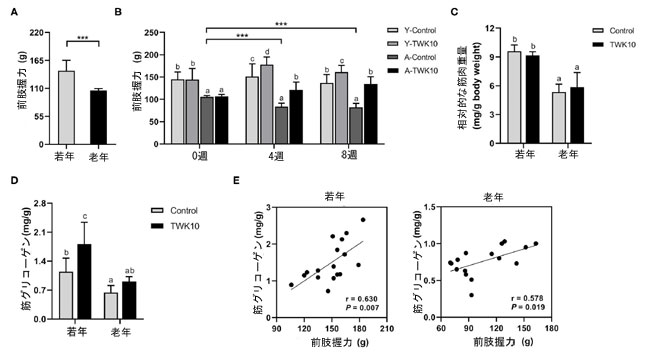 |
図1 Lactobacillus plaantarumTWK10 は、筋肉の質を改善することで、加齢に伴う骨格筋の強度低下を緩和しました。 (A) 若いマウス (n = 17) と高齢マウス (n = 16) の前肢握力を投与前に測定しました。 (B) マウスはL.plantarumTWK10 または リン酸緩衝生理食塩水で 8 週間処理され、前肢握力は 0 週目、4 週目、8 週目に測定されました。 (C、D) 腓腹筋を摘出し、筋肉の湿重量とグリコーゲン レベルを測定しました。 データは平均 ± SD として表されます。群間の統計的差異は、事後 Tukey-Kramer 検定による 2 元配置分散分析によって分析されました。異なる文字 (a、b、c) は群間の有意差を示します (P < 0.05)。2 群の比較は、対応のない Student t 検定を使用して分析されました。***P < 0.001。 (E) L.plantarumTWK10/PBS 投与 8 週間後の若年マウスと高齢マウスの握力と筋グリコーゲン レベル間のピアソン相関。 |
| 3.2.L.plantarumTWK10の加齢に伴う骨量減少に対する効果 |
| 骨質の回復における L.plantarumTWK10の有効性を評価するために、マウスの大腿骨を 8 週間の治療後に採取し、皮質および海綿骨の微細構造をマイクロ CT を使用して分析しました。大腿骨の 骨密度(BMD)および海綿骨パラメータに対するL.plantarumTWK10の影響を表 2 にまとめます。加齢に伴う骨量減少は、Y-Control群と A-Control群を比較した場合、骨梁数(Tb.N)の有意な減少 (P < 0.05) および 骨梁間隔(Tb.Sp) の増加 (P = 0.1547) として示されました。Y-TWK10群の骨容積率(BV/TV)値は Y-Controlマウスと比較して有意に高く (P < 0.05)、Y-TWK10 群の骨梁間隔(Tb.Sp)値は有意に低く (P < 0.05) なりました。 L.plantarumTWK10の投与により、Controlマウス (Y-Control群と A-Control群) と比較して、Y-TWK10 マウスと A-TWK10 マウスの 骨梁数(Tb.N) が有意に増加しました (P < 0.05)。A-TWK10 の血清ビタミン D レベルも A-Control群と比較して増加しました (P = 0.0584)。4 つの群 (Y-Control、Y-TWK10、A-Control、および A-TWK10) 間で、骨梁厚(Tb.Th)、骨梁パターン係数(Tb.Pf)、構造モデル指数(SMI)、または 骨密度(BMD)値に有意差は認められませんでした。 |
| 表2. Lactobacillus plantarum TWK10 によって骨質が改善されました。 |
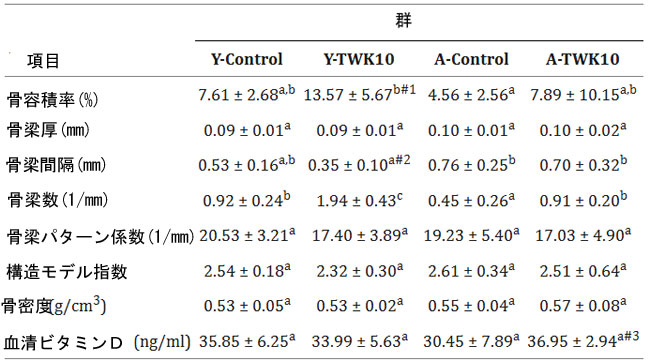 |
| データは平均±SDとして表される。群間の統計的差異は、事後Tukey-Kramer検定を用いた二元配置分散分析によって分析された。ノンパラメトリックデータは、Dunn検定を用いたKruskal-Wallis検定によって統計的に分析された。行内の異なる文字(a、b、c)は、群間の有意差を示す(P < 0.05)。骨容積率(BV/TV)(P = 0.0274)#1に関するControl群とTWK10投与若年群間の統計的差異は、Mann-Whiney U検定によって分析された。骨梁間隔(Tb.Sp)(P = 0.0173)#2に関するControl群とTWK10投与若年マウス群間の統計的差異、および血清ビタミンD(P = 0.0584)#3に関するControl群とTWK10投与老齢マウス群間の統計的差異は、それぞれStudentの無対t検定によって分析された。BV/TV:骨容積率。 Tb.N:骨梁数、Tb.Sp:骨梁間隔、Tb.Pf:骨梁パターン係数、BMD:骨密度、SMI:構造モデル指数。 |
| 3.3.L.plantarumTWK10が加齢に伴う空間学習および記憶障害に与える影響 |
| L.plantarumTWK10が加齢に伴う空間学習および記憶力の低下に与える影響を評価するために、モリス水迷路 (MWM) テストを実施しました。2 元配置分散分析の結果、「加齢 (対照群 vs. 高齢者)」という因子が、それぞれ 1 日目、2 日目、3 日目に空間学習および記憶力に有意な影響を与えていることが示されました (P < 0.05)。高齢マウス(3日目のA-Control)の平均逃避潜時は、若年マウス(3日目のY-Control)の平均逃避潜時よりも有意に長かった(P < 0.01)。Y-TWK10群の2日目と3日目の平均逃避潜時は、1日目のY-TWK10群と比較して有意に短かった(それぞれP < 0.05、P < 0.01)のに対し、Y-Controlマウスではどの時点(1日目、2日目、3日目)でも有意差は認められなかった(図2A)。Y-TWK10群の1日目と3日目の平均逃避潜時の差は、Y-Controlマウスと比較して減少傾向(P = 0.1388)を示した(図2B)のに対し、A-TWK10群のそれはA-Control群よりも有意に短かった(P < 0.05)(図2C)。 |
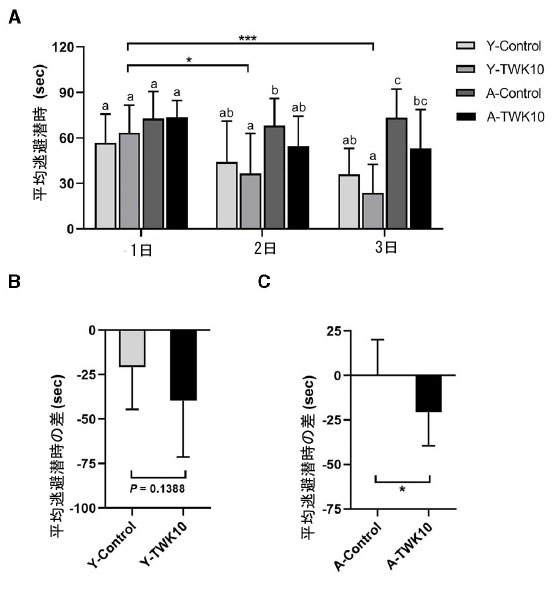 |
図2 空間学習および記憶力の低下は、Lactobacillus plantarum TWK10 によって緩和されました。L.plantarumTWK10または リン酸緩衝生理食塩水を 8 週間投与した後、空間学習および記憶能力をモリス水迷路 (MWM)を使用して評価しました。 (A) プラットフォームまでの平均逃避潜時は、試験 1、2、3 日目に記録されました。 (B、C) 1 日目と 3 日目の間の逃避潜時の差は、若いマウスと高齢マウスで調べられました。 データは平均 ± SD として表されます。試験日中の平均逃避潜時の群間の統計的差は、事後 Tukey-Kramer 検定による 2 元配置分散分析によって統計的に分析されました。異なる文字 (a、b、c) は群間の有意差を示します (P < 0.05)。2 群の比較は、対応のない Student t 検定を使用して分析されました。***P < 0.001。対照群とTWK10投与群間の逃避潜時のデルタタイムの差は、Mann-Whitney U検定を使用して分析されました。*P < 0.05。 |
| 3.4.L.plantarumTWK10の体脂肪調節効果 |
| A-Control群の平均腹部脂肪量 (%) は、Y-Control コントロール マウスと比較して減少傾向 (P = 0.1330) を示しました。L.plantarumTWK10を 8 週間投与した後、Y-TWK10 マウスの平均腹部脂肪量は Y-Control群よりも有意に低下 (P < 0.05) しましたが、高齢マウス群間では有意差は認められませんでした (図 3A、B)。有意な相互作用 (P < 0.05) [老化 (若齢 vs. 高齢) × 治療 (Control vs. L.plantarumTWK10投与)] は、それぞれ精巣上体脂肪体(EFP)重量の平均と精巣上体脂肪体(EFP)中の脂肪細胞の平均脂肪細胞の断面積 (CSA )で認められました。Y-TWK10 マウスの平均精巣上体脂肪体(EFP)重量と精巣上体脂肪体(EFP)中の脂肪細胞の平均脂肪細胞の断面積(CSA )は、Y-Control群よりも有意に低かった (P < 0.05) (図 3C~E)。 A-Control群の肩甲骨間褐色脂肪組織(BAT)脂肪細胞の平均数は、Y-Controlマウスと比較して差はありませんでした。しかし、L.plantarumTWK10投与により、A-TWK10 群ではControl群と比較して肩甲骨間褐色脂肪組織(BAT)脂肪細胞数が有意に増加しました (図 3F、G)。L.plantarumTWK10処理による肩甲骨間褐色脂肪組織(BAT)組成への影響をよりよく理解するために、ミトコンドリアを含む領域の割合として、相対的な肩甲骨間褐色脂肪組織(BAT)褐色領域 (%) (H&E 染色後に視覚化) を定量化しました。Y-TWK10 群と A-TWK10 群のマウスの肩甲骨間褐色脂肪組織(BAT)における平均相対褐色領域 (%) は、Control群と比較して大幅な増加傾向 (P = 0.0858) を示しました (図 3H)。 |
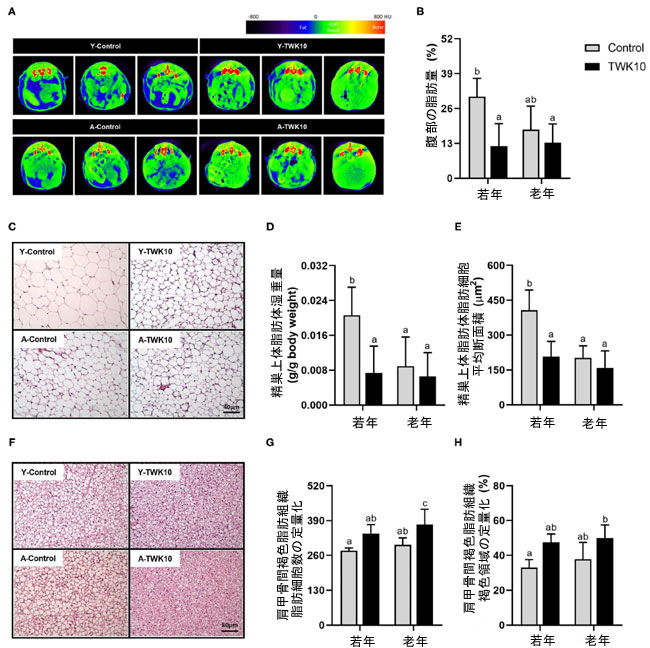 |
図3 Lactobacillus plantarumTWK10の体脂肪調節への影響。 (A) 腹部の構成を示す代表的なマイクロCT画像。 (B) 8週間のリン酸緩衝生理食塩水またはL.plantarumTWK10投与後の腹部L1~L5の脂肪量の定量化。精巣上体脂肪体(EFP)と肩甲骨間褐色脂肪組織(BAT)は、L.plantarumTWK10PBS投与8週間後に採取され、組織学的分析にかけられました。 (C) ヘマトキシリンおよびエオシン(H&E)染色によって得られた精巣上体脂肪体(EFP)の代表的な画像。 (D、E) 精巣上体脂肪体(EFP)湿重量および精巣上体脂肪体(EFP)脂肪細胞の平均断面積の測定。 (F) H&E染色によって得られた肩甲骨間褐色脂肪組織(BAT)の代表的な画像。 (G) 高倍率視野(HPF)あたりの肩甲骨間褐色脂肪組織(BAT)脂肪細胞数の定量化。 (H) H&E染色後の肩甲骨間褐色脂肪組織(BAT)褐色領域の定量化。 データは平均±SDとして表されます。群間の統計的差異は、事後Tukey-Kramer検定による2元配置分散分析によって分析され、異なる文字(a、b、c)は有意差を示します(P < 0.05)。ノンパラメトリックデータは、Dunn検定によるKruskal-Wallis検定によって統計的に分析されました。 |
| 3.5.L.plantarumTWK10の腸内細菌叢への影響 |
| L.plantarumTWK10の腸内細菌叢への影響を調べるため、L.plantarumTWK10/PBS 投与 8 週間後に排泄されたばかりの糞便サンプルを採取し、16S rRNA 遺伝子配列解析を行いました。糞便細菌叢の配列解析の結果、合計 5,314,481 の品質フィルタリングされたリードが得られました。これは、サンプルあたり平均 69,927 リードに相当します。キメラ除去後、サンプルあたりのリードは 52,190 に減少しました。配列解析データに基づいて、合計 580 の運用分類単位 (OTU) が得られ、267 属の細菌が特定されました。運用分類単位 (OTU)に関する分類および系統発生情報は、補足表 1 に記載されています。 |
| L.plantarumTWK10投与によって細菌組成の全体的なプロファイルがどのように変化するかを判断するために、アルファ多様性とベータ多様性を分析しました。α多様性のシャノン指数は、L.plantarumTWK10投与群と対照群の若いマウスおよび高齢マウスの間で有意差は見られませんでしたが、A-TWK10 群の豊富さ指数は Y-TWK10 群よりも有意に低かった (P < 0.05) (図 4A)。4 つの群間の細菌群集構造の違いを評価するために、重み付けなしおよび重み付け UniFrac 距離行列の非計量多次元尺度法 (NMDS) 順序付けと PERMANOVA によってベータ多様性を測定しました。図 4B に示すように、分類群の有無 (加重なし UniFrac 距離) を考慮した NMDS 分析では、Y-Control群の糞便細菌組成は A-Control群とは有意に (PERMANOVA、P < 0.05) 異なっていました。分類群の有無と相対的存在量 (加重 UniFrac 距離) を考慮すると、Y-TWK10 マウスは Y-Control マウスと比較して明らかに異なる (PERMANOVA、P < 0.05) β 多様性プロファイルを示し、L.plantarumTWK10が若いマウスの腸内細菌組成の再編成を誘発したことを示唆しています。しかし、A-TWK10 マウスは、加重 UniFrac 距離 (P = 0.309) で NMDS によって決定された A -Control群と同様の細菌組成プロファイルを示しました。 |
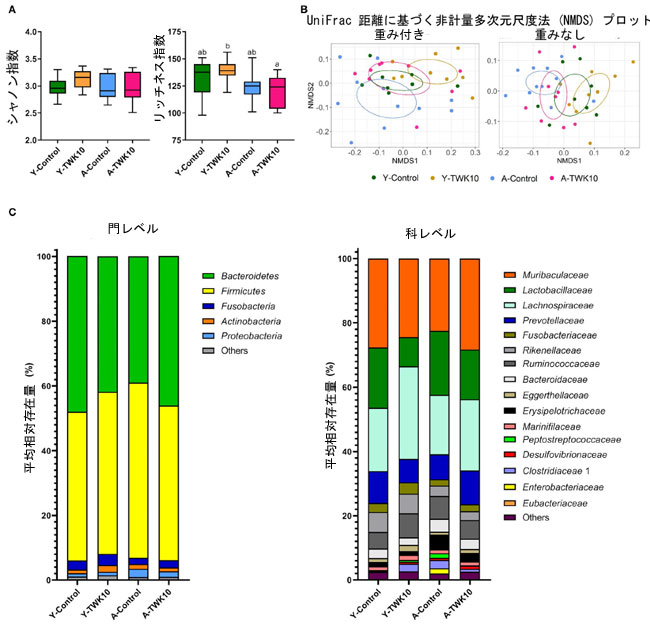 |
図4 Lactobacillus plantarum TWK10 による腸内細菌組成の調整。 腸内細菌叢の特徴は、 (A) アルファ多様性 (シャノン指数とリッチネス指数) で表されました。箱ひげ図は、最小値と最大値、25% 四分位値と 75% 四分位値、および中央値を示します。 (B) ベータ多様性は、重み付きおよび重みなし UniFrac 距離に基づく非計量多次元尺度法 (NMDS) プロットで表され、 (C) 門および科レベルでの細菌分類群の平均相対存在量の累積棒グラフです。 群間の統計的差異は、事後 Tukey-Kramer 検定による 2 元配置分散分析によって分析され、異なる文字 (a、b、c) は群間の有意差を示します (P < 0.05)。ノンパラメトリック データは、Dunn 検定による Kruskal-Wallis 検定によって統計的に分析されました。 |
| 細菌組成のこれらの変化をさらに調査するため、門および科レベルでの主要分類群の相対的存在量を分析しました。門レベルでは、A-TWK10 群は A-Control マウスと比較して、Bacteroidetes 門およびFusobacteriaの量は有意に高くないもののわずかに高く、Firmicutes 門およびProteobacteria の量はわずかに低くなりました。さらに、L.plantarumTWK10投与により、老齢マウスのFirmicutes/Bacteroidetes (F/B) 比が低下する傾向がありました (補足図 1A)。Y-TWK10 群の放線菌の量は、Y-Controlマウスよりも有意に高くなりました (P < 0.05) (図 4C、補足図 1A)。科レベルでは、Y-TWK10 群の Lachnospiraceae および Eggerthellaceae の相対存在量は Y-Control マウスと比較して有意に高かった (P < 0.05) のに対し、Y-TWK10 群の Lactobacillaceae の存在量は Y-Control マウスと比較して有意に低かった (P < 0.01)。Y-TWK10 群の Eubacteriaceae (P = 0.0823) および Ruminococcaceae (P = 0.0927)、A-TWK10 群の Muribaculaceae (P = 0.1119) および Eggerthellaceae (P = 0.1308) の存在量は、それぞれ Y-Control マウスおよび A-Control マウスのこれらの集団の存在量と比較した場合、有意に向かって弱い増加傾向を示した (補足図 1B)。 |
| 各マウス群内の特定の特徴的分類群を分析するために、群間の相違に基づいて LEfSe [線形判別分析 (LDA) 効果サイズ] を実行しました。Y-TWK10 群に豊富に含まれる系統型には、Actinobacteria 門と主にClostridia綱に属する属、すなわち Lachnospiraceae UCG-006、Lachnoclostridium、Phascolarctobacterium、Ruminiclostridium 6、Tyzzerella、Eubacterium brachy群、および Ruminococcaceae UCG-004 が含まれ、Y-Control 群の系統型はBacilli 綱に属していました (補足図 2A)。 A-TWK10マウスでは、Bacteroidetes 門が最も差次的に豊富な細菌分類群であったのに対し、A-Control群の細菌叢はFirmicutes 門(LDAスコア[log10]> 3)が優勢であった(補足図2B)。 |
| 3.6.L.plantarumTWK10の腸内細菌ネットワークへの影響 |
| 若齢マウスと高齢マウスの両方に L.plantarumTWK10を投与した後、腸内細菌群集における細菌相互作用の変化を調べるために、SparCC を使用して微生物相関ネットワーク分析を実施しました。Y-Controlマウスでは、Prevotellaceae、Lactobacillaceae、および Ruminococcaceae が、それぞれ Bacteroidaceae、Burkholderiaceae、Anaeroplasmataceae、および Staphylococccaceae と正の相関を示しました (図 5A)。Y-TWK10群は、優勢な Ruminococcaceae が Anaeroplasmataceae、Enterobacteriaceae、および Enterococcaceae と負の相関を示し、Clostridiales vadinBB60群と正の相関を示しました。Eggerthellaceae は Fusobacteriaceae と正の相関を示しました (図 5B)。 A-Control群では、最も優勢な Lachnospiraceae は、Enterobacteriaceae および Enterococcaceae と負の相関を示し、Peptococcaceae と正の相関を示しました。Chitinibacteraceae、Fusobacteriaceae、および Rhodocyclaceae の 3 つの科は、互いに正の相関を示しました (図 5C)。A-TWK10 マウスでは、Lachnospiraceae は、Desulfurvibrionaceae、Lactobacillaceae、Muribaculaceae、Prevotellaceae、Ruminococcaceae、および Streptococcaceae と負の相関を示しました。Ruminococcaceae は、Chitinibacteraceae、Prevotellaceae、Staphylococcaceae、Streptococccaceae、および Lachnospiraceae と負の相関を示し、Clostridiales vadinBB60 群および Desulfurvibrionaceae と正の相関を示しました。 Muribaculaceae は Streptococcaceae と正の相関を示し、Christensenellaceae とは負の相関を示しました。Chitinibacteraceae、Chromatiaceae、Fusobacteriaceae、および Rhodocyclaceae の 4 つの科は、互いに正の相関を示しました。そのうち 3 つ (Chitinibacteraceae、Fusobacteriaceae、および Rhodocyclaceae) は、Christensenellaceae とも負の相関を示しました (図 5D)。 |
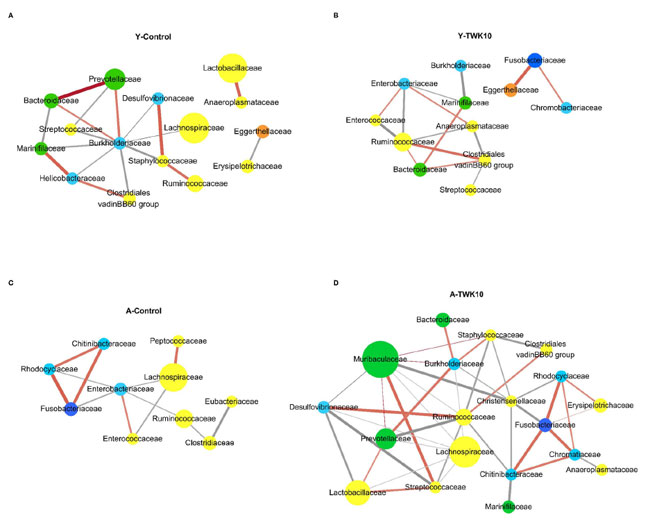 |
図5 腸内細菌叢の共起ネットワーク分析。 共起ネットワークは、 (A) 若いコントロール マウス (Y-Control)、 (B) 若い L.plantarumTWK10投与マウス (Y-TWK10)、 (C) 高齢コントロール マウス (A-Control)、 (D) 高齢L.plantarumTWK10投与マウス (A-TWK10) の相対的存在量に基づいてファミリー レベルで実行されました。 接続は、強い (SParCC の rho カットオフ = 0.6) かつ有意な (P < 0.01) 相関を示しています。各ノードはファミリーを表し、門に従って色分けされています。 黄色: Firmicutes、緑: Bacteroidetes、水色: Proteobacteria、濃青: Fusobacteria、オレンジ: Actinobacteria。 ノードのサイズは、各群内の各ファミリーの相対的存在量 (> 1% の場合) を表します。各エッジは、2 つのファミリー間の正の相関 (オレンジと赤の線) または負の相関 (灰色の線) を表します。SparCC の相関係数は ±0.6 で、エッジの陰影は相関の大きさを示します。 |
| 3.7.L.plantarumTWK10が 短鎖脂肪酸産生腸内細菌の豊富さと腸内短鎖脂肪酸レベルに与える影響 |
| 酢酸、プロピオン酸、酪酸などの短鎖脂肪酸は、腸の恒常性維持に重要な代謝物です。そこで次に、L.plantarumTWK10が若いマウスと高齢マウスの腸内細菌叢の調整を通じて短鎖脂肪酸の産生を促進できるかどうかを調べました。Y-TWK10 マウスの盲腸内容物中の酢酸、プロピオン酸、酪酸の濃度は、Y-Control群と比較して有意に (P < 0.05) 高かったのに対し、A-Control群と A-TWK10群の間には有意差は認められませんでした。 |
| 若いマウスでは、L.plantarumTWK10を投与すると短鎖脂肪酸産生細菌の存在量が増えることがわかりました。酢酸産生細菌の場合、Peptococcaceae 科 (P < 0.01) および Ruminococcaceae 属 UCG-004 (P < 0.01) の存在量が有意に多く、Y-TWK10 マウスでは Y-Control群と比較して Prevotella 9 属 (P = 0.1111)、Ruminococcaceae 6 属 (P = 0.0755)、および Ruminiclostridium 9 属 (P = 0.0745) の強い増加傾向が見られました。 TWK10 投与の結果、酪酸産生細菌、すなわち Lachnospiraceae 科 (P < 0.05) および Lachnospiraceae 属 UCG-006 (P < 0.001) の存在量が Y-TWK10 マウスでは Y-Control 群と比較して有意に増加し、Ruminococcaceae 科 (P = 0.0927) および Eubacterium 属 (P = 0.1388) および Roseburia 属 (P = 0.1388) の存在量についても有意な増加傾向が認められました。高齢マウスにL.plantarumTWK10を投与すると、有意ではないものの、特に Prevotellaceae 属 UCG-001 に属する酢酸産生細菌の存在量が増加しました (P = 0.1519)。 A-TWK10 群と対照群のマウスにおける酪酸産生細菌の豊富さには有意差はありませんでした。さらに、プロピオン酸産生細菌 Phascolarctobacterium 属は Y-TWK10 群と A-TWK10 群の両方で観察されました (表 3)。 |
| 表 3. Lactobacillus plantarum TWK10 は短鎖脂肪酸産生腸内細菌を増強し、短鎖脂肪酸の産生を促進しました |
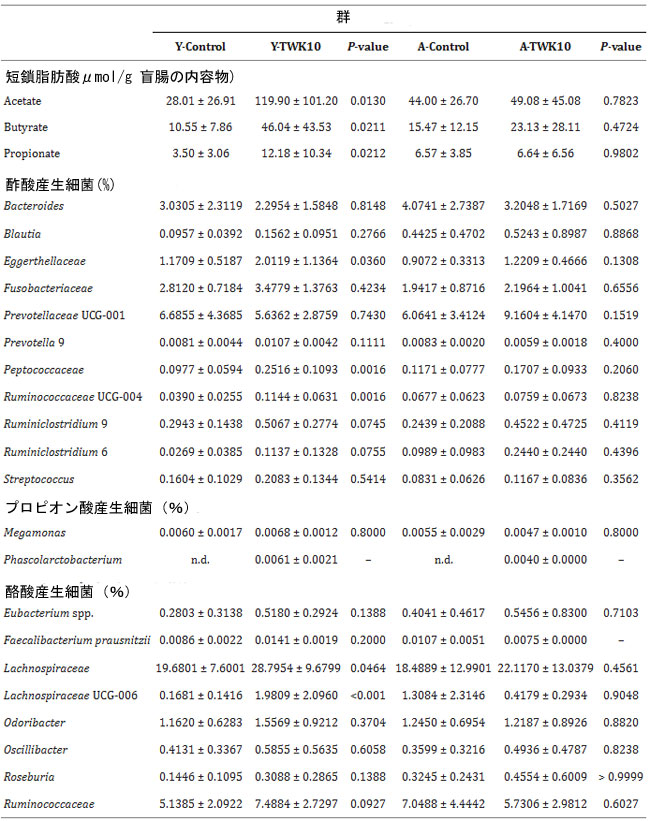 |
| 若いマウスと老齢マウスにおける短鎖脂肪酸 関連腸内細菌の相対的存在量と盲腸短鎖脂肪酸含有量。データは平均 ± SD として表されます。短鎖脂肪酸産生腸内細菌の相対的存在量に基づくControl群とL.plantarumTWK10投与群間の統計的差異は、Mann-Whiney U 検定によって分析されました。Control群とL.plantarumTWK10 TWK10 投与群の盲腸短鎖脂肪酸含有量間の統計的有意性は、Student の無対 t 検定によって分析されました。P < 0.05 は統計的に有意な差異を表します。n.d:検出されませんでした。 |
| 3.8.腸内細菌叢の構成とL.plantarumTWK10による健康効果の相関関係 |
| 若いマウスと高齢マウスにおける、腸内細菌分類群の相対的存在量と、空間学習と記憶、筋肉と骨の質を含む 7 つの加齢関連宿主表現型特性との相関関係を、スピアマンの相関分析を使用して評価しました。高齢マウスをL.plantarumTWK10で治療した後、逃避潜時は有意に短縮しました (A-TWK10 群、図 2C)。逃避潜時の変化は、Enterobacteriaceaeの存在量と有意かつ正の相関を示しました (図 6)。一方、A-Controlマウスにおける Enterococcaceae および Prevotellaceae 集団の存在量は、逃避潜時の変化と負の相関を示しました。若いマウスでは、Chitinibacteriaceae, Fusobacteriaceae, およびRhodocyclaceae (Y-Control 群) の存在量は、逃避潜時の変化と正の相関を示しました。 Y-TWK10 マウスへのL.plantarumTWK10投与による腹部脂肪量 (%) の減少は、それぞれ Prevotellaceae、Desulfurovibrionaceae、Staphylococccaceae の存在と有意に正および負の相関を示した。一方、Enterococcaceae の豊富さは A-TWK10 マウスの脂肪量と正の相関を示した。Y-Control 群では、Clostridiaceae と Erysipelotrichaceae、Enterococcaceae の豊富さがそれぞれ脂肪量と正および負の相関を示したが、Lachnospiraceae の豊富さは A-Control マウスの脂肪量と正の相関を示した。筋肉の老化に関連するパラメータについては、Y-Control 群では Burkholderiaceae の豊富さが握力と正の相関を示したが、Helicobacteraceae、Lachnospiraceae、および Marinifilaceae の個体群は握力と負の相関を示した。高齢マウスでは、A-Control群ではErysipelotrichaceaeの存在が握力と正の相関関係にあったのに対し、A-TWK10またはY-TWK10マウスでは有意な相関関係は見られませんでした。さらに、筋肉グリコーゲンレベルは、若年マウスと高齢マウスの両方でL.plantarumTWK10投与によって増加し(図1D)、Lactobacillaceaeの存在と正の相関関係がありました。Y-Control群では、BacteroidaceaeとPrevotellaceaeの存在が筋肉グリコーゲンレベルと正の相関関係にあったのに対し、A-Control群では有意な相関関係は見られませんでした(図6)。A-TWK10マウスでは、Proteobacteria門の存在と筋肉グリコーゲンレベルの間に強い負の相関関係も見られました(補足図3)。骨の健康パラメータに関しては、L.plantarumTWK10投与後、若年群と高齢群で骨梁数(Tb.N)と骨容積率(BV/TV)値が高くなりました(表2)。 Y-TWK10マウスの大腿骨では、Enterobacteriaceaeが骨梁数(Tb.N)と正の相関を示した。高齢マウス群では、骨梁数(Tb.N)と強い相関を示す細菌科はなかった。しかし、Y-TWK10マウスでは、Enterobacteriaceae と Peptostreptococcaceaeの存在が骨容積率(BV/TV)と正の相関を示したのに対し、Y-Control群では、Fusobacteriaceae と Streptococcaceaeの集団が骨容積率(BV/TV)と負の相関を示した。A-TWK10の血清ビタミンDレベルも、A-Controlマウスと比較して高かった(P = 0.0584)(表2)。しかし、A-TWK10群では、細菌分類群と血清ビタミンDレベルの間に有意な相関は見られなかった。 A-Controlマウスでは、Clostridiales vadinBB60 群のメンバー、Eggerthellaceae、Helicobacteraceae、Marinifilaceae の豊富さが血清ビタミン D レベルと正の相関を示しましたが、Erysipelotrichaceae と Peptostreptococcaceae は負の相関を示しました。Y-Controlマウスでは、Lactobacillaceae の豊富さが血清ビタミン D レベルと負の相関を示しました。 |
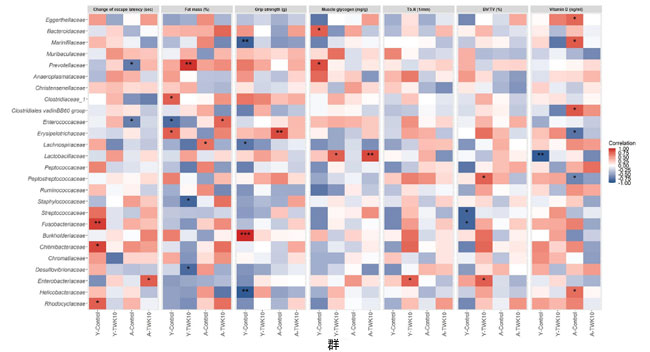 |
図6 腸内細菌叢と加齢に伴う宿主表現型特性の変化とのスピアマン相関分析のヒートマップ。 スピアマン相関分析は、26 の主要なファミリーの相対的存在量と、Y-Control と Y-TWK10 または A-Control と A-TWK10群間の差分分析から選択された 7 つの加齢に伴う宿主表現型特性の値との相関関係を調べるために実施しました。 赤い四角は正の相関関係を示し、青い四角は負の相関関係を示します。*P < 0.05、**P < 0.01、***P < 0.001。 |
| 4.考察 |
| 4.1.L.plantarumTWK10の筋力に対する効果 |
| 加齢により、骨格筋の同化障害が引き起こされ、その結果、筋肉量と筋力が低下します (45)。筋肉量と筋力の低下は、高齢者の死亡率と密接に関連していることを示す証拠が増えています。この 2 つのうち、高齢者の機能的制限と健康状態の悪化の決定要因としては、筋力の方が筋肉量よりも重要であるようです (2、3、46~49)。グリコーゲンは、ヒトと動物の骨格筋活動を支える重要なエネルギー基質であり、筋肉のグリコーゲン生成率は加齢とともに低下します (50)。これまでの研究で実証されているように、L.plantarumTWK10の投与により、マウスとヒトの筋力が強化され、筋肉量が増加し、筋肉の質と持久力が向上しました (30、31)。しかし、筋肉の老化の進行を遅らせるL.plantarumTWK10の影響は明らかではありませんでした。 |
| 本研究では、自然に老化するマウスの筋力低下などの老化関連疾患の進行に対するL.plantarumTWK10の経口投与の効果を実証することを目的とした。そのため、若いマウスと高齢マウスの前肢の握力(生体内神経筋パフォーマンスの評価に広く使用されるマーカー)、腓腹筋の筋肉重量、および筋肉グリコーゲンレベルを測定した。 |
| 結果が示すように、高齢マウスでは若年マウスと比較して前肢の握力 (P < 0.001)、筋肉重量 (P < 0.05)、およびグリコーゲンレベル (P < 0.05) の有意な減少が観察されました (図 1A~D)。L.plantarumTWK10投与により、若年マウスと高齢マウスのどちらでも、対照群と比較して筋肉重量が有意に増加したようには見えませんでしたが、握力と筋肉グリコーゲンレベルは有意に増加しました。さらに、若年マウス (P < 0.01) と高齢マウス (P < 0.05) の両方で、握力と筋肉グリコーゲンレベルの間に有意な正の相関が観察されました (図 1E)。これらの結果は、L.plantarumTWK10によって発揮される筋力改善効果が、特に筋肉組織内のグリコーゲン濃度の増加による筋肉の質の改善に起因する可能性があることを示しています。 L.plantarumTWK10が老齢マウスの筋力を改善し、筋肉の減少を防ぐ効果は、L. paracasei PS23 を老化促進マウス傾向 8 (SAMP8) マウスに使用した以前の研究と一致しています (27)。以前の研究では、L.plantarumTWK10投与により 6 週齢のマウスの筋肉量を増やすことができることがわかりました (30)。しかし、本研究では、L.plantarumTWK10投与によって筋肉量は増加しませんでした (図 1C)。ただし、L.plantarumTWK10を投与された若いマウスでは筋線維の断面積が拡大しました (データ未掲載、Y-Control vs. Y-TWK10群、531 ± 73 vs. 641 ± 92 μm2)。筋肉量に関するこれらの矛盾した結果は、研究対象動物の年齢の違いによるものと考えられます。一般に、3~6 か月齢のマウスは成熟した成体とみなされ、老化関連の研究では通常参照群として使用されます。マウスは生後2~3か月まで急速に成長するので、発達段階でL.plantarumTWK10を投与すると、成熟した成体段階よりも成長を促進しやすくなる可能性があります。 |
| 筋肉のグリコーゲン含有量は、ミトコンドリアの質、食事の栄養、炎症関連のサイトカインや同化ホルモンのレベルとともに、筋肉の量と質に影響を及ぼします。上記の結果から、L.plantarumTWK10の投与により老齢マウスの筋力が著しく改善したことが確認されましたが、その根底にあるメカニズムを解明するにはさらなる調査が必要です。さらに、Khandelwal ら (51) は、老齢ラットの肝臓グリコーゲン合成酵素とホスホリラーゼの活性低下は、加齢に伴う肝臓のグリコーゲンの代謝回転の低下を示していると報告しました。筋肉のグリコーゲン合成は加齢とともに低下します (50)。骨格筋は、インスリンが GLUT4 の転座を介して血液からのグルコースの取り込みを刺激する主要な組織です (52)。したがって、L.plantarumTWK10は、加齢に伴うグリコーゲン代謝の低下の調節に役割を果たし、加齢とともに一般的に観察される耐糖能障害に寄与している可能性があります。 |
| 4.2.L.plantarumTWK10の骨質への影響 |
| 骨粗鬆症は、高齢者の骨量と骨密度の減少を特徴とする一般的な加齢性疾患です (6)。加齢または骨粗鬆症の発症に伴い、骨容積率(BV/TV)、骨梁厚(Tb.Th)、骨梁数(Tb.N)、骨密度(BMD) は減少しますが、骨梁間隔(Tb.Sp)、骨梁パターン係数(Tb.Pf)、構造モデル指数(SMI)は通常増加します (53)。ビタミン D はカルシウム代謝に関与する必須因子であるため、骨形成に必要です。加齢に伴い、ビタミン D 欠乏のリスクも大幅に増加します (54)。A-Controlマウスでは、Y-Control群と比較して、骨梁数(Tb.N)の大幅な減少 (P < 0.05) と、骨梁間隔(Tb.Sp) のわずかな増加 (P = 0.1547) が検出され、加齢に伴う骨量減少が観察されました。 L.plantarumTWK10投与により、若年マウスと高齢マウスの両方で骨容積率(BV/TV)と 骨梁数(Tb.N)値が有意に増加 (P < 0.05) し、若年マウスの骨梁間隔(Tb.Sp)が有意に減少 (P < 0.05) しました。血清ビタミン D レベルのほぼ有意な増加 (P = 0.0584) も A-TWK10 マウスで観察されました (表 2)。これらの結果は、L.plantarumTWK10投与が加齢に伴う骨量減少に対する保護効果を示しただけでなく、マウスの骨質も改善したことを示しています。これは、卵巣摘出 SAMP8 マウスにおける L. paracasei GKS6 および L. plantarum KM3 補給の効果を調査した以前の研究と一致しています (29)。 |
| 4.3.L.plantarumTWK10の体脂肪に対する効果 |
| 一般的に、加齢は除脂肪体重の減少をもたらすだけでなく、体脂肪量の増加をもたらします (55)。加齢とともに、脂肪は皮下から内臓領域に再分配されます。腹部区画における脂肪組織の蓄積は、心血管疾患、インスリン抵抗性、2 型糖尿病などの慢性疾患のリスク増加と関連しています (56、57)。ここでは、マイクロ CT によって、加齢に伴う体組成の変化と L.plantarumTWK10の体脂肪に対する影響を評価しました。L.plantarumTWK10投与により、若いマウスの平均腹部脂肪量、平均精巣上体脂肪体(EFP)重量、および精巣上体脂肪体(EFP)脂肪細胞の平均 脂肪細胞の断面積(CSA )が Y-Control群と比較して有意に低下 (P < 0.05) することがわかりました (図 3A–E)。これは、以前の調査結果 (30) と一致しています。 A-Control 群では、Y-Control マウスと比較して、加齢に伴う体脂肪の増加が見られると予想していましたが、脂肪量に有意な変化は見られませんでした。Hamrick ら (58) が以前に報告したように、この好ましくない結果は、加齢マウスで自然に起こる脂肪量の大幅な減少によるものと考えられます。本研究で使用した高齢マウスは、脂肪量評価時に 27~30 か月齢であったため、L.plantarumTWK10が加齢に伴う体脂肪の蓄積に与える影響を調べるには高齢すぎた可能性があります。また、加齢は肩甲骨間褐色脂肪組織(BAT)量と活性の低下と関連しています (59)。加齢に伴う肩甲骨間褐色脂肪組織(BAT)の減少は、ミトコンドリア機能の低下、炎症の増加、交感神経系の障害、および内分泌シグナル伝達の変化に関連しています (60~63)。当社のデータによると、Y-Control マウスと A-Control マウスを比較した場合、肩甲骨間褐色脂肪組織(BAT)に影響を与える有意な変化は検出されませんでした。しかし、L.plantarumTWK10の投与により、Y-TWK10 マウスと A-TWK10 マウスの両方で平均脂肪細胞数とミトコンドリアを含む領域のサイズが増加しました (図 3F~H)。これは、L.plantarumTWK10が脂肪組織の褐色化を促進する可能性があることを示唆しています。私たちの知る限りでは、肩甲骨間褐色脂肪組織(BAT)は熱生成とエネルギー消費に不可欠です。したがって、L.plantarumTWK10が体脂肪に与える影響を明らかにするために、肩甲骨間褐色脂肪組織(BAT)の活性と褐色脂肪と白色脂肪の比率をさらに評価する価値があります。これらの結果は、L.plantarumTWK10 が体組成を変え、より健康的な体型を促進し、代謝を改善する可能性があることを示しています。 |
| 4.4.L.plantarumTWK10の学習と記憶に対する効果 |
| 学習と記憶の障害は、高齢者によく見られるもう 1 つの健康問題です。げっ歯類の空間学習と記憶を評価するモリス水迷路 (MWM) は、最も広く使用されている行動テストの 1 つです (35)。私たちは、マウスの加齢に伴う空間学習と記憶の低下に対する L.plantarumTWK10TWK10 の効果を評価するために、モリス水迷路 (MWM)テストを実施しました。 |
| 逃避潜時は加齢とともに有意に増加(P < 0.001)しましたが、L.plantarumTWK10投与により、若いマウスでは 2 日目と 3 日目に逃避潜時が有意に短縮しました(図 2A)。A-TWK10 マウスと Y-TWK10 マウスの 1 日目と 3 日目の間の逃避潜時の変化は、それぞれ有意な減少(P < 0.05)と減少傾向(P = 0.1388)を示しました(図 2B、C)。これらの結果は、L.plantarumTWK10が若いマウスの学習能力と記憶能力を改善し、高齢マウスの記憶喪失を軽減したことを示しています。これらの観察結果は、SAMP8 マウスにおける L. paracasei PS23 (26)、D-ガラクトース誘発老化 C57BL/6 マウスにおける L. paracasei BCRC 12188、L. plantarum BCRC 12251、および Streptococcus thermophilus BCRC 13869 の混合プロバイオティクス (64)、および SAMP8 マウスにおける Bifidobacterium bifidum、Bifidobacterium lactis、L. acidophilus、および Lactobacillus casei からなる多菌株プロバイオティクス製剤 (65) などの以前の研究と一致しています。 |
| 4.5.L.plantarumTWK10の腸内細菌叢に対する効果 |
| 加齢に伴う変性は、通常、腸内細菌叢の不均衡と関連しています (9、11、66~68)。私たちの結果は、加齢がマウスの腸内細菌叢を変化させることを示す以前の研究結果と一致しています (69)。A-Control群マウスの糞便微生物構造は、Y-Control群と比較して、Firmicutes に富み、Bacteroidetes の豊富さが低かった (補足図 1A)。Firmicutes の大きな集団と Bacteroidetes の小さな集団は、一般的に、異常な細菌叢の特徴(dysbiotic microbial signature)と健康状態の悪さと関連しています (70、71)。プロバイオティクスは、腸内細菌組成の不均衡を調整し、腸内細菌群に有益な機能を与えます (72)。我々の研究結果は、L.plantarumTWK10投与により、A-Control群と比較して、高齢マウスの Firmicutes/Bacteroidetes (F/B) 比が減少したことを示しています (補足図 1A)。ただし、この減少は統計的に有意ではなく、これは研究で使用したサンプル サイズが小さかったためである可能性があります。この結果は、L.plantarumTWK10投与によりFirmicutes/Bacteroidetes (F/B)比が修正され、腸内細菌バランスが回復し、健康上の利点が得られる可能性があることを示唆しています。 |
| 老齢マウスでは、若齢マウスに比べてProteobacteria門およびEnterobacteriaceae 科のメンバーが多く存在していました (補足図 1A、B)。Proteobacteria門は、Enterobacteriaceae 科、Pseudomonadaceae科、 Vibrionaceae科、Yersiniaceae科など、さまざまなグラム陰性病原体で構成されています。その中でも、Enterobacteriaceae科は 30 属 130 種を超える中核細菌群です。これらの腸内細菌には、病原性の可能性のある細菌 (病原性細菌と呼ばれる) が含まれており、これは宿主の粘膜免疫系に特定の影響を及ぼし、加齢により宿主の抵抗力が失われた場合に感染症の主な原因となります (73、74)。さらに、Enterobacteriaceae科やその他のグラム陰性細菌の加齢に伴う増加は、弱くなった腸管バリアに対するエンドトキシン攻撃の増加につながり、慢性炎症につながる可能性がある(74)。私たちの研究結果は、L.plantarumTWK10が高齢マウスのEnterobacteriaceae科の存在量を減らすことができたことを示している(補足図1B)。L.plantarumTWK10投与後、若いマウスと高齢マウスで、Chitinibacteraceae科、Enterobacteriaceae科、Helicobacteraceae科、Rhodobacteraceae科、Rhodocyclaceae科(Burkholderiaceae科を除く)など、Proteobacteria門の主要な科の存在量の大幅な減少が観察された(補足図4)。これらの結果は、L.plantarumTWK10が高齢者の腸の健康を促進する可能性があることを示唆した以前の報告(75、76)と一致している。 |
| さらに、若いマウスと高齢マウスの間で微生物群集の共起パターンに違いがあることを発見しました。共起ネットワークは、Bacteroidetes門に属する微生物がY-Control群で強い正の相関関係にあることを示しました(図5A)。一方、A-Controlマウスでは、Proteobacteri門(Chitinibacteraceae科、Enterobacteriaceae 科、Rhodobacteracea科)に属するグラム陰性日和見病原体とEnterococcaceae科が互いに正の相関関係にありました(図5C)。Y-TWK10群では、L.plantarumTWK10投与後に共起ネットワークのトポロジーが変化しました。Y-TWK10群のメインネットワークで最も豊富な細菌科であるRuminococcaceaeは、Anaeroplasmataceae、Enterobacteriaceae、Enterococcaceaeと相互に制限を示しました(図5B)。代謝の健康と長寿に関連するChristensenellaceae科(77~80)は、Chitinibacteraceae、Fusobacteriaceae、Rhodocyclaceaeと負の相関関係を示しました。さらに、病原微生物間のつながりが老齢マウスへのL.plantarumTWK10投与によって破壊されることもわかりました(図5D)。これは、病原微生物間の相関相互作用の破壊が老化プロセスを遅らせる潜在的な用途を持つ可能性があることを示唆しています。 |
| 腸内細菌叢由来の短鎖脂肪酸には、酢酸、プロピオン酸、酪酸などがあり、腸と代謝の健康維持に重要なメディエーターです (20、81、82)。微生物による短鎖脂肪酸の産生増加は健康に有益であると考えられています。乳酸は、糖発酵の主な最終産物として乳酸菌によって産生され、短鎖脂肪酸産生腸内細菌の重要な成長因子の 1 つでもあります (83)。そのため、乳酸菌はプロバイオティクスとして広く使用されています (84、85)。Chen ら (86) は、L. paracasei PS23 の投与により、老齢の SAMP8 マウスにおける酪酸産生 Lachnospiraceae UCG 001 の存在量が大幅に減少したと報告しました。対照的に、L.plantarumTWK10投与により、Y-TWK10 マウスでは Lachnospiraceae の存在量が有意に増加し (P = 0.0464)、A-TWK10群では高いレベル (総糞便微生物叢の 20% 以上を占める) で維持されることが観察されました。Wang ら (87) は、プロバイオティクスの L. plantarum P-8 が、高齢者を含むすべての年齢層で糞便中の酢酸とプロピオン酸の濃度を高めることで、ヒトの胃腸の健康を改善できることを報告しました。さらに、プロバイオティクスの L. acidophilus DDS-1 は、Akkermansia muciniphila と Lactobacillus spp. を増加させ、Proteobacteria spp. の存在量を減少させることで腸内細菌叢を調整し、高齢の C57BL/6J マウスの盲腸のプロピオン酸と酪酸レベルの上昇を伴いました (21、22)。 L.plantarumTWK10の投与は若いマウスと老齢マウスの腸内細菌叢を調節すると思われることから、L.plantarumTWK10は腸内短鎖脂肪酸産生を高めるために使用できるという仮説を立てました。その結果、Y-Control群と A -Control群の間で盲腸 短鎖脂肪酸レベルに有意差は見られませんでしたが、Y-TWK10 群では Y-Controlマウスと比較して 短鎖脂肪酸レベルの有意な増加が観察されました (表 3)。Y-TWK10群では、Y-Controlマウスと比較して酢酸および酪酸産生細菌の全体的な増加が観察されました。また、L.plantarumTWK10の投与により老齢マウスの腸内酢酸および酪酸レベルがわずかに上昇することも観察されました。これらの結果は、L.plantarumTWK10が特に若いマウスにおいて、細菌叢の調節、ひいては短鎖脂肪酸産生を介して健康上の利点を発揮する可能性があることを示唆しています。 |
| 我々は以前の研究で、L.plantarumTWK10を投与するとマウスの骨格筋量と筋力が著しく増加することを実証しました (30)。増え続ける証拠から「腸-筋軸」という概念が生まれ、腸内細菌叢が筋肉の健康の媒介として機能し (88、89)、サルコペニアなどの筋肉関連疾患の治療に期待が持たれていることが示唆されています (90)。多くの研究で、筋肉のグリコーゲン濃度と疲労耐性の間には密接な関係があることが強調されています (91)。しかし、筋肉のグリコーゲン濃度と筋肉機能を結び付けるメカニズムは依然として不明です。短鎖脂肪酸は骨格筋のグリコーゲン濃度を増加させることが示されており (92~95)、骨格筋量と身体機能を増加させる可能性を秘めています (96)。本研究では、若いマウス(P = 0.007)と高齢マウス(P = 0.019)の筋力と筋グリコーゲンレベルが有意に正の相関関係にあることがわかりました(図1E)。また、Y-TWK10マウスとA-TWK10マウスでは、乳酸菌類の存在量が筋グリコーゲンレベルと有意な正の相関関係を示しました(図6)。また、各治療群における加齢に伴う宿主表現型の特徴の変化が短鎖脂肪酸レベルと相関しているかどうかも評価しました。しかし、これらの相関関係のうち統計的に有意なものはごくわずかでした(P < 0.05)。若いマウス群では、L.plantarumTWK10投与後、筋グリコーゲン濃度は短鎖脂肪酸レベルと正の相関関係を示しました。さらに、Y対照群では、酪酸と筋グリコーゲンレベルの間に負の相関関係が見られました(補足図5)。これらの相関関係がL.plantarumTWK10投与の結果として生じたかどうかを判断することは依然として重要です。腸内細菌叢によって生成された短鎖脂肪酸が L.plantarumTWK10 投与後に骨格筋に影響を及ぼすメカニズムを理解するには、さらなる研究が必要です。 |
| 5.結論 |
| 本研究では、L.plantarumTWK10が自然に老化するマウスの老化関連疾患を軽減するという証拠を初めて示しました。L.plantarumTWK10の投与により、筋肉の質が向上し、筋肉グリコーゲン レベルが上昇することで、老化関連の筋力低下が軽減されることが分かりました。L.plantarumTWK10は、マウスの大腿骨における加齢関連の骨量減少の進行を遅らせ、骨梁数を増加させました。L.plantarumTWK10投与はまた、モリス水迷路 (MWM)テストの平均逃避潜時を短縮することで、高齢マウスの認知機能障害を大幅に軽減しました。さらに、16S rRNA 遺伝子の次世代シーケンシング (NGS) による腸内細菌叢分析により、L.plantarumTWK10 投与後に腸内細菌組成が大幅に変化し、加齢とともに発生する病原性微生物 (Enterobacteriaceae と Enterococcaceaeなど) の自然蓄積が減少し、有益な短鎖脂肪酸産生細菌の量が増加することが明らかになりました。 |
| 結論として、Lactobacillus plantarum TWK10は、老化関連疾患を軽減し、腸内細菌叢の不均衡を調整することで、健康的な老化を促進する潜在的な治療薬として考えられることが確認されました。L.plantarumTWK10が人間に健康を促進する効果を確認する前に、臨床現場で動物モデルを使用して得られた結果を検証するための今後の研究が必要です。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
| この文献は、Front Nutr. 2021 Oct 13;8:708096. に掲載されたLactobacillus plantarum TWK10 Attenuates Aging-Associated Muscle Weakness, Bone Loss, and Cognitive Impairment by Modulating the Gut Microbiome in Mice.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |