Roberta Prete,Sarah Louise Long et al., |
要約 |
| 胆汁酸 (BA) の特徴は、多くの病状で変化します。 胆汁酸代謝は、腸内での定着と存続、および胃腸 (GI) 通過中の微生物の生存を助ける重要な微生物の機能であり、潜在的なプロバイオティクス細菌の重要な基準です。 ゲートウェイ胆汁酸修飾酵素である胆汁酸塩加水分解酵素(BSH)を発現する微生物は、腸内で有利であると考えられています。 この特性は、腸関連微生物に選択的に限定されると報告されています。 食品関連微生物は、人間の消費者に健康上の利益をもたらす可能性があります。 今回我々は、食品に関連したLactobacillus plantarum株は胆汁酸代謝が可能であり、胆汁酸に関連したストレスに耐えて増殖することができ、これが胃腸消化管(GIT)生存にとって重要な特徴として認識されているということを報告する。 さらに、我々は、これらの食品関連Lactobacillus plantarum株が、ヒトにとって有益となる受容体活性化に有利に胆汁酸サインを変化させる選択的能力を有することを報告する。 実際、調べた株はすべて、ヒトグリコール結合胆汁酸を変更する明らかな優先性を示しましたが、株に依存した修飾も明らかでした。 この研究は、食品由来の非病原性細菌による胆汁酸代謝が微生物と人間の両方にとって有益であることを実証し、食物関連の非病原性分離株の中で、これまで腸内居住者に特有であると考えられていた進化的に保存された特徴を特定するものである。 |
| 目次(クリックして記事にアクセスできます) |
| 1.はじめに |
| 2.結果 |
| 2.1.胆汁ストレス時の胃腸消化管環境における細菌生存の可能性 |
| 2.2.細菌による胆汁酸プロファイルの変化:胆汁酸塩加水分解酵素脱抱合活性 |
| 3.議論 |
| 4.方法 |
| 4.1.細菌株 |
| 4.2.胆汁酸塩耐性 |
| 4.3.胆汁耐性 |
| 4.4胆汁酸塩加水分解酵素脱抱合活性 |
| 4.5.統計分析 |
| 5.補足情報 |
| 本文 |
| 1.はじめに |
| 食品関連微生物は人間の宿主と相互作用し、腸の健康に潜在的な利益をもたらすと考えられています(1,2)。 しかし、これらの利点が食事の成分のみに関連しているのか、宿主微生物叢に対する食品の直接的な影響や、それらが餌を与えている微生物に関係しているのか、あるいはそれらが運ぶ微生物によって引き起こされる効果と連動しているのかを判断することは困難であることが判明しています。 なぜなら、食品由来の微生物の蔓延を調査した研究はほとんどないからです(1-3)。 消化管通過を生き延びて腸内に存続するためには、常在微生物と一過性微生物が動的な生理学的ストレスに耐える能力を備えていなければなりません。 経口摂取後、細菌は胆汁などの分泌物に関連する多くの人間の防御システムに遭遇します。 胆汁は胆嚢の消化分泌物であり、肝臓でコレステロールから合成され、回腸末端で肝臓に再循環されます(4,5)。 胆汁は脂質の乳化と可溶化に主要な役割を果たしており、胆汁の流れを生成して脂質、内因性代謝産物、生体異物の肝胆汁分泌を促進します(6)。 胆汁の主成分は胆汁酸で、有機成分の 50% 以上を占め、消化界面活性剤として働き、食物脂質と脂溶性ビタミンを提示して乳化し、消化と摂取を促します(7)。 特定の胆汁酸部分は、その洗浄特性により、高レベルで細菌および宿主細胞膜の破壊、タンパク質の変性、鉄およびカルシウムのキレート化などの抗菌性および炎症性の特性を有しており、DNA に酸化的損傷を引き起こす可能性があります7。 したがって、胆汁および胆汁酸に耐える微生物の能力は、微生物の生存と消化管内での存続にとって重要であると認識されています( 8)。 |
| 肝臓では、コレステロールは一次胆汁酸であるケノデオキシコール酸(CDCA)とコール酸(CA)に変換され、その後N-アシルアミデートとして主にアミノ酸のタウリンまたはグリシンに結合します。 そして、小腸で自己集合して脂肪を提示するミセルになります(9)。 腸内細菌叢、特にLactobacilli spp. は胆汁酸を環境シグナルとして、また場合によっては栄養素や電子受容体として利用することができます(10)。 重要なことに、腸内細菌は、主に微生物胆汁酸塩加水分解酵素(BSH)を介して胆汁酸塩代謝に主要な役割を果たしており、アミド結合の加水分解によって胆汁酸塩の脱共役を触媒し、ステロイド核からグリシン/タウリン部分を放出します(9、13、14)。 この機能は、胆汁酸で生体内変換における重要なゲートウェイ反応として認識されており (9,14)、一次遊離胆汁酸コール酸およびケノデオキシコール酸を再生し、コール酸、リトコール酸 (LCA)、およびウルソデオキシコール酸 (UDCA) からの二次胆汁酸デオキシコール酸 (DCA) の微生物形成を促進します。 ケノデオキシコール酸およびさまざまな中間体から得られます (Long et al., 2017 によるレビュー)。 これらの多くは現在、宿主の脂質、コレステロール、グルコース、エネルギー代謝、薬物代謝、免疫応答の調節の調節に関与する強力な細胞シグナル伝達分子として認識されています(15,16)。 これらの受容体には、胆汁酸リガンド活性化核内受容体、胆汁酸恒常性を制御する肝臓と腸のファルネソイド X 受容体 (FXR)、ビタミン D 受容体 (VDR) が含まれます。 これは、エネルギー恒常性 G タンパク質共役胆汁酸受容体 TGR516 と同様に、多くの異なる役割を持っています。 |
| 脱結合によって放出されたアミノ酸は、細菌の栄養と生存のための炭素および窒素源としてさらに利用できます(17)。 胆汁酸塩加水分解酵素はまた、細菌膜へのコレステロールの組み込みを促進し、膜電位、流動性、および引張強度を変化させる可能性があります (18)。 これは、胆汁酸塩耐性と宿主ディフェンシンに対する感受性を強化することにより、腸内細菌の生存率を効果的に改善することが提案されています(4)。 胆汁酸塩加水分解酵素は、乳酸菌の消化管生存率を高める界面活性剤ショックタンパク質である可能性があることが示唆されています(19)。 これを裏付けるように、微生物の生息地と胆汁酸塩加水分解酵素活性の間に強い相関関係があることが報告されています(8,20)。 さらに、腸内胆汁酸塩の脱結合は、胆汁酸プールと腸内細菌叢の両方のサイズと組成に影響を与えるため、腸の恒常性にとって不可欠です(21)。 |
| 胆汁酸塩加水分解酵素脱結合活性は腸内微生物属の間でよく表され(10,22)、主にグラム陽性菌種であるBifidobacterium (23-25)、Clostridium (26,27)、Enterococcus (28,29)、Listeria (30)、および Lb. plantarum種 (39-43) を含む Lactobacillus (31-38)の間で特徴付けられています。 さらに、何人かの研究者は、胆汁耐性は菌株特異的な形質であり、種の耐性は種または属内で一般化することはできないと報告しています(15,16)。 初期の報告では、胆汁酸塩加水分解酵素活性は消化管関連共生生物に割り当てられていましたが、環境分離株には割り当てられていませんでした(8)。 彼らは、bsh コード配列が主要な門のすべてに存在し、胆汁酸塩加水分解酵素活性が環境によって異なることを発見しました。 彼らの研究で調べたところ、すべての腸関連乳酸菌株は胆汁酸のグリコとタウロの両方を脱結合させることができました。 対照的に、発酵乳から単離された乳酸菌は、糖結合胆汁酸を好む傾向を示しますが、タウロ脱結合(特に TDCA)についてはいくつかの重複がいくつかの菌株で検出されました (44)。 |
| 胆汁酸塩加水分解酵素活性による 胆汁酸代謝は、プロバイオティクス菌株を選択するための重要な EFSA (訳者注:欧州食品安全機関)基準です(45)。 これは、菌株が胃腸通過を生き延びる可能性があることを示すものと考えられます。 より最近では、微生物の 胆汁酸塩加水分解酵素活性が微生物と宿主の対話に影響を与え、宿主の脂質代謝を機能的に調節し、コレステロール代謝を変化させ、潜在的に体重減少に影響を与えることが示されました(21)。 |
| 胆汁酸の特徴が体重減少に影響を与える正確なメカニズムは現在不明であるため、胆汁酸塩加水分解酵素に関しては一般化された規則を適用できません。胆汁酸代謝の重要性は、腸関連微生物においてのみ研究されています(4)。 この研究では、著者らによって分離され特徴づけられた食品由来のLb. plantarum と既知のタイプ株が検査され、in vitro で胆汁関連ストレスに耐える能力が異なることが示されました。 さらに、胆汁酸部分を代謝する各菌株の特異的能力を調べた。 胆汁酸誘導体はヒトの代謝性疾患を治療するための治療薬として認識されているため、この研究は、食品関連微生物の特性が宿主にも利益をもたらす可能性があることを示唆しています。 そしてそれは、機能性食品における検討と開発の重要なターゲットとしての食品ベースの微生物の役割を強化します。 |
| 2.結果 |
| 2.1.胆汁ストレス時の胃腸消化管環境における細菌生存の可能性 |
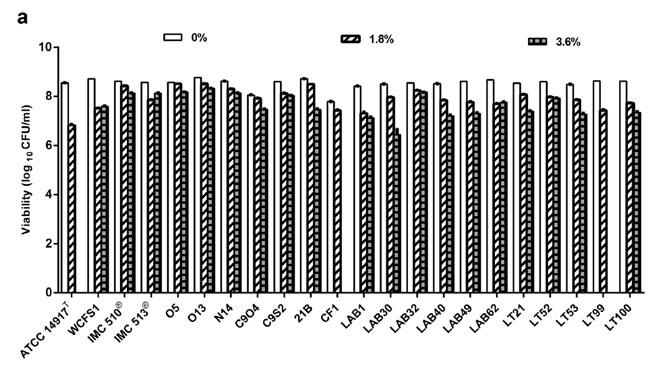
| 胆汁酸塩耐性と耐性胆汁酸塩の存在下で増殖する能力は、胆汁酸塩の濃度を増加させながら (0 ~ 3.6% w/v) 共培養中に 600 nm での光学密度を測定することによってモニタリングされました。 1.8%未満の試験濃度では、大部分の株で微生物の生存率は変化しませんでした(図1aおよびSI図S1a)。 しかし、キャベツのピクルス、サワードウ、生乳チーズからそれぞれ分離されたタイプ株 Lb. plantarum ATCC14917、CF1、および LT99 では、生存率の大幅な低下が記録されました。 この効果は、3.6%ウシ胆汁の存在下でこれらの菌株の方が顕著であり、これらの3菌株はすべて食品由来で生存可能な増殖が記録されません(図1a)。 食品由来の残りの菌株(オリーブ、キャベツのピクルス、サワー種、生乳チーズ)は、ヒト分離株 WCFSI(ヒト唾液由来)、IMC510、および IMC513(ヒト腸分離株)と同程度の胆汁レベルに耐えることができました。 これらのデータは、食品由来のLb. plantarum菌株が、腸内共生のLb. plantarum菌の代表と同程度に、ヒトの腸内で見られる濃度よりも高い濃度の胆汁酸塩ストレスに耐える強い能力を示唆している。 |
 |
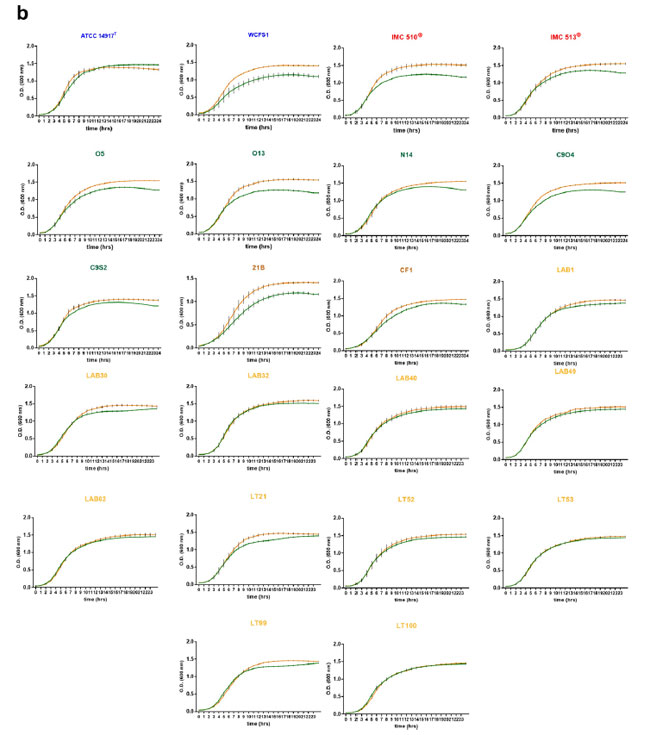 |
図 1 食品に関連するLb. plantarum株は胆汁酸塩に対して耐性があります。 (a) Lb. plantarum株の生存率を胆汁酸塩への曝露に関して評価しました。 データは平均値として表され、バーは 3 回の反復から得られた標準偏差 (SD) を示します。 データは一元配置分散分析に続いてBonferroni'sの多重比較事後検定によって分析されました (0% vs 1.8% p < 0.05、0% vs 3.6% p < 0.001)。 (b) 食物関連のLb. plantarum株は、胆汁の存在下で嫌気性増殖が可能です。 Lb. plantarum株を、胆汁の存在下および非存在下での嫌気条件下での増殖について評価した。 オレンジ色の線は MRS ブロスのみを示し、緑色の線は 0.5% ブタ胆汁 (0.5% w/v) を添加した MRS ブロスを示します。 データは、3 回の生物学的複製の平均からの標準偏差 (SD) を伴う平均として表されます。 |
| 胆汁酸塩はリン脂質とミセルを形成し(胆汁全体に含まれるため)、純粋な胆汁酸塩の人工溶液よりも抗菌活性が低い可能性がある(48)ことを考慮して、すべての菌株を0.5% w/v ブタ胆汁(胆嚢由来)を含む MRS ブロス中で培養して胆汁耐性を調査しました。 そして、細菌の増殖を24 時間にわたって1時間ごとに監視しました(図1bおよびSI図S1b)。 試験したすべての菌株は、好気条件および嫌気条件の両方で、胆汁にさらされている間、高い増殖能力と生存能力を示しました。 これらの条件下では、胆汁酸の存在下では 3 つのヒト分離株すべての増殖が実験期間中減少し、倍加時間 (DT は約 240 分) を示しました。 同様にLb. plantarum株O5、O13、C9O4(テーブルオリーブ由来)および21B(サワードウ由来)はすべて、ブタ胆嚢胆汁の存在下で倍加時間の減少(それぞれ270、240、240、および300 分)を示しました(図1b)。 これらの倍加時間は、有酸素条件下では同様でした (図 S1)。 これらのデータは、増殖の遅いこれらの菌株が、以前は腸内共生細菌に限定されていた機能である胆汁酸を代謝する能力を持っている可能性があることを示唆しています。 より疎水性の高い部分への変換は細菌の増殖を減少させる可能性があり、これは胃腸消化管において利点となる可能性があります。 このため、起源に関係なく、すべてのLb. plantarum株について、胆汁酸を代謝する特異的能力を調査しました。 |
| 2.2.細菌による胆汁酸プロファイルの変化:胆汁酸塩加水分解酵素脱抱合活性 |
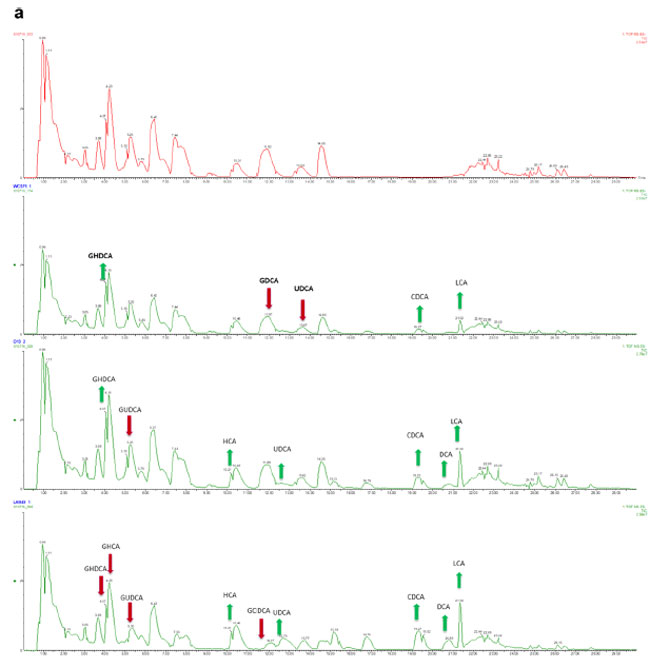
| 胆汁酸脱抱合能力または胆汁酸塩加水分解酵素活性を決定するために、標的メタボロミクスを22株すべてに適用し、30を超える胆汁酸部分が評価されました(表S1)。 in vitro共培養分析に由来する胆汁酸は、液液抽出手順によって抽出されました (詳細については補足情報を参照)。 Joyce らによって開発された方法(21 )に従って、胆汁酸抽出の効率は 2 つの重水素化内部標準を使用して評価されました。これにより、正確な胆汁酸の同定と各部分の検量線の高い直線性が確保され、ng レベルまでの信頼性の高い定量化が容易になりました(図 S2)。 UPLCクロマトグラムは、図2aに示すように、食品由来のLb. plantarum株が胆汁酸プロファイルを変化させる明確な能力を示しました。ここでは、いくつかの非抱合胆汁酸が生成されていることは明らかです(緑色の矢印)。 すべての胆汁酸の主成分分析(PCA)(図2b)により、微生物と胆汁の共培養により、これらのグループが分離して以来胆汁酸部分の変化が示されたことが確認されています。 これらは、腸環境から直接分離されていないにもかかわらず、すべての菌株が胆汁酸塩を脱結合させる能力を有することを示す対照反応とは異なっていた(図3)。 ヒトの腸および唾液に関連する菌株と比較して、食品から分離された Lb. plantarum株は、程度は異なるものの、二次胆汁酸であるデオキシコール酸、リトコール酸およびウルソデオキシコール酸を含む遊離胆汁酸を生成する能力において同等の効率を示した。 さらに、すべての菌株は明らかな糖特異的脱結合活性を示しますが、タウロ結合胆汁酸のレベルは大きく変化しません (図 4)。 興味深いことに、胆汁酸塩加水分解酵素対立遺伝子の存在について菌株を調べたところ、これらの菌株が保有する 胆汁酸塩加水分解酵素の数は、数 (1 ~ 4 胆汁酸塩加水分解酵素) と長さが異なりました (データは示さず)。 それらの活性レベルは、特定の 胆汁酸塩加水分解酵素対立遺伝子の存在に依存しているようには見えません。 |
 |
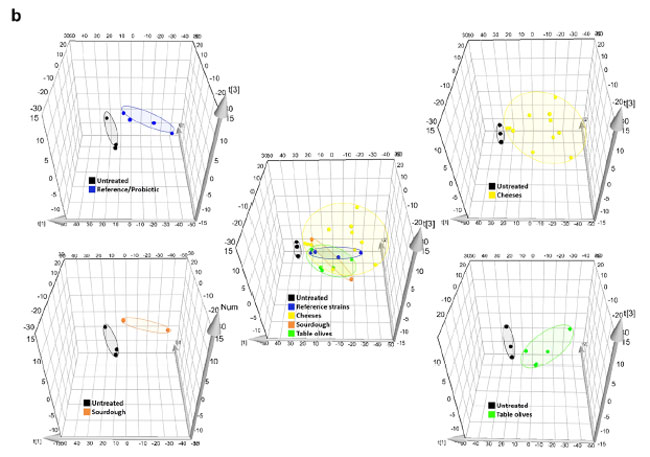 |
図 2 食品関連の Lb. plantarum 株は示差的な胆汁酸代謝能力を示します。 (a) 変化因子 (FC) によって設計された食品由来のLb. plantarum株による代表的な胆汁酸部分の変化を示す UPLC TMS クロマトグラム。 変化因子は、各胆汁酸変化の処理サンプルと未処理サンプルの平均値を比較することによって計算されました。 上から: 未処理サンプル、ヒト Lb. plantarum WCFS1 処理サンプル、食品由来 Lb. plantarum O13 処理サンプル。 食品由来のLb. plantarum LAB49 処理サンプル。 (b) 主成分分析 (PCA) 分析 (MassLynx ソフトウェア V4.2 SCN943 (WATERS Corporation, USA) を使用して生成) は、次のような未処理および Lb. plantarum処理胆汁酸調整を表します: 異なる起源ごとにグループ化された Lb. plantarum株: 参照/プロバイオティクス ( 青)、チーズ(黄色)、サワードウ(オレンジ)、テーブルオリーブ(緑)。 |
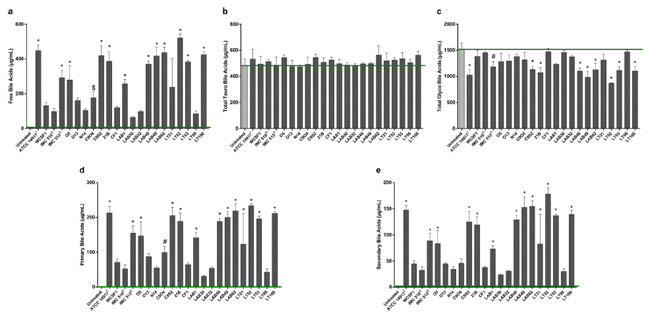 |
| 図 3 Lb. plantarum株による胆汁酸プロファイルの変化。 共培養上清中の UPLC Q-TOF-MSE によって評価。 緑色のバーは、未処理のブタ胆汁中の胆汁酸の基礎レベルを示します。 (a) 遊離胆汁酸。 (b) 総タウロ結合胆汁酸 (c) 総糖結合胆汁酸。 (d) 一次胆汁酸の総レベル (e) 二次胆汁酸の総レベル。 すべてのデータは平均 ± SD として表されます。 統計分析は、一元配置分散分析に続いてBonferroni'sの多重比較事後検定によって実行されました ($p < 0.05、#p < 0.01、および *p < 0.001)。 |
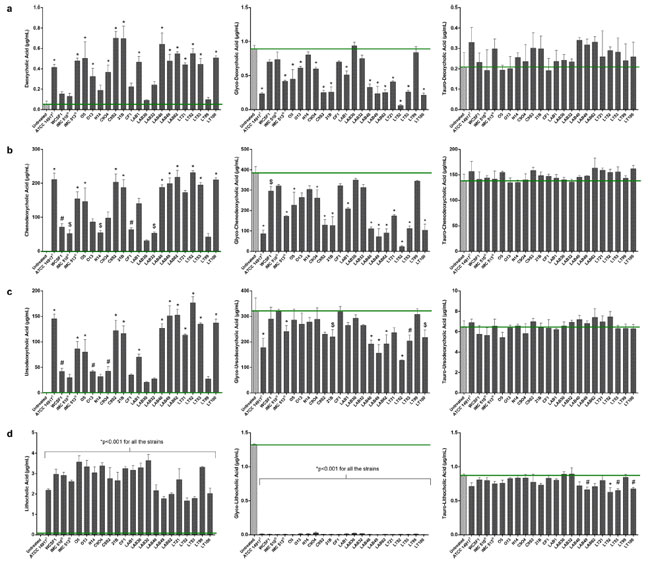 |
| 図4 Lb. plantarum株の胆汁酸塩加水分解酵素活性。 代表的な胆汁酸部分として、(a) デオキシコール酸、(b) ケノデオキシコール酸、(c) ウルソデオキシコール酸、および (d) リトコール酸の遊離型、複合糖質型およびタウロ複合体型があり、基礎胆汁酸と比較して非抱合胆汁酸の糖特異的生成が高いことが示されています。 未処理のブタ胆汁中のレベル (緑色のバー)。 データは平均値 ± SD として表示されます。 統計分析は、一元配置分散分析に続いてBonferroni'sの多重比較事後検定によって実行されました ($p < 0.05、#p < 0.01、および *p < 0.001)。 |
| 遊離胆汁酸を生成する分離株の脱共役強度は次のとおりです: ATCC14917 ≥ C9S2 ≥ LT52 ≥ LT100 ≥ LAB62 ≥ LAB49 ≥ LAB40 ≥ C9O4 > IMC513 > O5 > LAB1 > LT21 > WCFS1 ≧ N14 ≧ CF1 ≧ LAB32 > Lab30。 これらのデータは、成長の遅さは脱共役の程度によるものではなく、むしろ生成される特定の胆汁酸のレベルによるものである可能性があることを示しています。 この点に関して、UPLC Q-TOF-MSE メソッドは、いくつかの遊離胆汁酸、具体的には デオキシコール酸、ケノデオキシコール酸、ウルソデオキシコール酸、および リとコール酸 の高生産を検出しました。これらはすべて糖鎖結合活性から生成されました (図 5)。 胆汁酸脱結合プロファイルの変動は、株依存的な様式での選択的修飾を示しています。 特に、すべての株は、オキシコール酸、ケノデオキシコール酸、ウルソデオキシコール酸のレベルを生成する異なる能力を明らかに示し、IMC513はヒト関連株の中で最も効率的な株を表しています(図4)。 胆汁酸塩加水分解酵素活性における同様の違いは、同じ起源の食品関連菌株でも報告されています。 実際、2 つのサワードウ関連株、21B および CF1 は、異なるレベルの は デオキシコール酸、ケノデオキシコール酸および ウルソデオキシコール酸を生成する明確な活性を示しました (図 4)。 同様の効果は生乳チーズ株でも明らかであり、LAB30、LAB32、LT99 は同じ起源を持つ他のすべての株と比較して低い活性を示し、食品の種類と特定の B胆汁酸塩加水分解酵素活性の間に関係がないことが確認されました。 それにもかかわらず、この研究では、すべての食品関連分離株の間で広く見られる特性として、顕著な胆汁酸塩加水分解酵素活性とその結果としてのデオキシコール酸、ケノデオキシコール酸、ウルソデオキシコール酸、リとコール酸などの遊離胆汁酸の遊離を強調することが重要である。 |
 |
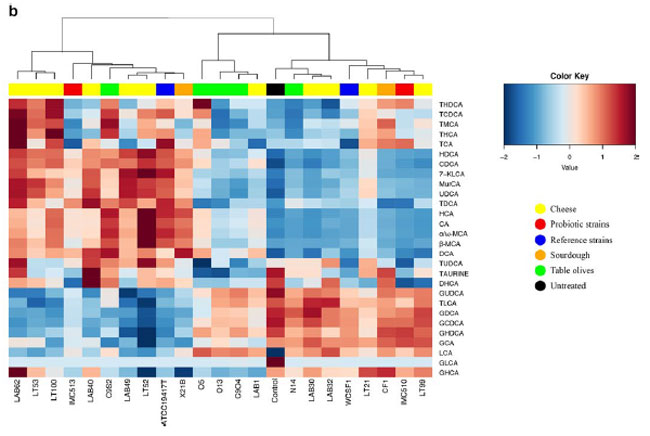 |
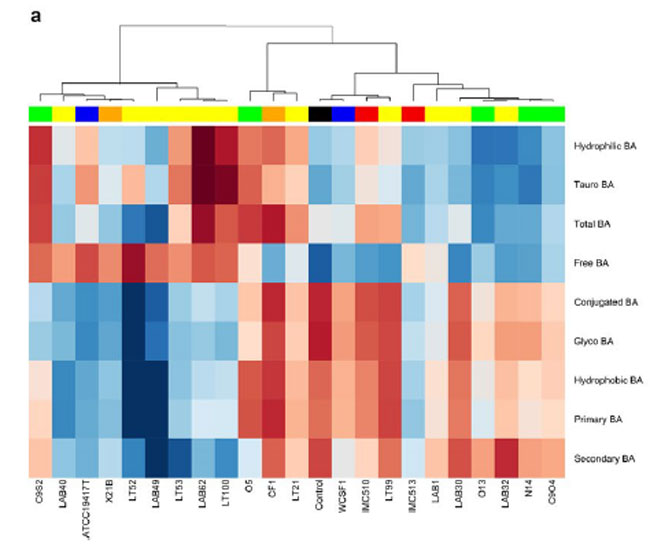
| 図 5 異なる起源の細菌株の胆汁酸塩加水分解酵素活性。 ヒートプロットは、胆汁酸クラス (a) および個々の 胆汁酸(b) の変化を示します。 ヒートマップは、RStudio ソフトウェア バージョン 1.2.1335 (http://www.rstudio.com) を使用して生成されました。 |
| 3.議論 |
| ヒトの消化管内で生存するために、微生物は、pHの変動、低酸素レベル、栄養素の制限、高浸透圧などの数多くの環境損傷に耐え、生き残る必要があります。 腸内環境における細菌の生存の観点からは、胆汁酸関連ストレスに耐える潜在的なプロバイオティクス株の能力を評価する必要があります(49,50)。 胆汁への曝露は腸内の深刻な問題を表します。 胆汁は脂質を乳化する消化液ですが、疎水性が高く、大量に摂取すると細胞膜の変化にも影響を与える可能性があるため、胆汁耐性は微生物の生存と胃腸消化管定着の両方にとって重要な特性です(4)。 胆汁は宿主特異的(51)および非特異的(52) 防御機構において基本的な役割を果たしており、この阻害効果の大きさは胆汁酸濃度に強く影響されます(53)。 ヒトの消化管では、平均胆汁濃度は 0.3% w/v であると考えられており、この濃度は胆汁耐性と胆汁耐性をスクリーニングするのに重要かつ十分であると考えられています(54)。 すべての食品関連の Lb. plantarum 株は 1.8% 胆汁の存在下でも生存を保ち、3.6% 胆汁に曝露された場合に増殖阻害を示したのは 3 株 (Lb. plantarum ATCC14917、CF1 および LT99) だけでした。 腸内の胆汁酸濃度は静的なものではなく、時間の経過とともに腸内で局所的に変化します。 これは食後に特に顕著であり、胆汁酸塩濃度は約 100 グラムまで急激に増加することがあります。 十二指腸では 15 mmol/L、回腸では活発な回腸胆汁酸再吸収により濃度は 4 mmol/L 未満になります(51)。 胆汁酸がリン脂質、コレステロール、脂肪酸(全胆汁)とミセルを形成していることを考慮すると、純粋な胆汁酸の人工溶液よりも抗菌活性が低い(48)。また、すべての菌株を 0.5% w/v ブタ胆汁(胆嚢の起源)で培養することにより胆汁耐性を調査しました。 全ての株は、嫌気性および好気性条件の両方でブタ胆汁に耐性があり、胆汁の存在下または非存在下で同様の増殖を示した(図1bおよびS1、図S1b)。 |
| これらのデータは、人間の消化管だけでなく、乳製品、肉、多くの野菜発酵を含む幅広い環境ニッチを占める多用途乳酸菌であるLb. plantarumの適応能力を裏付けています(55)。 Lb. plantarumは、乳酸菌の中で特徴付けられた最大のゲノムの 1 つ (3.3 Mb) を持ち、これはいくつかの環境ニッチを占有する生態学的柔軟性を反映している可能性があります(56)。 Lb. plantarum WCFS1 ゲノム(57 )には、bsh1 から bsh4 と注釈が付けられた 4 つの 胆汁酸塩加水分解酵素関連遺伝子が含まれると予測されており、それらはゲノム全体に広がっており、bsh1 はLb. plantarum株の胆汁酸代謝能力に単独で関与していると考えられます。 |
| 私たちのデータは、この研究に適用されたすべての菌株、食品由来の分離株とヒト分離株の両方が主要な胃腸ストレスの 1 つを生き延びる能力を示しており、これを in vitro 胃腸モデルでの以前の研究と組み合わせると、我々は提案します(46,47)すべての菌株が十二指腸に到達し、消化管を通過する際に生存できるはずです。 微生物の胆汁耐性はプロバイオティクス株の選択基準として認識されており、食事補助剤として使用する株を選択する場合には胆汁酸塩加水分解酵素活性が望ましい特性です(10、13、58)。 この点において、微生物の胆汁酸塩加水分解酵素活性は腸内での胆汁酸塩生体内変換の最初のステップです。 胆汁酸塩加水分解酵素活性は、腸内細菌の主要門および腸古細菌に分布する腸内微生物の共通の特徴と考えられています(8)。 さらに、この活性は、宿主の脂質代謝を機能的に調節し、コレステロール代謝において重要な役割を果たす微生物と宿主の対話を媒介する上で重要であると認識されている(16,59-61)。 微生物の 胆汁酸調節と代謝は宿主の生理機能に大きな影響を与えることが示されており (13)、マウスの体重と脂質代謝を調節する可能性を伴う血中コレステロールレベルの変化と関連付けられています (19,36,62,63)。 これは、宿主の体重増加に影響を与える新しい戦略として微生物の胆汁酸変化を強調しています(21,61)。 |
| 現在まで、発酵食品から分離された細菌による胆汁酸代謝を調べた文献はほとんどありません。 本研究では、食品由来の Lb. plantarum 株は、程度は異なるものの、ヒト分離株から報告されているものと同様のレベルで胆汁酸を代謝できます。 したがって、胆汁酸塩加水分解酵素の活動が腸内常在菌または侵入者である病原菌株に厳密に限定されているという考えは再考されるべきです。 この研究は、食品分離株間での 胆汁酸脱結合能の菌株特異的変動を強調しています (図 5)。食品の起源と 胆汁酸塩加水分解酵素活性の間には厳密な相関関係はありません。 さらに、この研究からの食品関連分離株は糖特異的脱抱合活性に適応しているが、タウロ抱合胆汁酸塩に作用する能力は有意ではないことを実証しました(図3)。 胆汁酸塩加水分解酵素陽性細菌は、結合基質に対して異なる親和性と相対活性を持っています(4)。 タウリン結合胆汁酸よりもグリシンを優先して代謝することも、Lb. plantarum WCFS1の特徴です。 この特異性は、ヒト胃腸管におけるグリシン結合胆汁酸の有病率の増加と、タウリン結合胆汁酸(64)と比較してより高い毒性が提案されていることを考慮すると、グリシン結合胆汁酸に対する進化的優先性を表している可能性がある。 胆汁酸アミデートが胆汁酸塩加水分解酵素によって選択される正確な機構は解明されていないが、糖結合またはタウロ結合した胆汁酸特異的基質に対する親和性の違いは、胆汁酸塩加水分解が生体内での細菌の生存にとって重要である可能性を示唆している。 |
| この研究から得られたLb. plantarum食品分離株は、タウロ結合型胆汁酸を比較的そのまま維持しながら、遊離胆汁酸デオキシコール酸、ケノデオキシコール酸、ウルソデオキシコール酸、および リとコール酸の蓄積をサポートします(図4および5)。 これらの遊離胆汁酸は、核胆汁酸受容体ファルネソイドX受容体(FXR)やビタミンD受容体(VDR)を含む多くの臓器や組織(肝臓、胆嚢、腸)で発現される特定の宿主受容体の活性に影響を与えるリガンドです。 これは、宿主における胆汁酸の消化とシグナル伝達の二重の役割を示しています (16、59、60、65)。 これらの相互作用は、エネルギー代謝、肝機能、腸の健康に影響を与える可能性があり、インスリン感受性や体重減少も改善する可能性があります(16,21,66)。 ファルネソイドX受容体は、当社の菌株が蓄積を促進する特定の遊離胆汁酸および二次胆汁酸によって優先的に刺激されます。 ファルネソイドX受容体活性は結腸直腸がんに対して保護的であると考えられており、胆汁酸脱結合が ファルネソイドX受容体(67)の抗がん効果を増強する可能性があることを示唆しています。 タウリン結合二次胆汁酸による TGR5(訳者注:TGR5とは、ヒトの消化管内に発現している胆汁酸受容体である)の活性化:TDCA および TLCA、ここで報告されている Lb. plantarum分離株の存在下で無傷のまま残される結合体、胆汁酸活性化 TGR5 は白色脂肪組織の褐変を増加させる可能性があり、チロキシンの生成を誘導してエネルギー消費を増加させ、それによって食事誘発性の肥満に影響を与えます(68)。 胆汁酸はまた、全身のグルコースおよび脂質代謝、ひいては体重の調節に寄与すると提案されています(61)。 胆汁酸プールの細菌による調節、特にファルネソイドX受容体(および TGR5 シグナル伝達の調節は、宿主の恒常性ならびに脂質およびコレステロールの代謝の調節に基本的な役割を果たしていることが広く認識されています(9,61)。 さらに、胆汁酸プロファイルの調節は末梢概日時計にも関連していることが判明しており、これは食物摂取や肥満と相関している可能性がある興味深い側面です(69)。 したがって、微生物の胆汁酸塩加水分解酵素活性は、肝硬変、脂肪肝疾患、胆汁うっ滞、結腸直腸がん、特定の種類のヘリコバクター関連がん、ならびに肥満および関連する代謝性疾患を含む胆汁酸の腸内毒素症を伴う関連する症状の新しい予防および治療戦略にとって重要である可能性がある有益な特性と考えられる可能性があります。 |
| 結論として、これは腸内環境に存続するための適応メカニズムとして、胆汁酸を代謝する能力について発酵食品から単離されたLb. plantarum株のコレクションを調査した最初の研究の1つである。 ここで調べた食品由来のすべての Lb. plantarum株は、腸に関連していないにもかかわらず、ヒト分離株と同レベルの胆汁酸塩加水分解酵素活性を有することが示されました。 胆汁酸脱共役プロファイルの変動は、糖共役Lb. plantarumを特定の基質として利用する明確かつ顕著な能力を備えた、株依存的な微妙な選択的修飾を示しました。 すべての分離株における胆汁酸塩加水分解活性活性は、デオキシコール酸、ケノデオキシコール酸、ウルソデオキシコール酸、リとコール酸などの遊離胆汁酸の蓄積を引き起こし、これらはすべて、全身のグルコースおよび脂質代謝の調節に寄与する胆汁酸シグナル伝達分子として認識されています。 |
| 宿主の観点から見ると、この微生物の活動は、胆汁酸プールと宿主受容体のシグナル伝達を調節することにより、宿主の代謝プロセスの変化を引き起こす可能性があります。 さらに、この研究で報告された実験的証拠は、胆汁酸塩加水分解酵素活性が腸関連微生物のみに特有の特徴であるという概念を再考する必要があることを示唆しています。 全体として、この研究は、発酵食品の微生物成分(Lb. plantarum種として)による胆汁酸代謝への有益な寄与と、宿主の代謝プロセスを調節する可能性を強調しています。 最後に、この研究は、食品由来の微生物が腸内で生存するための装備をある程度備えていることを強調しています。 微生物は通常、消化管内での増殖を促進する可能性のある食品とともに導入されることを考慮すると、我々のデータは、消化管通過を通じて食品に関連する非病原性微生物を監視および追跡する将来の研究の重要性を示しています。 安定した定着と宿主との相互作用からの一過性を判断し、その増殖と食物と腸内環境の両方への遺伝的適応を監視するためです。 |
| 4.方法 |
| 4.1.細菌株 |
| この研究では、テラモ大学の実験室培養コレクションに属する 22 株のLactobacillus plantarumのコレクションを調査しました。 Lb. plantarum WCFS1とATCC14917、および実証されたプロバイオティクス活性を持つ2つの菌株を除き、すべての菌株はもともと生乳チーズ(11菌株)、テーブルオリーブ(5菌株)およびサワードウ(2菌株)として発酵食品から分離されました。 IMC 510と IMC 513 (イタリア、カメリーノの SYNBIOTECH よりご提供) は参照株として研究に含まれています (表 1)。 すべての株は以前に Lb. plantarum 種として特徴付けられており、胃腸通過で生存する潜在的な能力を含むいくつかの特性についてテストされました (46,47)。 これらは、de Man, Rogosa and Sharp (MRS) 培地 (Oxoid) を使用し、37°Cで微好気条件下で日常的に増殖されました。 |
| 表 1 この研究で調査したLb. plantarum株とその起源と供給源の食物を示します。 |
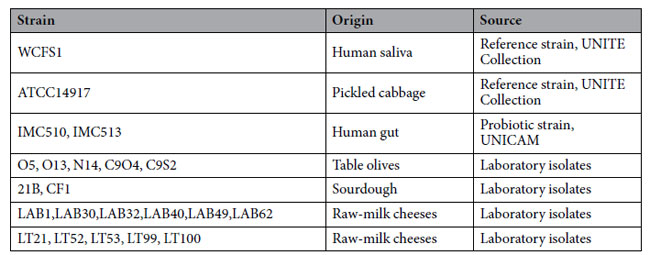 |
| 4.2.胆汁酸塩耐性 |
| 微生物の胆汁酸塩耐性を調べるために、Gilliland らによって記載された修正方法を使用して、分光光度分析が実施されました(51)。 分離株をMRSブロス中で37 ℃で増殖させ、一晩培養した。 次に、段階希釈液の 0.5 ml を MRS ブロスに添加して、初期接種濃度 103 CFU/ml を取得し、すべての株を MRS ブロス中で 37 °C で 24 時間、胆汁酸塩 (OXOID) の濃度を増加させながら共培養しました (0 %、0.30%、0.60%、1.8%、3.6% w/v)。 24 時間の培養後、600 nmでの光学密度を測定することにより、微生物の増殖を濁度測定的に評価しました(図1a)。 |
| 4.3.胆汁耐性 |
| 微生物の胆汁耐性は、胆嚢ブタ胆汁の存在下での増殖をモニタリングすることによって各菌株について評価した。 全ての菌株を、ブタ胆嚢胆汁(0.5% w/v)を含むMRSブロス中で、好気条件および嫌気条件の両方で培養し、細菌の増殖を追跡するために、O.D. 600 nmで24 時間毎時間撮影されました(図1bおよび図S1、図S1b)。 |
| 4.4胆汁酸塩加水分解酵素脱抱合活性 |
| 胆汁酸を脱結合させる細菌の能力は、同時in vitro 分析を実行し、その後、MSE モード (UPLC Q-TOF-MSE) で動作するエレクトロスプレーイオン化四重極飛行時間型質量分析計と組み合わせたウルトラパフォーマンス液体クロマトグラフィーを実行することによって調査されました。 簡単に言うと、すべての Lb. plantarum 株をブタ胆嚢胆汁 (0.5% w/v) 中で 90 分間共培養し、その後、液液抽出を実行してすべての胆汁酸を回収しました (表 S1)。 Joyce ら (21 )によれば、UPLC Q-TOF-MSE を実行して 胆汁酸プロファイルを分析しました。 研究では、Lb. casei NCDO161 を bsh 陰性対照として使用しました (図 S3)。 実験の詳細については、SI Materials and Methods(70、71)を参照してください。 |
| 4.5.統計分析 |
| 3 つの生物学的複製からのデータは、標準偏差付きの平均値として報告され、一元配置分散分析 (ANOVA) およびスチューデント t 検定によって統計的に分析されました。 ひずみ平均間の差異は、PRISM 7.0 (GRAPHPAD Software Inc.、La Jolla, CA) を使用した Bonferroni の多重比較検定によって有意性について検定されました。 胆汁酸プロファイルの微生物変化の主成分分析は、MassLynx Software V4.2 SCN943 (WATERS Corporation, USA)を使用して実施した。 |
| 5.補足情報 |
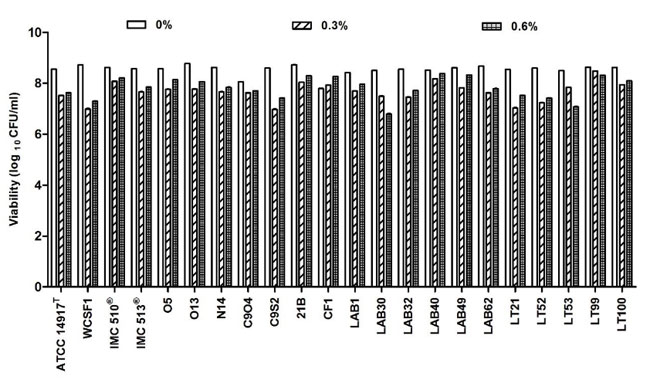 |
| 図S1a 胆汁酸塩曝露に対するLb. plantarum株の細胞生存率。 データは平均値として示され、棒は 3 回の反復から得られた標準偏差を示します。 一元配置分散分析とそれに続くBonferroni'sの多重比較事後テストでは、有意な差は示されませんでした。 |
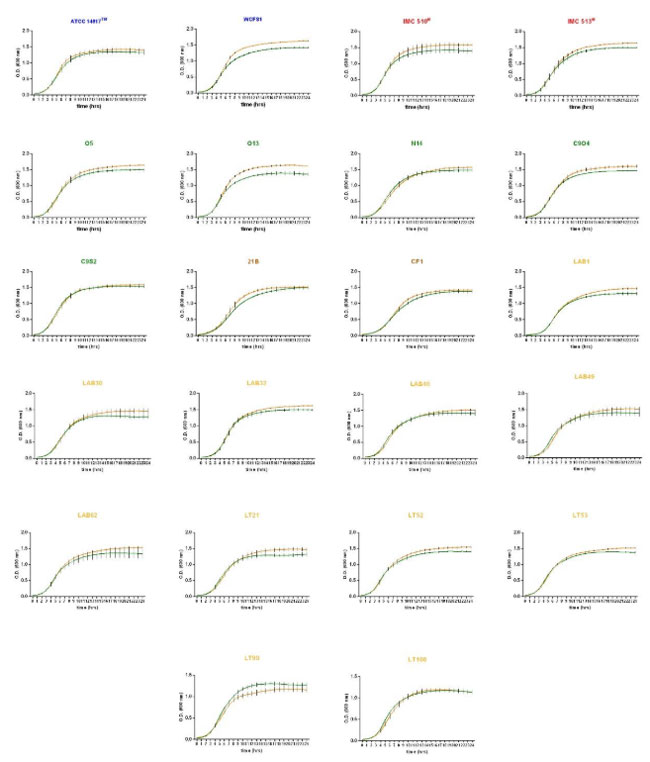 |
| 図S1b 好気性胆汁耐性増殖曲線 (オレンジ色の線は MRS ブロスを意味し、緑色の線はブタ胆汁 (0.5% w/v) を含む MRS ブロスを意味します。データは 3 つの生物学的複製からの平均標準偏差として報告されます。 |
 |
 |
| 図S2 標準曲線は、各胆汁酸部分の変化を定量化するために使用されました。標準希釈を利用して、検査した各分析物の標準曲線を作成しました (n=3)。 内部重水素化標準により、抽出効率の評価とサンプル間の正規化が容易になりました。 |
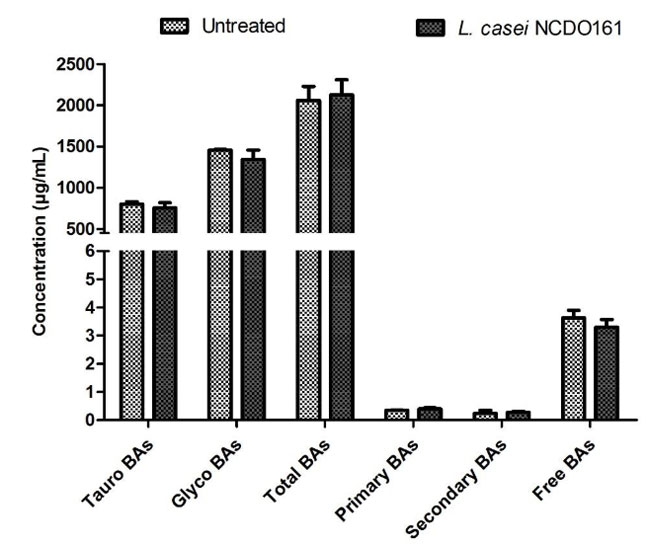 |
| 図S3 研究の bsh 陰性対照として、Lb. casei NCDO161 による胆汁酸の変化を調べました。 胆汁酸の特徴は UPLC Q-TOF-MSE によって評価され、データは標準偏差 (SD) を伴う平均値として報告されます。 Student t 検定と二元配置分散分析 (Bonferroni の事後検定) の両方を使用して統計比較を実行しましたが、未処理のサンプルと比較した場合、有意差は見つかりませんでした (p = 0.9758 および p = 0.5650)。 |
| 表S1 この研究では、UPLC Q-TOF-MSE によって胆汁酸部分を分析し、その中性式、質量、標準曲線 R2 値および範囲を検出しました。 |
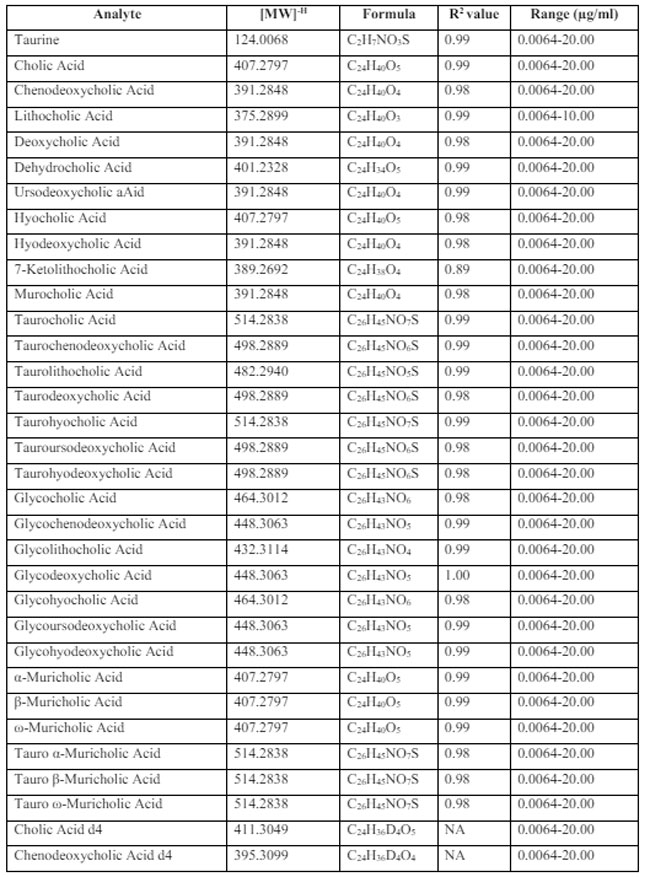 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
| この文献はSci Rep. 2020; 10: 1165.に掲載されたBeneficial bile acid metabolism from Lactobacillus plantarum of food origin.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |