Patrycja Lipska al., |
要約 |
| アトピー性皮膚炎(AD)は、複雑な病因・病態生理を持つ慢性炎症性皮膚疾患であり、広範な研究にもかかわらず、その病態生理は依然として完全には解明されていません。この疾患は世界人口のかなりの割合に影響を与え、重大な臨床的負担となっています。近年、アトピー性皮膚炎を含む炎症性疾患の病態生理を調節する可能性のある因子として、腸内細菌叢への注目が高まっています。本レビューの目的は、腸内細菌叢の異常とアトピー性皮膚炎で観察される炎症過程の悪化との関連性に関する最新の知見を批判的に検討することです。これまでの研究から、腸内細菌叢の構成変化は、腸皮膚軸の調節異常、腸管バリア透過性の亢進、および炎症促進機構の活性化を引き起こし、アトピー性皮膚炎症状の増幅に寄与する可能性が示唆されています。総合的に見ると、分析された知見は、腸内細菌叢がアトピー性皮膚炎の病態生理において重要でありながら十分に解明されていない要素であり、その調節が将来の治療戦略の新たな方向性を示す可能性があることを示唆しています。腸と皮膚の相互作用のメカニズムを解明することは、腸内細菌叢の調節を標的とした予防的アプローチの開発に役立つだけでなく、酸化還元バランスの不均衡が重要な役割を果たす全身性疾患としてのアトピー性皮膚炎のより広範な見方を支持することにもつながる可能性がある。 |
| 目次(クリックして記事にアクセスできます) |
| 1. はじめに |
| 2. アトピー性皮膚炎の病態生理 |
| 2.1. 遺伝的決定因子 |
| 2.2. 皮膚バリア機能障害と皮膚内細菌叢異常 |
| 2.3. 環境要因 |
| 2.4. アトピー性皮膚炎の病態生理における生活習慣因子 |
| 2.5. 免疫応答の調節異常 |
| 2.6. アトピー性皮膚炎の発症に関わるシグナル伝達経路と分子メカニズム |
| 3. アトピー性皮膚炎における腸内細菌叢異常 |
| 3.1. ヒト腸内細菌叢の主要構成 |
| 3.2. アトピー性皮膚炎の発症における短鎖脂肪酸と腸内細菌叢異常の役割 |
| 3.3. 乳児の腸内細菌叢と短鎖脂肪酸産生菌 |
| 3.4. 同一属に属する異なる細菌株の関連性の違い |
| 3.5. 免疫応答調節因子と腸内細菌叢の潜在的な関係 |
| 4. アトピー性皮膚炎における腸内細菌叢と酸化ストレス |
| 4.1. 腸内細菌叢、ディスバイオシス、および免疫恒常性 |
| 4.2. 微生物代謝産物と酸化還元調節 |
| 4.3. 双方向相互作用と臨床的証拠 |
| 4.4. 腸管-皮膚軸と酸化ストレス |
| 4.5. 皮膚微生物叢が消化管恒常性に及ぼす影響 |
| 5. 治療的展望 |
| 5.1. プロバイオティクス |
| 5.2. プレバイオティクス |
| 5.3. シンバイオティクス |
| 5.4. ポストバイオティクス |
| 5.5. 糞便微生物叢移植(FMT) |
| 5.6. バイオマーカーと標的治療戦略 |
| 6. アトピー性皮膚炎における抗酸化物質と抗酸化物質サプリメントの役割 |
| 6.1. ビタミンD |
| 6.2. ビタミンE |
| 6.3. ビタミンC |
| 6.4. カロテノイド |
| 7. 結論 |
| 本文 |
| 1.はじめに |
| 腸内細菌叢は、消化管に生息する細菌、古細菌、ウイルス、真菌からなる複雑かつ動的な微生物群集であり、全身の恒常性維持に重要な役割を果たしています。生理的条件下では、腸内細菌叢は、(i)栄養素の消化と代謝[1]、(ii)短鎖脂肪酸(SCFA)の合成[2]、(iii)免疫応答の調節[3]、(iv)認知機能に影響を与える神経伝達物質の産生[4,5]、(v)腸管バリア機能の強化[6,7]に寄与しています。これらの多面的な機能により、腸内細菌叢は局所免疫だけでなく、腸管と臓器間の相互作用を通じて全身免疫の回復力にも影響を与えます[8,9]。したがって、腸内細菌叢の量的・質的なバランスを維持することは、ヒトの健康にとって不可欠です。 |
| 腸内細菌叢の構成と機能の平衡が崩れることをディスバイオシスと呼びます。その発症に寄与する重要な要因には、(i) 食物繊維、ポリフェノール、短鎖脂肪酸が不足した不健康な食事などの内因性要因 [10]、(ii) 腸内細菌叢の構成と多様性に長期的な変化をもたらす過剰または不適切な抗生物質療法 [11,12]、(iii) 慢性ストレスなどの心理社会的要因 [13,14]、(iv) 大気汚染、重金属、農薬、食品添加物などの環境要因 [15,16,17] などがあります。これらの要因の蓄積は、腸内で慢性的な酸化ストレスを引き起こし、酸化還元バランスの崩壊を通じて、腸上皮の損傷や炎症シグナル伝達経路の活性化につながる可能性があります [18,19]。これらのプロセスは、腸粘膜バリアの弱化、腸管透過性の増加(「リーキーガット」)、および細菌細胞とその代謝産物(毒素やリポ多糖など)の全身循環への移行に直接関連しています[20,21]。その結果、宿主の一次および二次免疫応答が刺激され[22]、NF-κB、MAPK、NLR3などの炎症経路が活性化され、炎症性サイトカインやケモカインの産生につながります[23,24,25]。腸-皮膚軸のこの調節異常は、文献で報告されている研究の増加によって証明されているように、アトピー性皮膚炎の開始と進行につながる可能性があります。 |
| アトピー性皮膚炎患者では、腸内細菌叢の多様性の低下、ビフィドバクテリウムやラクトバチルスなどの有益な属の減少、および大腸菌やクロストリジウムなどの潜在的に病原性のある種の増加がしばしばみられます[26,27,28]。これらの変化は、疾患の重症度および炎症マーカーの上昇と直接相関しています。さらに、研究により、ディスバイオシスはTh1/Th2バランスを崩し、アトピー性疾患に特徴的なTh2プロファイルへの偏りを促進する可能性があることが示されています[29,30,31]。間違いなく、酸化ストレスは、活性酸素種(ROS)の産生と宿主の抗酸化防御機構との間の不均衡を通じて、この病理学的カスケードをさらに悪化させます[32,33]。この状況は、脂質過酸化、タンパク質およびDNA損傷、NF-κBなどの転写因子の活性化を促進し、それによって炎症性サイトカインの産生を増幅します[34,35]。アトピー性皮膚炎患者を対象とした臨床研究では、抗酸化能の低下と同時に酸化ストレスマーカーのレベルが上昇し、皮膚病変の強度と直接相関することが示されています[33,36,37,38]。これらの知見を総合すると、腸内細菌叢が重要な役割を果たす腸-皮膚軸の調節異常に酸化ストレスと腸内細菌叢の異常が関与しているという説得力のある証拠が得られます。腸内細菌叢は代謝変化に寄与し、それによって皮膚バリアの完全性、樹状細胞の活性化、T細胞の分極、および炎症性メディエーターの産生に影響を与えます。この複雑な多段階メカニズムは、アトピー性皮膚炎症状の臨床的重症度、頻繁な再発、および喘息やアレルギー性鼻炎などの他のアトピー性疾患との相関につながります[39,40,41]。 |
| しかしながら、アトピー性皮膚炎の発症機序の解明につながる広範な研究が進められてきたにもかかわらず、腸内細菌叢異常、酸化ストレス、および炎症性メディエーターを統一的なメカニズムの枠組みの中で包括的に臨床的に統合するには、依然として大きなギャップが存在する。本レビューは、腸内細菌叢の乱れがアトピー性皮膚炎の発症と進行に及ぼす影響に関する現在の知見をまとめることを目的としている。本レビューの新規性は、アトピー性皮膚炎における腸内細菌叢異常と慢性炎症を結びつける重要なメカニズム的リンクとして、酸化ストレスの役割に特に焦点を当てている点にある。 |
| 2. アトピー性皮膚炎の病態生理 |
| アトピー性皮膚炎は複雑な疾患であり、最も頻繁にみられる慢性炎症性皮膚疾患の一つです。その高い罹患率にもかかわらず、アトピー性皮膚炎の病態生理は依然として完全には解明されていません。発症要因としては、遺伝的要因、環境要因、皮膚バリア機能障害、免疫応答の調節異常などが重要な役割を果たしていると考えられています[42,43,44]。本稿では、明瞭性と構成を容易にするために各項目を区分していますが、ここで議論する要因は相互に強く関連しており、その影響は重複し、相互作用することで、最終的にアトピー性皮膚炎の発症に寄与していることを強調しておくことが重要です。 |
| 2.1. 遺伝的決定因子 |
| アトピー性皮膚炎の発症リスクを高める遺伝的異常の中でも、フィラグリン(訳者注:フィラグリン(Filaggrin)は、皮膚の表皮角層でバリア機能と保湿の中心的役割を担うタンパク質です。ケラチン線維を束ねて角質細胞を頑丈にすると同時に、分解されて天然保湿因子(NMF)となり肌の潤いを保ちます。不足するとドライスキンやアトピー性皮膚炎を引き起こす要因となります)機能を阻害する変異が最も注目されています[45,46,47,48]。フィラグリン(FLG)は、ヒスチジンを豊富に含む構造タンパク質であり、健康な皮膚では細胞の完全性を維持しています[49]。フィラグリンの前駆体はプロフィラグリンであり、顆粒層(主に)および有棘層の細胞質内にケラトヒアリン顆粒(KHG)を形成します。これらの顆粒は、ケラチンフィラメントとの相互作用および架橋促進を介して、表皮の角化に不可欠な役割を果たします。このプロセスにより、表皮バリアの凝集性が確保され、人体内部環境と外部環境との分離が可能になります[50]。 フィラグリンの変異は、アトピー性皮膚炎の発症と慢性化の両方に大きく寄与することが広く認識されている[49,51]。これらのフィラグリン変異は、表皮バリア機能の障害を伴うことが多く、アレルギー因子への曝露を促進し、皮膚が細菌に定着しやすくなる[52,53]。アトピー性皮膚炎患者のほとんどが皮膚バリア機能の障害を示すが、機能喪失をもたらすフィラグリンヌル変異を持つ患者は3分の1未満である。よく特徴づけられている変異には、R501Xと2282del4がある[54]。重要なことに、フィラグリン変異を持たないアトピー性皮膚炎患者でもフィラグリンレベルの低下が観察されており、このタンパク質が疾患の病因の根底にある基本的な因子であることを示している[48,55]。角質層(SC)中のフィラグリンは、遊離アミノ酸であるピロリドンカルボン酸(PCA)とウロカニン酸(UCA)に分解されることを強調しておく価値がある。これらは、適切な皮膚の水分保持、光防御、および免疫応答の調節に極めて重要である。これらの酸は、天然保湿因子(NMF)の基本要素である[56,57]。アトピー性皮膚炎の重症度、フィラグリン遺伝子型、およびフィラグリン分解産物のレベルの関係を評価した研究では、天然保湿因子の減少が中等度から重度の疾患の主要な構成要素であることが示された[57]。さらに、フィラグリンヌル変異のキャリアは、角質層の全層にわたって天然保湿因子レベルが著しく低下し、経表皮水分損失(TEWL)の増加とアトピー性皮膚炎の特徴である皮膚の乾燥が見られる[58,59]。 |
| 2.2. 皮膚バリア機能障害と皮膚内細菌叢異常 |
| ヒトの皮膚は、人体最大の臓器であり、非常に豊富で多様な微生物群集を宿しているとよく言われます[60,61,62]。定着する微生物の大部分は宿主にとって無害です。さらに、共生的な定着は数多くの利点をもたらします。皮膚微生物叢は、皮膚の代謝調節に関与し、有害な微生物の増殖を防ぐ主要な防御バリアとして機能する可能性があります[62,63,64]。皮膚に常在する微生物には、マラセチア属の真菌や、コアグラーゼ陰性ブドウ球菌(すなわち、表皮ブドウ球菌、ブドウ球菌ルグドゥネンシス、溶血性ブドウ球菌)などの細菌、さらに連鎖球菌科、クチバクテリア科、コリネバクテリア科のメンバーが含まれる[65]。しかし、特定の条件下、特にアトピー性皮膚炎の場合、皮膚のディスバイオシスが発生する可能性がある。定義によれば、皮膚のディスバイオシスとは、この臓器に存在するマイクロバイオームのバランスが崩れることである[63,66]。これにより、常在する微生物が欠如または不足し、病原性微生物が存在し、それらが集合的に宿主に悪影響を及ぼす[67]。健康な皮膚と比較して病変部の皮膚のマイクロバイオームの多様性が低下することは、アトピー性皮膚炎の特徴である[68]。数多くの研究により、アトピー性皮膚炎における黄色ブドウ球菌のコロニー形成の増加は、皮膚常在菌の減少につながり、病原性微生物の増殖を阻害するために皮膚で通常産生される抗菌ペプチド (AMP) の合成の低下をもたらすことが示されている [69,70,71]。したがって、抗菌ペプチド産生の障害は、ウイルス、細菌、真菌による皮膚感染症のリスクを高め、結果として患者の生活の質の低下につながる可能性がある [71]。表 1 は、アトピー性皮膚炎 と診断された個人の皮膚に生息する微生物集団で起こる変化の比較をまとめたものである。 |
| 表1.アトピー性皮膚炎に関連する皮膚の細菌叢異常 |
 |
| 皮膚バリアは、皮膚の微生物叢、環境要因、遺伝的決定因子、および宿主免疫システム間の複雑な相互作用の橋渡しとして機能します[49]。このバリアの完全性は、角質化または角質化として知られるプロセスを経る表皮によって維持されます。組織学的には、表皮は、体の部位によって異なる4つまたは5つの層で構成されています[50]。最も外側の層は角質層です。内側に向かって、表皮は、通常足の裏と手のひらにのみ存在する透明層、続いて顆粒層、有棘層、および基底層で構成されています[50,52]。バリア機能不全はアトピー性皮膚炎患者の大多数に影響し、この疾患の特徴的な特徴です[54,73]。バリアの機能は主に角質層によって決定され、そこで角化の段階が起こります。角質細胞は核を取り除かれ、扁平化して角質細胞になり、その細胞膜は角質化した外膜に置き換えられます[74,75]。表皮バリアが破壊されると、刺激に対する感受性が高まり、外来抗原が皮膚に侵入しやすくなり、そこで常在免疫細胞と相互作用して炎症を引き起こします[59,74,76]。逆に、皮膚の炎症が長引くと表皮バリアがさらに損なわれるため、アレルギー疾患の治療と予防の両方においてバリアの完全性を維持することの重要性が強調されます[74]。 |
| 2.3. 環境要因 |
| 皮膚は毒素や紫外線などの物質から身を守る防御器官としての役割を担っているため、個人の行動を含む外部環境要因がアトピー性皮膚炎の経過にどのように影響するかを考察することは興味深い[77]。環境要因の中でも、防腐剤、刺激の強い洗剤、香料などの化学物質は、アトピー性皮膚炎の病態生理に寄与することが知られている。強アルカリ性の洗浄剤の使用は、皮膚のpHバランスを崩し、局所的な炎症を促進する可能性がある[42]。水でさえ、表皮のフィラグリンレベルを低下させ、機能的な皮膚バリアの構造を破壊する可能性がある[55]。皮膚バリアが弱体化すると、環境要因の影響を受けやすくなり、アトピー性皮膚炎では炎症が悪化し、角質層への二次的な損傷を引き起こし、病気の悪循環を永続させる[73]。上記要因に加え、環境汚染もアトピー性皮膚炎の発症に影響を与えると考えられている[76]。居住地の大気汚染への曝露は、男性のみにおけるアトピー性皮膚炎の発症リスクの上昇、および男女両方における空気中のアレルゲンに対するアレルギー反応の可能性の上昇と関連している[78]。さらに、二酸化硫黄、二酸化窒素、一酸化炭素などのガス、および空気中に存在する粒子状物質は、アトピー性皮膚炎の独立した原因物質として分類されている[79]。さらに、粒子状物質への曝露は、実験的にTNF-α(腫瘍壊死因子アルファ)を介してケラチノサイトにおけるフィラグリン発現を抑制し、経表皮水分損失の増加をもたらした[80]。収集されたデータは、化学物質から大気まで、広義の環境要因が皮膚バリアの完全性を著しく損ない、それによってアトピー性皮膚炎の発症と悪化に寄与する可能性があることを明確に示している。 |
| 2.4. アトピー性皮膚炎の病態生理における生活習慣因子 |
| 喫煙はアトピー性皮膚炎の罹患率を高める要因の一つと考えられています[81]。喫煙と複数のアレルギー疾患、特に喘息との間には遺伝的相関関係も認められています[82]。喫煙への曝露と喘息やアトピー性皮膚炎などの小児アレルギー疾患の発症との関連性を調査するため、合計53,505人の小児の対照データを含む研究が行われました[83]。注目すべきことに、妊娠中の母親の喫煙と受動喫煙の両方が小児喘息と関連していましたが、タバコへの接触と小児アトピー性皮膚炎の発症との間には統計的に有意な関連性は認められませんでした[83]。しかし、2024年のより新しいメタアナリシスでは、子どものアトピー性皮膚炎は妊娠中の母親の喫煙とは関連していませんが、受動喫煙とは相関関係があることが明らかになっています[84]。能動喫煙と受動喫煙とアトピー性皮膚炎の発症との関係を評価した別の研究では、より年齢の高い集団、すなわち韓国の青年を対象としています。 アトピー性皮膚炎の有病率と各タイプの喫煙との接触頻度および強度の両方に関して正の相関関係が観察されました [85]。幼少期のタバコの煙への曝露は、成人期のアトピー性皮膚炎発症につながる可能性があります [86]。興味深いことに、2018 年の二次横断分析では、喫煙の種類と参加者の性別の両方が成人の アトピー性皮膚炎の有病率に影響を与えることが示されました。女性では、アトピー性皮膚炎の発生は現在のタバコの使用と関連していることがわかりましたが、男性では、能動喫煙または受動喫煙とアトピー性皮膚炎の発生との間に相関関係は観察されませんでした [87]。 |
| 睡眠障害は、アトピー性皮膚炎患者によく見られる合併症であり、小児人口の約47%から80%、成人人口の33%から90%に影響を及ぼしています[88,89]。興味深いことに、睡眠の質の低下は、成人患者、男性、および重症アトピー性皮膚炎患者で特に顕著であり、一般的に、睡眠開始よりも夜間を通して回復睡眠を維持することがより困難であることと関連しています[90]。小児患者では、掻痒が睡眠障害の原因として報告される主な症状です。アトピー性皮膚炎の小児における睡眠障害の結果には、行動障害、低身長、認知機能障害、および気分の変動が含まれる可能性があります[91]。アトピー性皮膚炎に存在する炎症状態は、炎症性サイトカインが皮膚局所だけでなく全身で上昇するため、追加の精神障害の発症にも寄与する可能性があります。さらに、睡眠は免疫系の機能の中心的な調節因子として広く認識されています。さらに、睡眠障害は心理的ストレスの増加を伴うことが多い[92]。ストレスは、アトピー性皮膚炎を含むさまざまな疾患の経過を悪化させるよく知られた要因である[93]。ストレスはかゆみの閾値を低下させ、夜間の掻痒を引き起こし、すでに損なわれている皮膚バリアをさらに悪化させる可能性がある[92]。ストレス自体も、炎症性サイトカインの分泌を刺激し、創傷治癒を遅らせ、アトピー性皮膚炎、ニキビ、乾癬、蕁麻疹などの皮膚疾患を悪化させることにより、皮膚バリア機能を損なう可能性がある[93]。心理的ストレスがアトピー性皮膚炎の発症と悪化に重要な役割を果たしていることを示唆する証拠が増えているが、その根底にある病態生理学的メカニズムは完全には解明されていない[94]。 |
| 2.5. 免疫応答の調節異常 |
| 免疫系の機能障害は、アトピー性皮膚炎の病態生理において重要な要素です。主な複合的な異常は、Th1/Th2、Th17、およびTreg細胞の応答の調節異常です。アトピー性皮膚炎の急性期では、免疫応答は主にTh2細胞によって誘導されますが、慢性期ではTh1、Th17、およびTh22細胞の応答が優勢になります[53,95,96]。表皮のランゲルハンス細胞(LC)は主要な免疫細胞として機能し、真皮のより深部にはマクロファージ、T細胞、および樹状細胞(DC)が存在します。皮膚のケラチノサイトは、TNFやインターロイキンIL-1β、IL-6、IL-10、IL-18などの炎症促進性サイトカインを分泌する能力を持っています[69]。ケラチノサイトは、Thリンパ球の活性を調節する炎症促進性サイトカインである胸腺間質リンパ球増殖因子(TSLP)も合成することができます[53]。胸腺間質リンパ球増殖因子は、樹状細胞を誘導し、Th2型免疫応答を刺激して炎症を増幅し、アレルギー反応性を高めるため、主要な感作分子です[95,97]。機能不全の皮膚バリアは、ケラチノサイトによる胸腺間質リンパ球増殖因子、IL-1β、IL-25、およびIL-33の分泌を促進します[98]。これにより、ランゲルハンス細胞と樹状細胞が刺激され、Th2、Th17、およびTh22免疫応答の活性化が引き起こされます。Th1応答は、ランゲルハンス細胞によって以前に活性化された肥満細胞(MC)を介して誘導されます。Th1リンパ球はその後、IL-1、IL-6、IL-8、IL-10、TNF-α、およびIFN-γを分泌します。対照的に、Th2リンパ球はIL-4、IL-5、IL-13、IL-31、およびIL-33を放出します[98,99]。Th2駆動の炎症は、サイトカインの存在下で成熟するケラチノサイトに影響を与え、フィラグリン発現を低下させ、それによって既存の皮膚バリア機能障害を悪化させる可能性があることを強調する価値があります[54,73]。さらに、Th2細胞によって産生されるサイトカインは、Bリンパ球を刺激してIgE抗体を産生させ、それが今度は肥満細胞脱顆粒とヒスタミン、ロイコトリエン、プロスタグランジンなどのメディエーターの放出を誘導します。これが掻痒-掻きむしりサイクルを促進し、皮膚バリアをさらに損傷します[43,45,53,99]。図1は、アトピー性皮膚炎の主要な病態生理学的メカニズムの概略図を示しています。 |
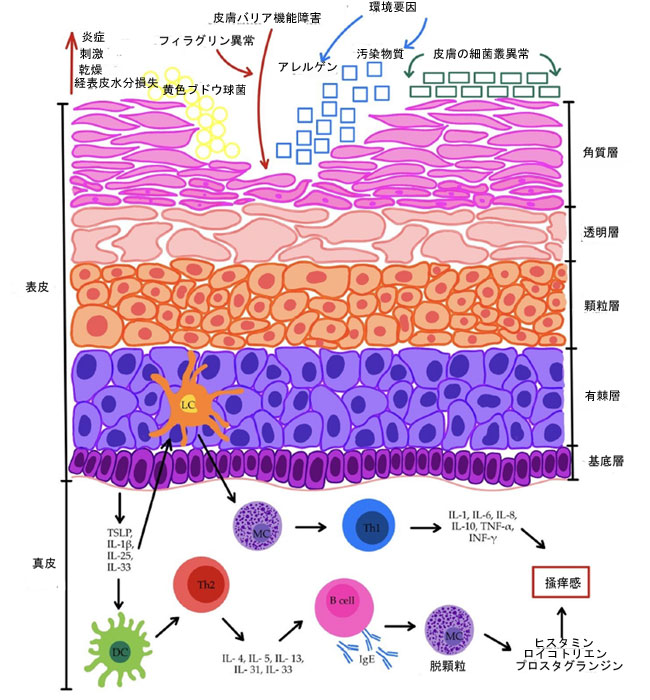 |
図1. アトピー性皮膚炎の多因子病因。 略語: B 細胞 - B リンパ球。 DC - 樹状細胞。 FLG - フィラグリン。 IFN-γ - インターフェロン ガンマ。 IgE - 免疫グロブリン E; IL-1 - インターロイキン 1; IL-1β - インターロイキン 1 ベータ。 IL-10 - インターロイキン 10; IL-13 - インターロイキン 13; IL-25 - インターロイキン 25; IL-31 - インターロイキン 31; IL-33 - インターロイキン 33; IL-4 - インターロイキン 4; IL-5 - インターロイキン 5; IL-6 - インターロイキン 6; IL-8 - インターロイキン 8; LC - ランゲルハンス細胞。 MC - マスト細胞。S. aureus- 黄色ブドウ球菌。 TEWL—経表皮水分損失;Th1—Tヘルパー1リンパ球;Th2—Tヘルパー2リンパ球;TNF-α—腫瘍壊死因子アルファ;TSLP—胸腺間質リンパ球増殖因子。 記号:黄色の円は黄色ブドウ球菌の定着を示します。青色の矢印と青色の四角は、アレルゲンや汚染物質などの環境要因が皮膚に浸透していることを示します。緑色の長方形は、皮膚微生物叢の構成の乱れ(皮膚ディスバイオシス)を示します。赤色の矢印は、皮膚バリア機能の障害が皮膚に及ぼす影響を示します。 |
| 2.6. アトピー性皮膚炎の発症に関わるシグナル伝達経路と分子メカニズム |
| アトピー性皮膚炎の病因は、複雑な分子メカニズムによって引き起こされます。その一つに、サイトカインの主要な細胞内シグナル伝達カスケードを構成するヤヌスキナーゼ-シグナル伝達兼転写活性化因子(JAK-STAT)シグナル伝達経路の活性化があります。JAKファミリーにはJAK1、JAK2、JAK3、チロシンキナーゼ2(TYK2)の4つのメンバーが同定されており、STATファミリーにはSTAT1、STAT2、STAT3、STAT4、STAT5A、STAT5B、STAT6の7つのメンバーが同定されています[100]。重要なことに、JAK-STAT経路はTh2細胞を介した免疫応答、特にアトピー性皮膚炎の急性期に関与するサイトカインであるIL-4、IL-13、IL-31によって引き起こされる免疫応答において極めて重要な役割を果たします[101]。 IL-4 は、フィラグリンなどの重要なタンパク質の発現を低下させ、B 細胞における IgE クラススイッチングを促進し、好酸球や肥満細胞などの免疫エフェクター細胞を動員することにより、皮膚バリアの完全性を損ないます [102,103]。IL-4 は、2 種類の受容体を介してシグナル伝達します。1 つは IL-4Rα 鎖と共通 γ 鎖で構成される I 型受容体、もう 1 つは IL-4Rα 鎖と IL-13 受容体 α1 (IL-13Rα1) 鎖で構成される II 型受容体です [100]。II 型 IL-4 受容体複合体は、主にケラチノサイト、線維芽細胞、免疫細胞などの非造血細胞に発現し、アトピー性皮膚炎の発症に大きく寄与します。一方、I 型 IL-4 受容体複合体は主に造血細胞に限定されます。さらに、II型IL-4受容体はIL-13受容体としても機能し、IL-13は免疫経路と神経経路を結びつける掻痒の重要なメディエーターである[102]。I型IL-4受容体を介した経路では、JAK1とJAK3がリン酸化によって活性化され、その後STAT6シグナル伝達につながる。逆に、II型IL-4受容体複合体の刺激はJAK1とTYK2の活性化をもたらし、JAK1はIL-4Rα鎖に、TYK2はIL-13Rα1鎖に結合している。これらの事象に続いてSTAT6とSTAT3が誘導され、最終的に2型炎症を促進する遺伝子の発現が上方制御される[100,102]。 IL-31受容体は、IL-31受容体α鎖(IL-31RA)とオンコスタチンM受容体β鎖(OSMRβ)からなるヘテロ二量体を含む2つの形態でも存在し得る[100]。DCの活性化を介して、IL-4、IL-13、およびIL-31は、CCL17、CCL22、およびCCL26などのケモカインの産生を促進し、それによってCCR4+ Th2細胞を炎症部位に動員する[102]。胸腺間質リンパ球増殖因子は、2型免疫応答の開始において重要なサイトカインでもある。アトピー性皮膚炎では、皮膚上皮細胞で高発現しており、掻痒によって誘発される皮膚バリアの破壊時にIL-33とともにケラチノサイトからさらに放出される[101]。胸腺間質リンパ球増殖因子は、一過性受容体電位(TRP)感覚ニューロンを活性化し、さらなる掻痒に寄与する。さらに、TSLPはケラチノサイトを刺激してペリオスチンとIL-33を放出させ、それがIL-13とIL-31の産生を増強し、かゆみと掻きむしりのサイクルを維持する[102,103]。さらに、胸腺間質リンパ球増殖因子は樹状細胞上のOX40L(訳者注:OX40L(OX40リガンド、TNFSF4とも呼ばれます)は、樹状細胞などの抗原提示細胞上に発現するタンパク質です。T細胞の表面にある受容体「OX40」と結合することで、T細胞の活性化や生存、増殖を強力に促す「共刺激分子」として機能します)の発現を促進し、ナイーブT細胞をTh2表現型に分極させるための重要なシグナルとなる[102]。 |
3. アトピー性皮膚炎における腸内細菌叢異常 |
| 3.1. ヒト腸内細菌叢の主要構成 |
| ヒトの腸内には、細菌、古細菌、真菌、ウイルスなど100兆個以上の微生物が生息し、複雑で多様な生態系を形成している[28,104]。腸内細菌の総重量は約1~1.5kgである[28]。成人の腸内に生息する細菌の大部分は、グラム陰性菌であるバクテロイデス門とグラム陽性菌であるフィルミクテス門の2つの細菌門である[28,105]。細菌の99%以上は、フィルミクテス門、バクテロイデス門、プロテオバクテリア門、およびアクチノバクテリア門で構成されている[28]。ヒトの腸内には、その他の細菌門も存在し、その割合は個人によって大きく異なる。これらには放線菌門、フソバクテリア門、ベルコミクロビア門が含まれる[105]。例えば、ベルコミクロビア門は3~12歳の子供に多く見られる[106]。ビフィドバクテリウムはヒトの腸に最初に定着する細菌の一つであり、授乳期には腸内細菌叢の優勢なメンバーであり、通常は少なくとも最初の3~4ヶ月間は優勢である[107]。 |
| 3.2. アトピー性皮膚炎の発症における短鎖脂肪酸と腸内細菌叢異常の役割 |
| 生後1ヶ月目から、腸内細菌叢はヒトの免疫系の成熟に不可欠です。これは、翻訳後修飾ペプチド、アミノ酸代謝物、短鎖脂肪酸、オリゴ糖、糖脂質、非リボソームペプチドなど、全身の免疫応答に影響を与える様々な代謝物やシグナル分子の産生と密接に関連しています[105]。健康な腸内細菌叢は、Th1細胞の発達を促進することでTh1/Th2バランスを維持します。腸の健康に影響を与える主な要因としては、微生物の多様性、短鎖脂肪酸産生菌の豊富さ、そして潜在的に病原性のある菌の存在などが挙げられます[106]。細菌種の多様性の低下は、腸内細菌叢異常と関連しており、炎症の亢進にさらに寄与する可能性があります。 アトピー性皮膚炎の発症と重症度は、腸内細菌叢の異常の程度と相関すると考えられています。さらに、アトピー性皮膚炎患者では短鎖脂肪酸のレベルが低下していることが報告されています [105,108]。酪酸、プロピオン酸、酢酸、乳酸などのこれらの酸は、腸内細菌による食物繊維の発酵によって生成されます。これらは、腸上皮バリアの完全性を維持する上で重要な役割を果たします。短鎖脂肪酸レベルが低いと、毒素、未消化の食物粒子、腸内微生物が全身循環に容易に侵入し、皮膚の炎症を悪化させます [105]。短鎖脂肪酸はまた、自然免疫を強化し、炎症に対する皮膚の防御を改善し、獲得免疫をサポートします。これは、T 細胞と B 細胞の分化に影響を与えることによって起こります [109]。特に、Th1、Th17、および制御性 T 細胞 (Treg) の産生を調節することによって起こります [106,110]。 短鎖脂肪酸の中でも、酪酸は粘膜Treg分化に不可欠である[110]。Treg細胞の調節異常やサイトカインの不均衡、特にIL-6やIL-17などの炎症性サイトカインの増加は、炎症性皮膚疾患の発症と進行に寄与する可能性がある[106,110]。短鎖脂肪酸、腸管バリア、および全身性免疫調節の相互作用を図2(簡略図)にまとめた。 |
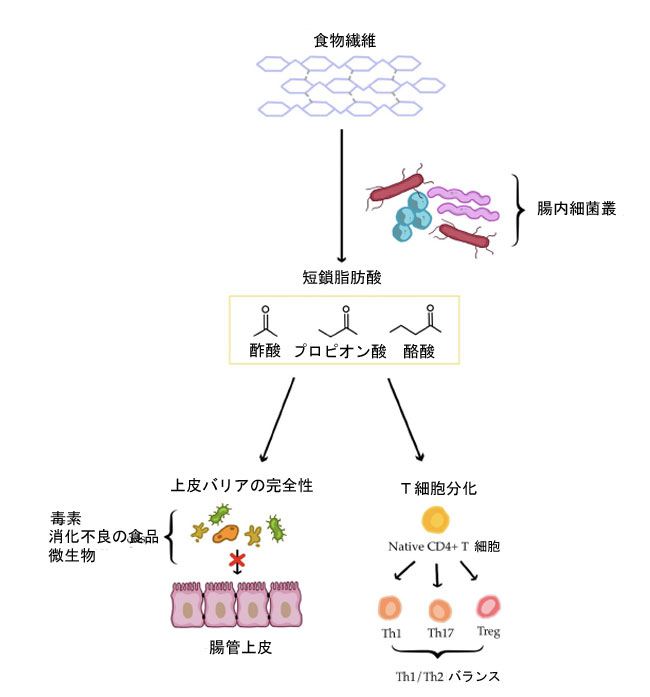 |
図2. ヒトの腸管および免疫系における短鎖脂肪酸の役割(簡略図) 略語:Th1—T1ヘルパーリンパ球;Th17—T17ヘルパーリンパ球;Treg—制御性Tリンパ球。 |
| 3.3. 乳児の腸内細菌叢と短鎖脂肪酸産生菌 |
| アトピー性皮膚炎の発症率は乳児期(生後12ヶ月)に最も高く、幼児期まで続きます[111]。乳児の腸内細菌叢の多様性の低さとアトピー性皮膚炎発症予測との間に正の相関関係があることは、様々な研究で実証されています[108,112,113]。アトピー性皮膚炎の乳児は、健康な対照群と比較して、腸内細菌叢の多様性が著しく低く、クロストリジウム・ディフィシル、大腸菌、バクテロイデス属菌のレベルが高いことが示されています[114,115]。クロストリジウム属菌は、好酸球性炎症を介して炎症レベルを悪化させる可能性があります[115]。クロストリジウム科の増加は、毒素の過剰放出と関連しており、これは好中球の走化性を阻害し、好酸球の活性を抑制し、それによって腸の炎症を促進することが、24のコホートにわたるアトピー性皮膚炎リスクを調べた研究で示されています[116]。別の研究[117]では、アトピー性皮膚炎や病原性大腸菌などの種が、乳児の腸上皮バリアの漏出増加と関連していました。「リーキーガット症候群」は、短鎖脂肪酸産生不足にも関係しています[105]。不十分な産生は、病原性種による短鎖脂肪酸産生細菌の抑制による可能性があります。注目すべきことに、短鎖脂肪酸を産生するクロストリジウム目の一部、例えばクロストリジウム目ファミリーXIII Incertae Sedisなどは、アトピーセイ皮膚炎患者で減少しています[106]。Alamらによる研究[108]では、アトピーを発症するリスクが最も高い就学前児童は、乳児期に測定されたビフィドバクテリウム、アッケルマンシア、フェカリバクテリウムなどの特定の細菌属の初期レベルが低いことが示されました。これは、アッケルマンシア、ラクトバチルス、フェカリバクテリウム・プラウスニッツィ、ビフィドバクテリウムのレベルの低下がアトピー性皮膚炎患者に特異的であったモニアガらの研究結果とほぼ一致しています[105]。重要なことに、アッケルマンシア、ビフィドバクテリウム、フェカリバクテリウムは短鎖脂肪酸の産生菌です[116]。アトピー患者におけるユーバクテリア科の減少は、酪酸産生の障害と強く関連しています。さらに、ユーバクテリア科の存在量が多いと、Th2 応答に関与する炎症促進性メディエーターであるエオタキシン/CCL11 (好酸球走化性タンパク質/C-C モチーフケモカイン 11)、MDC/CCL22 (マクロファージ由来ケモカイン/C-C モチーフケモカイン 22)、および Flt3L (fms 関連チロシンキナーゼ 3 リガンド) のレベルが低下することが関連付けられています [106]。 |
| 3.4. 同一属に属する異なる細菌株の関連性の違い |
| ビフィズス菌属のレベル低下は、母乳育児の欠如によって影響を受ける可能性がある[118,119,120]。ビフィズス菌属は、マウスモデルにおいて短鎖脂肪酸を産生することにより、炎症性皮膚疾患に対する保護因子として一般的に考えられている[110]。さらに、腸内で乳酸菌とビフィズス菌によって産生されるγ-アミノ酪酸は、皮膚のかゆみを抑制することが示されている[105]。しかし、ビフィズス菌カテヌラタムやビフィズス菌シュードカテヌラタムなどの特定のビフィズス菌種の発生率が高いと、さまざまな年齢層でアトピー性皮膚炎の発症に影響を与えることが報告されている[108,121]。別の研究では、Depnerらは、糞便サンプルの 16S rRNA アンプリコン シーケンスを使用しました [107]。生後最初の 2 か月では、ビフィドバクテリウム属のメンバーが優勢であり、アトピーセイ皮膚炎と最も強く一貫した逆相関を示したことがわかりました。母乳中のヒトミルクオリゴ糖 (HMO) を消化し、有益な芳香族乳酸を生成する特定の能力を持つ ビフィズス菌ロンガムやビフィズス菌ビフィダムなどの種は、生後 2 か月で最も多く見られました。2〜12 か月のアトピー性皮膚炎患者では、ビフィドバクテリウム属の構成が大きく変化し、特に ビフィズス菌アドレセンティスとB.カテヌラタムなどの成人のような ビフィドバクテリウム属への早期の移行を示しました。この現象は、アトピーセイ皮膚炎に対するビフィドバクテリウムの保護効果は、重要な期間と適切な種の組み合わせに依存することを示唆しています [107]。 |
| 先に引用した研究では、アトピーセイ皮膚炎患者におけるF. プラウスニッツィの存在量の減少が強調されているが[105,108]、このグループではその数が増加している可能性も示唆されている[120,122]。F. プラウスニッツィは、健康な人では短鎖脂肪酸の主要な産生菌である。しかし、短鎖脂肪酸産生が不十分なF. プラウスニッツィの亜種は、アトピー性皮膚炎患者でより一般的である[123]。これが研究結果の潜在的な違いを説明できる可能性がある。さらに、短鎖脂肪酸耐性デンプンを分解する炭水化物活性酵素(CAZymes)をコードする遺伝子は、アアトピー性皮膚炎患者の腸内細菌叢では十分に存在していない[124]。 |
| 3.5. 免疫応答調節因子と腸内細菌叢の潜在的な関係 |
| ベイロネラ科、フシカテニバクター、フラボニフラクター、オドリバクターなどの他の細菌は、メンデルランダム化研究で示されているように、循環炎症性サイトカインへの影響を介して、炎症性皮膚疾患に対する潜在的な保護因子として同定されている。サイトカインIL-15RAは、細菌科ベイロネラ科と湿疹を結びつける重要なメディエーターとして同定されている。IL-15RAはMAPKシグナル伝達経路を活性化し、IL-6、IL-8、TNF-αなどの炎症性サイトカインの分泌を促進する。フシカテニバクターとオドリバクターは、IL-10産生とTreg分化を調節することで免疫バランスの維持に役立つ。中等度から重度のアトピー性皮膚炎ではフラボニフラクターの存在量の増加が観察されており、免疫調節異常におけるその潜在的な役割が示唆されている[109]。Kalashnikovaらが実施した別の研究では、小児の血清中の微生物群集の多様性とサイトカインレベルとの間の潜在的な関連性が分析された[106]。アトピー群では、Th2応答の特徴であるいくつかのマーカーのレベルが有意に増加していることが観察された。これらには、MDC/CCL22、IL-5、IL-8、IL-13、IFN-γ(インターフェロンγ)、TNF-α、MIP-1α/CCL3(マクロファージ炎症性タンパク質1-α/ケモカイン(C-Cモチーフ)リガンド3)、およびVEGF(血管内皮増殖因子)が含まれる。一方、アトピー患者ではIL-2、IL-1α、IL-15、およびIL-17Aのレベルが低下していた[106]。 IL-18R1と湿疹のリスク増加の間にも因果関係がある。IL-18RはTh1、ナチュラルキラー細胞、肥満細胞など様々な免疫細胞に発現しており、広範な免疫調節機能を示唆している[109]。別の研究[123]では、SCORAD(アトピー性皮膚炎のスコアリング)指数を用いて評価したアトピー性皮膚炎の重症度が高い小児では、糞便中のカルプロテクチン濃度が高く、血中好酸球数とIgE濃度も上昇しており、消化管の炎症状態が亢進していることが示された。さらに、腸内微生物の影響を受ける血流中のIL-10とIFN-γの濃度は、ストレスホルモンであるコルチゾールのレベルを変化させる。その結果、コルチゾール濃度の上昇は腸内細菌叢の構成と腸管バリア透過性に悪影響を及ぼし、悪循環を生み出す可能性がある[105]。 MDC/CCL22はアトピー性疾患の重要なメディエーターです。パスツレラ科とMDC/CCL22レベルの間には正の相関関係が見られました。一方、MDC/CCL22レベルとバーネシエラ科、オシロスピラ科、ペプトコッカス科、ユーバクテリウム・コプロスタノリゲネス、クロストリジウム UCG-014などの細菌分類群の間には負の相関関係が見られ、これらの種が保護効果を持つ可能性が示唆されました。注目すべきことに、同じ研究では、アトピー患者ではパスツレラ科、ペプトコッカス科、クロストリジウム属 UCG-014の存在量が有意に増加したのに対し、バルネシエラ科、ユウバクテリア科、クロストリジウム目ファミリー XIII Incertae Sedis(訳者注:Incertae sedisは、分類学において「所属が不確実」であることを意味します)、オシロスピラ科、アナエロボラ科、フラボバクテリア科などの他のグループでは対照群と比較して減少が見られました[106]。 24のコホートのデータセットの分析によると、ラクノスピラ科UCG001株は腸内細菌叢の異常を悪化させ、さらにアミノ酸代謝障害、血中尿酸値の上昇、CD4 TH17誘導性炎症と相関関係にあることが示されています[116]。特定の細菌株のさまざまな影響は表2と表3にまとめられており、アトピー性皮膚炎の発症に対して保護的または有害な影響を与える可能性のある微生物が示されています。 |
表2.アトピー性皮膚炎の発症に対して保護効果を持つ可能性のある細菌 |
| 略語: eotaxin/CCL11—好酸球走化性タンパク質/C-Cモチーフケモカイン11; Flt3L—fms関連チロシンキナーゼ3リガンド; HMO—ヒトミルクオリゴ糖; IL-10—インターロイキン10; IL-15RA—インターロイキン15受容体αサブユニット; MDC/CCL22—マクロファージ由来ケモカイン/C-Cモチーフケモカイン22; SCFAs—短鎖脂肪酸; Treg—制御性Tリンパ球。 |
| 表3.アトピー性皮膚炎の発症に悪影響を及ぼす可能性のある細菌 |
 |
| 略語: CD4 TH17 - T ヘルパー 17 リンパ球。大腸菌 - 大腸菌; MDC/CCL22 - マクロファージ由来ケモカイン/C-C モチーフ ケモカイン 22 |
| レビューされた研究結果は、腸内細菌叢と免疫系の関係が非常に複雑であり、湿疹の診断と治療の両方において複数の潜在的な標的が存在することを示している。 |
| 4. アトピー性皮膚炎における腸内細菌叢と酸化ストレス |
| 4.1. 腸内細菌叢、ディスバイオシス、および免疫恒常性 |
| 腸内細菌叢は、免疫寛容の形成、酸化バランスの調節、および上皮バリア機能の維持において中心的な役割を果たす、非常に動的で代謝的に活発な生態系を構成しています[125]。その構成の乱れ、一般にディスバイオシスと呼ばれるものは、アトピー性皮膚炎を含む慢性炎症性疾患の発症と持続における主要な要因としてますます認識されています[126]。ディスバイオシスは宿主の免疫平衡を乱すだけでなく、酸化還元恒常性にも大きな影響を与え、それによって酸化ストレスと炎症の自己持続的なループを助長し、アトピー性皮膚炎における皮膚病変の慢性化と悪化に寄与することが、多くの証拠によって示されています。腸内細菌叢異常の最も重大な結果の1つは、抗炎症作用と抗酸化作用を持つ有益な共生菌、特にF. プラウスニッツィと様々なビフィドバクテリウム属菌の喪失と、腸内細菌科やクロストリジウム・パーフリンゲンスなどの病原菌の増加である[96,122,127]。この構成の変化は、微生物代謝の深刻な障害につながる。 |
| 4.2. 微生物代謝産物と酸化還元調節 |
| 健康な状態では、常在菌は食物繊維を発酵させて短鎖脂肪酸、特に酪酸、プロピオン酸、酢酸を生成します。酪酸は、大腸上皮細胞のエネルギー源として機能し、腸管バリア機能を強化し、強力な免疫調節作用と抗酸化作用を発揮するため、特に重要です[128]。メカニズムとしては、酪酸はNrf2シグナル伝達経路(訳者注:Nrf2シグナル伝達経路は、細胞の酸化ストレス応答と解毒・抗炎症作用を司る最も重要な生体防御システムの一つです。この経路は、老化防止や疾患予防の観点から非常に注目されています)を活性化し、スーパーオキシドジスムターゼ、カタラーゼ、グルタチオンペルオキシダーゼなどの抗酸化応答因子の転写を促進します[129,130]。さらに、短鎖脂肪酸はヒストン脱アセチル化酵素阻害を介してエピジェネティック制御を調節し、制御性T細胞(Treg)の分化と安定性を促進します。 短鎖脂肪酸の欠乏は、制御性T細胞機能の障害、炎症促進性のTh2およびTh17応答への偏り、および活性酸素種生成と組織損傷を直接刺激するIL-4、IL-5、IL-13、IL-17、IL-22などのサイトカインの過剰放出をもたらします[131,132]。トリプトファン代謝は、ディスバイオシスと酸化ストレスの間のもう1つの重要なリンクを構成します[133]。共生細菌は、トリプトファンを免疫調節代謝物(インドール誘導体やキヌレニン経路中間体など)に変換する主要な貢献者であり、これらはアリール炭化水素受容体(AhR)を活性化します。アリール炭化水素受容体シグナル伝達は、上皮バリアの完全性と酸化還元恒常性の維持の中心です。腸内細菌叢のバランスが崩れた状態では、トリプトファンの異化を行う微生物の能力が低下するため、これらの保護代謝産物のレベルが低下し、酸化ストレスに対する感受性が高まり、粘膜の免疫寛容が失われる。 |
| 4.3. 双方向相互作用と臨床的証拠 |
| 酸化ストレス自体がディスバイオシスを悪化させ、双方向的な病原性関係を確立する。活性酸素種レベルの上昇は微生物のDNAとタンパク質を損傷し、変異原性副産物を生成し、ストレス耐性表現型が強化された細菌を選択する[134]。活性酸素種はまた、腸管内腔の酸化還元電位を変化させ、酪酸産生菌などの偏性嫌気性菌を抑制し、酸化ストレス下で増殖する通性嫌気性菌や日和見病原菌にとって好ましいニッチを作り出す[135]。上皮レベルでは、活性酸素種はミトコンドリア機能を障害し、脂質過酸化を増加させ、上皮タイトジャンクションと粘膜防御を弱める。バリア機能の喪失は、さらなる微生物の移行と抗原への曝露を許容し、免疫活性化を持続させ、活性酸素種産生を加速させ、炎症とディスバイオシスのサイクルを永続させる[136]。臨床的および実験的証拠は、この双方向の関係を支持しています。8-ヒドロキシ-2′-デオキシグアノシン(8-OHdG)、高度酸化タンパク質産物、マロンジアルデヒド(MDA)などの酸化ストレスのバイオマーカーは、アトピー性皮膚炎の疾患の重症度と強い相関関係を示します[36]。アトピー性皮膚炎コホートにおける並行したマイクロバイオータプロファイリングは、一貫して短鎖脂肪酸産生分類群の減少と炎症関連細菌の過剰発現を示しています。マウスモデルでは、抗生物質誘発性ディスバイオシスが短鎖脂肪酸を枯渇させることによりアトピー性皮膚炎様皮膚炎症を悪化させることが示されており、腸内代謝と皮膚炎症を結びつける微生物の不均衡の因果的役割が確認されています[36]。 |
| 4.4. 腸管-皮膚軸と酸化ストレス |
| 腸管由来の酸化および炎症シグナルが皮膚組織に影響を与える重要な病態生理学的メカニズムの一つは、腸管透過性の亢進、一般に「リーキーガット」と呼ばれる現象である[36]。活性酸素種誘導性の上皮損傷は、炎症性サイトカインの活性によってさらに悪化し、クローディン、オクルディン、ゾヌラオクルデンス-1などのタイトジャンクションタンパク質を破壊する。その結果、リポ多糖(LPS)、ペプチドグリカン、フラジェリン、その他の代謝産物などの微生物成分が全身循環に移行する[99]。これらの病原体関連分子パターン(PAMP)は、自然免疫細胞上のトール様受容体(TLR)およびヌクレオチド結合オリゴマー化ドメイン(NOD)様受容体を活性化し、全身性炎症を増幅させる。循環する炎症性メディエーターは、ケラチノサイト、ランゲルハンス細胞、真皮樹状細胞などの皮膚免疫細胞に作用し、炎症プロセスを腸から皮膚へと拡大します[96]。この腸と皮膚のクロストークは、アトピー性皮膚炎における炎症の全身性伝播のメカニズム的基盤を確立します。治療の観点から、これらの知見は、ユーバイオシスを回復し、酸化ストレスを軽減することを目的とした介入の可能性を強調しています。プロバイオティクスとプレバイオティクスの補給は、短鎖脂肪酸レベルを増加させ、上皮バリア機能を強化し、活性酸素種蓄積を抑制することが示されています[137]。ラクトバチルス・ラムノサスGGやB. ロンガムなどの菌株は、免疫応答を調節し、アトピー性皮膚炎の重症度を軽減する上で特に有効であることが実証されています。さらに、発酵性食物繊維とポリフェノールが豊富な食事介入は、短鎖脂肪酸の産生をサポートし、抗酸化防御を強化します。 N-アセチルシステイン、ビタミンE、ポリフェノール化合物(例:レスベラトロール、ケルセチン)の使用など、酸化ストレスを標的とする並行戦略は、酸化バイオマーカーの減少と臨床症状の改善に有効であることが示されています[36]。重要なことに、腸内細菌叢の調節と抗酸化物質の補給を組み合わせたアプローチは、ディスバイオシスと酸化ストレスの悪循環を断ち切る上で優れている可能性があります。アトピー性皮膚炎における腸内ディスバイオシス、酸化ストレス、皮膚炎症を結びつける提案されたメカニズムは、図3にまとめられています。 |
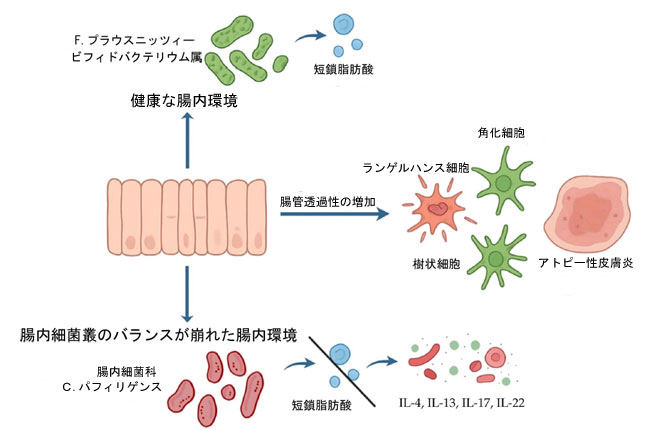 |
図3. アトピー性皮膚炎における腸-皮膚軸の提案。 健康な腸では、F.プラウスニッツィーやビフィドバクテリウム属などの共生細菌が、腸管バリアの完全性と免疫恒常性をサポートする短鎖脂肪酸を産生します。腸内細菌科と C. パフィリゲンス(訳者注:Clostridium perfringens(ウェルシュ菌)は、土壌や腸管に常在する偏性嫌気性グラム陽性桿菌です。主に食中毒(下痢・腹痛)やガス壊疽の原因菌として知られ、100℃で1時間耐える芽胞を形成し、大鍋調理などの大量調理現場で増殖しやすい特徴を持ちます)の過剰増殖を特徴とするディスバイオシスは、短鎖脂肪酸産生を減少させ、腸管透過性の増加 (「リーキーガット」) に寄与します。これにより、免疫調節異常が促進され、皮膚の樹状細胞とランゲルハンス細胞の活性化、炎症性サイトカイン (IL-4、IL-13、IL-17、IL-22) レベルの上昇、および アトピー性皮膚炎の発症につながります。 略語: IL-13—インターロイキン 13; IL-17—インターロイキン 17; IL-22—インターロイキン 22; IL-4—インターロイキン 4; SCFA;短鎖脂肪酸。 |
| 4.5. 皮膚微生物叢が消化管恒常性に及ぼす影響 |
| 腸-皮膚軸は複雑な双方向コミュニケーションネットワークであり、皮膚バリアの破壊や環境刺激に対する皮膚の反応は、全身免疫と腸の健康に大きな影響を与える[125]。従来の研究では腸が皮膚に及ぼす影響に焦点が当てられてきたが、最近のマウスを用いた実験モデルでは、皮膚損傷や皮膚ヒアルロン酸の消化が腸内微生物叢を直接変化させ、腸管免疫恒常性を阻害することが示されている[138]。具体的には、マウスの皮膚損傷やヒアルロニダーゼの活性化により、ヒアルロン酸(HA)断片が全身循環し、損傷関連分子パターン(DAMP)として機能する。これらの皮膚由来シグナルは、結腸におけるReg3やMuc2などの宿主防御遺伝子の発現増加を引き起こす。特に、ヒト結腸上皮細胞およびマウス結腸組織を用いた研究では、ヒアルロン酸断片がReg3の発現を直接誘導することが確認されている。さらに、マウスの皮膚の炎症は全身性炎症反応を引き起こし、腸の炎症を悪化させるIgEやTNF-αなどのメディエーターを上昇させる[138,139]。この「逆」軸シグナル伝達により、マウスの腸内の生菌総数が測定可能なほど減少する一方で、バクテロイデス・テタイオタオミクロンのような毒性の強い日和見病原菌が選択される。生き残ったこれらの微生物は、マウスの粘液層や結腸上皮を貫通する能力が高まり、宿主の重症大腸炎に対する感受性が増加する。 |
| 5. 治療的展望 |
| 腸内細菌叢を標的とした治療戦略は、有益な腸内細菌叢を促進することで酸化ストレスを軽減できる。最も一般的なアプローチとしては、プロバイオティクス、プレバイオティクス、シンバイオティクス、ポストバイオティクス、そして糞便微生物移植(FMT)などが挙げられる。同時に、バランスの取れた食事と適切なサプリメントの選択の重要性も強調されている[140]。 |
| 5.1. プロバイオティクス |
| 国連食糧農業機関(FAO)と世界保健機関(WHO)によると、プロバイオティクスは、最適な量を摂取することで宿主に健康上の利益をもたらす生きた微生物と定義されています[141,142]。最も一般的に使用されているプロバイオティクス種はグラム陽性菌であり、ラクトバチルス属(L.ラムノサス GG、L.スポロジェン、L.ロイテリ RC-14、L.プランタルム299v、L.アシドフィルス、L.ラクティス)、ビフィドバクテリウム属(B. ビフィダム、B.ロンガム、B.インファンティス)、ストレプトコッカス属(S.サーモフィルス、S.ラクティス)、エンテロコッカス属などが含まれます。これらの細菌は、健康な人の腸内常在菌叢の一部を構成しています[143,144]。 |
| さらに、ラクトバチルスやビフィドバクテリウムなどのプロバイオティクスは、スーパーオキシドジスムターゼ(SOD)やカタラーゼ(CAT)などの酵素を発現し、グルタチオン(GSH)レベルを上昇させ、酪酸などの抗酸化特性を持つ代謝産物を生成することで、活性酸素種の生成を抑制し、抗酸化活性を示します[148,149,150]。L. カゼイとL.ファーメンタムを補給した後、スーパーオキシドジスムターゼとカタラーゼが有意に増加することが観察されました[149,150]。数多くの研究が行われているにもかかわらず、プロバイオティクスの抗酸化作用の正確なメカニズムはまだ完全には解明されていません。 |
| 29件の全文記事のメタ分析(うち15件は統合メタ分析に含まれた)では、プロバイオティクスが酸化ストレスマーカーに及ぼす影響が評価された。この研究では、低用量(0.4 × 10¹⁰ CFU未満)のプロバイオティクス補給により、血清マロンジアルデヒド濃度が有意に低下することが示された。さらに、プロバイオティクス補給により、グルタチオン、総抗酸化能(TAC)、および一酸化窒素(NO)のレベルが増加した。注目すべきは、プロバイオティクス補給の期間が酸化ストレスの調節に大きく影響することである。投与期間が10週間未満の研究では、初期段階の酸化ストレスマーカーであるマロンジアルデヒドレベルの低下が観察された。この低下は、初期段階におけるスーパーオキシドジスムターゼの活性化に起因する可能性がある。対照的に、長期にわたるサプリメント摂取期間(10週間以上)は、グルタチオンレベルを増加させることで抗酸化メカニズムの次の段階を促進し、グルタチオンレベルはスーパーオキシドジスムターゼによって生成された過酸化水素(H2O2)の中和に関与する[151]。研究結果では、従来のアトピー性皮膚炎治療とプロバイオティクスの同時投与により、外因性アトピー性皮膚炎およびCC(C-159T)遺伝子型の成人における標準治療の有効性が有意に向上することが報告されている[152]。 |
| 小児アトピー性皮膚炎の高い罹患率と影響を考慮すると、あるメタアナリシスでは、出産前後に母親と乳児にL.ラムノサスを投与することが、アトピー性皮膚炎の予防に効果的な戦略となる可能性があることが示唆されている。しかし、最適な投与量、介入期間、および作用機序を決定するには、さらなる研究が必要である[153]。同様に、別のレビューでは、単一成分および複数成分のプロバイオティクスを補給することでアトピー性皮膚炎のリスクが減少することが示されている[154]。 |
| 5.2. プレバイオティクス |
| プレバイオティクスは、腸内における有益な細菌株の増殖や活性を特異的に刺激する難消化性食品成分であり、宿主の健康に好影響をもたらします[141,155]。プレバイオティクスには、食物繊維(オリゴ糖)、特にフラクトオリゴ糖、ガラクトオリゴ糖(GOS)、イヌリンなどが含まれます[156]。これらの物質は、ビフィズス菌や乳酸菌の増殖を促進し、主に腸内細菌叢の構成に影響を与えることが示されています[157]。プレバイオティクスは、宿主の消化管内で腸内細菌叢の作用により発酵され、短鎖脂肪酸、すなわち酢酸、プロピオン酸、酪酸が生成されます。これらの短鎖脂肪酸は抗炎症作用を発揮し、毒性発酵産物の形成を減少させ、グルタチオン産生のためのニコチンアミドアデニンジヌクレオチドリン酸(NADPH)合成を刺激し、それによって活性酸素種生成を減少させる[150,156]。さらに、プレバイオティクスはTh1/Th2比と腸管関連リンパ組織におけるリンパ球および/または白血球の数を増加させる。また、IgA(免疫グロブリンA)分泌を促進し、IL-18(抗炎症性サイトカイン)を上方制御する一方で、IL-6(炎症性バイオマーカー)および酸化ストレスに関与する遺伝子を下方制御することが示されている[156]。 |
| β-グルカンやイヌリンなどのプレバイオティクスがアトピー性皮膚炎の発症と進行に及ぼす影響を調査する研究が行われた。この目的のために、研究者らはオキサゾロン(OX)誘発アトピー性皮膚炎様マウスモデルを用いた。プレバイオティクスは、感作期間終了後2週間(治療研究)と感作開始前3週間(予防研究)に経口投与された。この研究の結果、プレバイオティクス補給により皮膚病変の重症度と炎症反応が有意に軽減されることが示された。さらに、プレバイオティクスを投与されたマウスの真皮では、表皮の厚さと免疫細胞浸潤が有意に減少した。特に、β-グルカンとイヌリンの早期投与は、アトピー性皮膚炎マウスの腸内善玉菌の増殖を刺激し、それによってアトピー性皮膚炎の進行を阻止した[132]。 |
| 5.3. シンバイオティクス |
| プロバイオティクスとプレバイオティクスの相乗効果を利用したシンバイオティクスは、製品処方において高い有効性を示すことが実証されている[158]。シンバイオティクスは主に、プレバイオティクスによってプロバイオティクス微生物を胃酸から保護することで、プロバイオティクス微生物の生存率と機能性を向上させることを目的としている[141]。 |
| ラクトバチルス・カゼイとイヌリンからなるシンバイオティクスを投与すると、マロンジアルデヒド、過酸化水素、グルタチオン、遊離スルフィドリル基の存在などの酸化ストレスマーカーを効果的に調節できる可能性がある[155]。さらに、メタアナリシスでは、1歳以上の小児のアトピー性皮膚炎の管理に対するシンバイオティクスの好ましい影響が確認されている[159]。 |
| 5.4. ポストバイオティクス |
| プロバイオティクス、プレバイオティクス、シンバイオティクスに加え、ポストバイオティクスは、プロバイオティクス微生物が発酵過程を経て生成する生物活性代謝物または副産物と定義されます[160]。ポストバイオティクスは、ヨーグルト、ザワークラウト、漬物、コンブチャなどの食品に天然に存在するほか、ラクトバチルス属、ビフィドバクテリウム属、ストレプトコッカス属、ユーバクテリウム属、フェカリバクテリウム属、サッカロミセス属など、様々な細菌や真菌によっても産生されます[161]。ポストバイオティクスは、その独特な構造特性により保存安定性を提供し、炎症や酸化ストレスの調節に関わる多様な生物学的経路を活性化することができます[162]。ポストバイオティクスは、複製能力のない非生存微生物代謝物であるため、免疫機能が低下している人にとってプロバイオティクスよりも安全な代替手段となることを強調すべきである[163]。 |
| ラクチプランチバチルス・プランタルム由来のポストバイオティクスは、皮膚の炎症に関与する細菌に対して抗菌作用を示します。また、TNF-α/IFN-γで刺激されたケラチノサイトにおいて、炎症性サイトカイン(IL-6、IL-8、TSLP)および皮膚バリアに関連する遺伝子(INV、FLG、LOR)を調節し、皮膚バリアを強化します[164]。あるメタアナリシスでは、ラクトバチルス属ポストバイオティクスの経口投与がアトピー性皮膚炎の重症度を軽減する可能性があることが示されました[165]。有望な結果にもかかわらず、アトピー性皮膚炎の治療におけるポストバイオティクスの臨床使用に関する決定的な証拠は、現時点では不十分です。 |
| 5.5. 糞便微生物叢移植(FMT) |
| 糞便微生物移植は、アトピー性皮膚炎に対する有望な治療戦略です。これは、健康なドナーから採取した糞便サンプルから得られた腸内微生物叢を、治療目的でレシピエントの消化管に移植するものです[166]。糞便微生物移植は酪酸レベルの上昇を促進し、リポ多糖濃度の低下、酸化ストレスの軽減、炎症反応の抑制、腸管バリア機能の維持につながります[167]。さらに、糞便微生物移植は酸化ストレスマーカーの発現を正常化することで、これらのマーカーにも影響を与えます[168]。 |
| アトピー性皮膚炎マウスにおける腸内細菌叢の回復は、疾患症状を軽減するための戦略として研究された。糞便微生物移植後、マウスはTh1/Th2バランスが再確立され、IgEレベルが低下し、肥満細胞、好酸球、好塩基球の数が減少した。免疫応答を調節することにより、糞便微生物移植はヒトのアトピー性皮膚炎治療のための新しいアプローチとなる可能性があるが、さらなる研究が必要である[169,170]。表4は、アトピー性皮膚炎に対する微生物叢を標的とした治療戦略の概要を示している。 |
| 表4.アトピー性皮膚炎治療のための腸内細菌叢を標的とした治療戦略 |
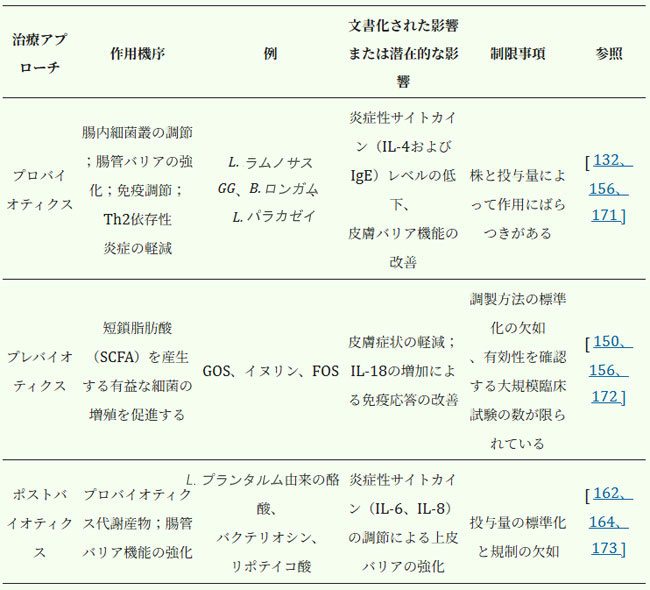 |
 |
| 略語: FMT - 糞便微生物叢移植。 FOS - フラクトオリゴ糖。 GOS - ガラクトオリゴ糖;。 |
| 5.6. バイオマーカーと標的治療戦略 |
| アトピー性皮膚炎の病態生理学的メカニズムの理解が進んだことで、疾患特異的なバイオマーカーが同定され、抗IL-31モノクローナル抗体(ネモリズマブ)、抗OX40L剤(IMG-007)、抗TSLP療法(APG777)、選択的JAK1阻害剤など、より効果的で分子標的的な治療戦略の開発への道が開かれました[176,177]。 |
| ネモリズマブは、インターロイキン-31受容体のαサブユニットのアンタゴニストとして作用するヒト化モノクローナル抗体であり、IL-31シグナル伝達経路を阻害します。IL-31は皮膚の受容体に結合すると、炎症細胞の動員を促進し、表皮バリアタンパク質の発現を減少させ、感覚神経を活性化し、これらが複合的に掻痒に寄与します[178,179]。したがって、IL-31はアトピー性皮膚炎における重要な治療標的となります。ネモリズマブの投与により、第III相試験で16週間後に患者の42~44%で皮膚病変が75%以上減少することが示されています。さらに、ネモリズマブと局所療法を併用すると、掻痒が有意に改善しました[177]。 |
| APG777は、皮下投与によりIL-13を標的とするヒト化モノクローナル抗体です。アトピー性皮膚炎における主要なバイオマーカーを大幅かつ持続的に阻害します。予備データでは、現在の治療法と比較して高い有効性と投与頻度の低減(3~6か月ごと)の可能性が示唆されています。フェーズベースの臨床試験が現在進行中です[180]。IMG-007は、アトピー性皮膚炎および円形脱毛症の治療薬として研究中の革新的な長時間作用型抗OX40モノクローナル抗体(抗OX40 mAb)です。この抗体はT細胞を排除することなく炎症経路を阻害し、良好な安全性プロファイルを維持しながら投与頻度の低減(12週間ごと)を可能にする可能性があります[180]。これら2つの薬剤は、投与間隔の延長と作用持続時間の延長により医療システムへの負担が軽減され、患者の治療遵守率が向上する可能性が高いため、治療および経済面で大きな可能性を秘めています[180]。 |
| JAK阻害剤(JAKi)は、アトピー性皮膚炎を含む免疫応答の調節異常を標的とする新しいクラスの治療薬です。JAK阻害剤は細胞内JAK-STATシグナル伝達経路(訳者注:JAK-STAT(ジャック-スタット)シグナル伝達経路は、細胞外のサイトカインなどのシグナルを細胞核へ伝え、遺伝子の発現を引き起こす情報伝達系です。免疫応答、細胞の増殖・分化・生存などを調節する極めて重要な経路で、その異常は自己免疫疾患やがんを引き起こします)を阻害し、疾患の発症に関与する複数の炎症性サイトカインの活性化を阻止することが示されています[181]。JAK阻害剤は高い有効性を示す一方で、比較的高い副作用のリスクも伴うことに注意することが重要です[178]。局所ルキソリチニブの有効性と安全性が評価され、炎症の軽減とより迅速な抗掻痒効果が得られました[182]。デュピルマブと併用して100mgまたは200mgの経口アブロシチニブを投与した場合の臨床的有用性は、全身治療を必要とする中等度から重度のアトピー性皮膚炎患者で評価されています。両薬剤とも皮膚病変の軽減において同等の活性を示しましたが、200mgのアブロシチニブ投与では、わずか2週間の治療で掻痒感が有意に軽減されました[183]。 2 mgまたは4 mgの用量で投与されたバルチシチニブによる介入は、中等度から重度のアトピー性皮膚炎患者において、掻痒の改善とともに持続的な長期治療効果をもたらすことが示されている[184]。JAK阻害剤やモノクローナル抗体などの新しい標的療法は、中等度から重度のアトピー性皮膚炎患者を対象とした臨床試験で有効性を示している。しかし、その正確な作用機序は完全には解明されていない。これらの分子メカニズムを解明し、長期的な安全性を評価するためには、さらなる臨床研究および実験研究が必要である。 |
| 6. アトピー性皮膚炎における抗酸化物質と抗酸化物質サプリメントの役割 |
| 本稿では、表皮バリアを損傷し慢性炎症を持続させることでアトピー性皮膚炎の発症機序において中心的な役割を果たす酸化ストレスのメカニズムについて概説する。したがって、活性酸素種を中和し、酸化と抗酸化のバランスを回復させる主要な機能を持つ抗酸化物質を検討することは妥当である[185,186]。アトピー性皮膚炎おいて最も広く研究されている抗酸化物質には、ビタミンD(VD)、ビタミンE(VE)、ビタミンC(VC)、カロテノイド、メラトニンなどがある[187]。メタアナリシスによると、抗酸化療法はアトピー性皮膚炎の重症度を大幅に軽減する可能性があるが、かゆみの緩和には顕著な効果は認められない。この研究には、アトピー性皮膚炎患者763名を対象とした18件のランダム化臨床試験が含まれている。最も大きな治療効果は、ビタミンDの経口投与、ビタミンDとビタミンEの併用投与、ビタミンA(VA)、ビタミンD、ビタミンEの併用投与、およびビタミンB12の局所投与で認められた。総じて、抗酸化物質はアトピー性皮膚炎患者に対する安全で潜在的に効果的な補助療法となる可能性がある[188]。 |
| 6.1. ビタミンD |
| ビタミンDは、日光からの紫外線(UV)照射によって皮膚で内因的に産生される脂溶性ステロイド化合物群です[189]。ビタミンDは、ケラチノサイトの分化過程、およびカテリシジンやヒトβ-ディフェンシンなどの抗菌ペプチドの産生に大きな影響を与えることが明らかになっています。これらのペプチドは、フィラグリン合成を促進し、炎症性サイトカインの産生を抑制することで、皮膚バリア機能の強化に寄与することが示されています。さらに、ビタミンDは皮膚感染症の予防にも重要な役割を果たします[187,190]。ビタミンDはTh2/Th17経路を調節し、Treg活性を促進することで、アトピー性皮膚炎の症状を改善する可能性を秘めています[190]。 39 人の アトピー性皮膚炎患児と 20 人の健康な非アレルギー患児を対象に、1000 IU/日 (25 μg/日) の経口ビタミンD補給による 3 か月間の アトピー性皮膚炎の重症度の変化を評価する研究が行われた。補給後、SCORAD 指数と IL-2、IL-4、IL-6、IFN-γ などの炎症性サイトカインのレベルの両方で減少が観察された [191]。別の研究では、SCORAD の減少に加えて、患部の皮膚における ビタミンD 受容体とカテリシジンの発現も増加したと報告されている。カテリシジンは抗菌特性を持つ AMP ファミリーに属するタンパク質である [192]。686 人の アトピー性皮膚炎患者を対象とした 11 件のランダム化比較試験のメタ分析により、ビタミンD 補給が小児と成人の両方でアトピー性皮膚炎の強度を軽減することが確認された [193]。さらに、血清ビタミンD濃度が低いアトピー性皮膚炎患者では、より重篤な経過をたどることが観察された[194]。既存の文献を包括的に分析した結果、ビタミンD補給の肯定的な効果とアトピー性皮膚炎症状の緩和については、より長期間の治療を含む大規模な研究によって確認する必要があると結論付けられる。 |
| 6.2. ビタミンE |
| ビタミンE(トコフェロールとトコトリエノール)は脂溶性の強力な生物学的抗酸化物質であり、酸素フリーラジカルを中和し、ケラチノサイトの分化を促進することで、細胞を酸化ストレスから保護します[36]。メタアナリシスでは、1日400 IUのビタミンE投与により、アトピー性皮膚炎患者のかゆみが改善する可能性が示唆されています[195]。ある研究では、アトピー性皮膚炎の小児におけるビタミンEの高用量摂取と血清中の総IgE/特異的IgEレベルとの関係を評価することを目的としました。この研究では、ビタミンEの摂取量が比較的多いほど血清中の総IgEレベルが低下し、アトピー性皮膚炎の症状が改善される可能性があると示唆されています[196]。 |
| 6.3. ビタミンC |
| ビタミンC(L-アスコルビン酸とも呼ばれる)は、細胞を酸化ストレスから保護し、コラーゲン生合成、カルニチンおよびカテコールアミン代謝に関与し、免疫機能をサポートし、食事からの鉄吸収を促進する必須の水溶性抗酸化物質です。ビタミンCは、果物や野菜の摂取を通じて食事からのみ得られます[197]。ビタミンCは、皮膚からの水分損失を防ぐことで表皮の完全性を維持するのに役立ちます[198]。アトピー性皮膚炎患者20名を対象とした非盲検試験では、ポリ-L-リジンとビタミンAおよびビタミンCを併用した製剤を28日間投与しました。この試験では、SCORADスケールで疾患強度が41.8%低下し、生活の質が改善したほか、紅斑、掻痒、乾燥も減少しました[199]。別の研究では、ビタミンCとビタミンEを豊富に含む母親の食事が新生児のアトピーの発症に及ぼす影響と、その潜在的な予防効果を評価することを目的とした。母乳中のビタミンC濃度が高いほど、乳児のアトピーのリスクが低下することが報告されている[200]。しかし、この研究は小規模なグループで行われたため、ビタミンCがアトピー性皮膚炎の症状の重症度を軽減すると断言することは難しい。その有効性を確認するには、さらなる研究が必要である。 |
| 6.4. カロテノイド |
| カロテノイド(β-カロテン、α-カロテン、γ-カロテン、β-クリプトキサンチンなど)は、植物、藻類、細菌によって合成される天然の疎水性色素であり、植物に黄色、オレンジ色、赤色を与えます。これらは抗酸化物質として働き、光合成装置を光酸化から保護するとともに、ビタミンAの前駆体としても機能します。カロテノイドは、免疫系、細胞間コミュニケーション、胚発生、造血、アポトーシスの調節に関与しています。これらの色素は、抗酸化作用、抗炎症作用、抗血管新生作用、抗増殖作用も示します[201]。カロテノイドは活性酸素種レベルを低下させ、脂質過酸化を抑制し、スーパーオキシドジスムターゼやグルタチオンペルオキシダーゼなどの内因性抗酸化酵素の活性を高めます。この作用は、特に皮膚バリアの保護に重要です[202,203]。さらに、カロテノイドは、TNF-α、IL-6、IL-1βなどの炎症誘発性サイトカインの発現を減少させ、COX-2(シクロオキシゲナーゼ-2)およびLOX(リジルオキシダーゼ)を阻害し、皮膚に抗炎症作用を発揮することが報告されている[203]。ある研究では、妊娠中に緑黄色野菜、柑橘類、β-カロテンを多く摂取すると、子供の湿疹や喘息の予防に役立つ可能性があることが示された[204]。別の研究では、カロテノイドファミリーに属する黄色色素であるルテインの濃度が高いほど、アトピー性皮膚炎のリスクが低いことが有意に関連していることが示唆された[205]。 |
| 7. 結論 |
| アトピー性皮膚炎は、世界中の小児および成人にみられる一般的な炎症性皮膚疾患です。その罹患率の高さにもかかわらず、アトピー性皮膚炎の病態生理は依然として解明されておらず、新たな治療戦略の探索における重要な焦点となっています。アトピー性皮膚炎発症の主な誘発因子としては、皮膚バリア機能障害、皮膚および腸内細菌叢の異常、遺伝的要因(特にフィラグリン遺伝子変異)、環境要因、免疫応答の調節異常などが挙げられます。本レビューでは、腸内細菌叢の異常と酸化ストレスの相互作用が、アトピー性皮膚炎の中心的な病原軸であることを強調しています。腸内細菌叢の異常は、免疫調節作用および抗酸化作用を持つ微生物由来代謝産物の産生を減少させ、炎症反応の亢進と活性酸素種産生の増加につながります。活性酸素種レベルの上昇は、腸内細菌叢と上皮バリアをさらに損傷し、ディスバイオシスを永続させ、最終的に皮膚に影響を与える微生物および炎症性メディエーターの全身への拡散を招きます。これらの相互に関連するプロセスは、炎症を持続させ、疾患を悪化させる慢性的な自己強化フィードバックループを確立します。さらに、プロバイオティクス、プレバイオティクス、シンバイオティクス、ポストバイオティクス、糞便微生物移植などの微生物を用いた新たな治療アプローチは、微生物バランスの回復と疾患の重症度軽減に有望な展望をもたらします。重要なのは、今後の研究において、アトピー性皮膚炎における腸内細菌叢の構成、酸化ストレス、および免疫調節の間の複雑な相互作用をより正確に解明することを目指すべきであるということです。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
| この文献は、Antioxidants (Basel). 2026 Feb 27;15(3):299.に掲載されたFrom Dysbiosis to Inflammation: Gut Microbiota and Oxidative Stress in Atopic Dermatitis.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |