Valentin Sencio et al., |
人間の胃腸管にコロニーを形成する細菌は、健康のために不可欠です。 腸内細菌叢は、肺の免疫とウイルス性呼吸器感染症に対する宿主の防御に重要な役割を果たします。 腸内細菌叢の組成と機能は、急性感染症を含む多くの病気の状況で深刻な影響を受ける可能性があり、これらの変化は病気の重症度を悪化させる可能性があります。 ここでは、腸内細菌叢がウイルス性呼吸器感染症を制御するために肺を武装させるメカニズムについて説明します。 ウイルス性呼吸器感染症が腸内細菌叢に及ぼす影響を要約し、腸内細菌叢の組成と機能の変化につながる潜在的なメカニズムについて説明します。 また、胃腸障害や二次細菌感染症などの疾患の転帰に対する腸内微生物の不均衡の影響についても説明します。 最後に、2019年のコロナウイルス病における肺腸軸の潜在的な役割について説明します。 |
前書き |
消化管は、一般に腸内細菌叢と呼ばれる複雑で非常に多様な微生物生態系(主に偏性嫌気性細菌)を宿しています。 まとめると、腸内細菌は300万を超える遺伝子のゲノムを構成します。これはヒトゲノムの150倍です(1)。 腸内細菌叢は、ファーミキューテス(ラクトバチルス、バチルス、クロストリジウムなど)、バクテロイデス門(バクテロイデスなど)、および程度は低いがプロテオバクテリア(エシェリキアなど)およびアクチノバクテリア(ビフィズス菌など)によって支配されています。 実験的および臨床的証拠は、腸内細菌叢が人間の健康の維持に重要な役割を果たしていることを示しています。 厳密に制御された微生物叢と宿主の相互作用は、免疫系の発達、命令、およびプライミングに影響を与えます(3–6)。 最近の調査研究では、この相互作用が感染の制御に大きな役割を果たしていることが示されています(7)。 腸内細菌叢が免疫応答を調節するメカニズムは、微生物に関連する分子パターン、微生物の代謝産物、および微生物と前駆細胞および成熟免疫細胞との相互作用に依存します(8–10)。 |
| 多くの要因が腸内細菌叢の多様性と組成を変化させ、腸内毒素症を引き起こす可能性があります。 これには、食事パターンや広域抗生物質や抗がん剤などのストレス誘発剤が含まれます。 感染症や慢性の炎症性または代謝性障害などの病気の状況も、腸内毒素症につながる可能性があります(11)。 腸内細菌群集の変化は、異生物微生物叢を用いた移動実験で示されているように、離れた臓器(肺を含む)でも病気の結果に影響を与える可能性があります(8–11)。 ここでは、腸内細菌叢がウイルス感染に対する肺の防御を遠隔的に促進するメカニズムを要約します。 腸内細菌叢に対する急性ウイルス性呼吸器感染症の影響をレビューし、腸内毒素症の潜在的なメカニズムについて議論し、腸内細菌叢の変化が二次疾患の転帰に及ぼす影響について議論します。 コロナウイルス病-19(COVID-19)の結果における腸-肺軸の潜在的な重要性は、この知識に照らして議論されます。 |
ウイルス性呼吸器感染症に対する 肺の防御における腸内細菌叢 |
腸内病原菌へのコロニー形成に対する耐性における腸内細菌叢の役割は十分に確立されています(7)。 したがって、抗生物質によって特に誘発される微生物叢の変化は、多かれ少なかれ完全に腸内細菌または特定の種を根絶し、サルモネラ菌や腸内凝集性大腸菌などの腸内感染症に対する感受性を高めます。 免疫における腸内細菌叢のこの有益な効果は、腸内だけでなく、全身や肺などの遠隔臓器にも及びます(12,13)。 微生物叢を欠くマウス(すなわち、無菌)または抗生物質(広域スペクトルまたはネオマイシン、メトロニダゾール、バンコマイシンなどの標的抗生物質)で経口投与されたマウスは、全身感染症および呼吸器感染症に対する反応が損なわれています。 たとえば、A型インフルエンザウイルス(IAV)や呼吸器合胞体ウイルス(RSV)によって引き起こされるウイルス性呼吸器感染症の状況では、化学療法による腸内細菌叢の生態系の深刻な障害が、宿主の先天的および適応的防御を弱めます(26)。 腸内細菌叢が肺を武装させてウイルス性呼吸器感染症を制御するメカニズムは数多くあります。 これらの点は、このレビューでは広範囲にレビューされませんが、いくつかの例が提供されます(レビュー、27–30)。 簡単に言えば、いくつかの研究は、腸内細菌叢が肺のI型インターフェロン(IFN)の産生に及ぼす影響を強調しています(15,16,18,31)。 COVID-19の病因である重症急性呼吸器症候群コロナウイルス2(SARS-CoV-2)を含むウイルス感染を制御することがよく知られています(32–34)。 この設定では、デスアミノチロシン(フラボノイドおよびアミノ酸代謝に由来)や短鎖脂肪酸(SCFA、共生細菌による食物繊維発酵の最終産物)などの微生物代謝産物が重要であることが示されています(15,18)。 たとえば、偏性嫌気性菌(クロストリジウム・オルビスシンデンス、メトロニダゾール、バンコマイシン感受性)が植物フラボノイドの消化から生成するデスアミノチロシンは、 インフルエンザ感染から保護するために、血液中に拡散し、肺に到達し、(自然)免疫系を刺激する可能性があります(18)。 デスアミノチロシンは肺におけるインターフェロン刺激遺伝子の合成を促進し、肺食細胞はこのメカニズムにおいて重要でした。 この設定では、デスアミノチロシンは、インターフェロン-α/β受容体およびシグナル伝達物質および転写活性化因子を介したインターフェロン増幅によってI型インタフェロンシグナル伝達を増強しました(1.18)。 短鎖脂肪酸の場合、ラクノスピラ科のメンバーを含む無数の共生生物が食物繊維を発酵させてつくることができます。 たとえば、酢酸塩は血液中に拡散し、間質細胞によって発現されるGタンパク質共役型受容体(GPR)43(遊離脂肪酸受容体2とも呼ばれる)を活性化できることが示されました。 この活性化経路は、I型インターフェロン応答を改善し、インターフェロン刺激遺伝子発現を増加させました(15)。 GPR43がI型インターフェロンを介したインフルエンザ感染からの防御を促進するメカニズムを解明する必要があります。 微生物代謝物に加えて、Toll様受容体(TLR)リガンドを含む、無傷の腸内細菌叢に由来する微生物膜成分も、ウイルス性呼吸器感染症に対する宿主防御において重要な役割を果たします。 実際、ポリ(I:C)(TLR3アゴニスト)またはペプチドグリカン(TLR2アゴニスト)の局所および遠位への接種は、抗生物質で治療されたマウスの免疫障害を救済しました。 腸内細菌叢から発せられるこれらの遠位信号に加えて、上気道からの局所微生物も下気道免疫を調節し、ウイルス感染に対する肺防御に役割を果たすことは注目に値します。 メカニズムは他の場所で説明および/またはレビューされており(24,29)、このレビューでは説明しません。 腸内細菌叢は、自然免疫応答に影響を与えるだけでなく、CD8 + T細胞エフェクター機能を促進します。これは、ウイルス(インフルエンザ)のクリアランスにも関与するプロセスです(17,25,31)。 上記のように、抗生物質で腸内細菌叢を変えると、インフルエンザなどのウイルス性呼吸器感染症の重症度が高まりました。 それどころか、高繊維食で微生物叢を刺激すると、逆の効果がありました(15,31)。 インフルエンザの場合、高繊維(発酵性イヌリン)食は2つの異なる経路を通じて保護をもたらしました。 食物繊維の発酵は骨髄造血の変化をもたらし、呼吸器合胞体ウイルス感染マウスの肺に代替的に活性化されたマクロファージの蓄積をもたらしました(31)。 これらのマクロファージは、ケモカイン(C-X-Cモチーフ)リガンド1の産生が少ないため、気道への初期の好中球浸潤を低下させます。 この経路は、誇張された肺の炎症と損傷を大幅に軽減しました。 食事はまた、CD8 + T細胞の代謝を促進し、CD8 + T細胞のエフェクター機能を強化し、ウイルスクリアランスを効果的に強化しました。 保護効果にはGPR41(遊離脂肪酸受容体3とも呼ばれる)が必要であり、保護を与えるには短鎖脂肪酸(酪酸)の経口投与で十分でした(31)。 呼吸器合胞体ウイルス感染時にも同様の保護効果が観察されています(15)。 高繊維(発酵性ペクチン)食は、肺上皮細胞のI型インターフェロン応答を調節し、肺のインターフェロン刺激遺伝子の発現を増加させることにより、呼吸器合胞体ウイルス感染から保護しました。 保護効果にはGPR43が必要であり、保護を与えるには酢酸塩の経口投与で十分でした(15) 。 |
| 肺で抗ウイルス効果を発揮する腸内共生細菌の性質はまだとらえどころのないです。 上記のように、クロストリジウム菌嫌気性菌(デスアミノチロシン)のメンバーと短鎖脂肪酸生産菌は、肺の自然免疫系のプライミングに重要です15,18。 最近では、感受性動物と耐性動物の比較に基づく研究により、ビフィズス菌属(ビフィドバクテリウム・シュドロンガムおよびビフィドバクテリウム・アニマルス)によるコロニー形成がインフルエンザ感染マウスの生存と強く関連していることが示されました(35)。 近い将来、他の共生メンバーが特定され、重度のウイルス性呼吸器感染症を経験している患者の重症度と死亡率を予測するための新しいバイオマーカーとして機能する可能性があります。 したがって、定常状態では、腸内細菌叢からの常在細菌は、ウイルス性呼吸器感染症と戦うために自然にそして遠隔で肺を強化することができます。 腸内細菌叢がSARS-CoV-2のようなコロナウイルスによる感染の初期制御(自然免疫)と後期制御(適応免疫)にも関与しているかどうかはまだ決定されていません。 SARS-CoV-2感染の結果に対する腸内細菌叢の枯渇(抗生物質による)の影響は、さらなる研究を待っています。 呼吸器ウイルスに対する肺の防御における腸内細菌叢の役割に関しては、微生物叢の組成と機能が変化すると、腸と肺の間の有益なクロストークが変化する可能性があります(図1)。 |
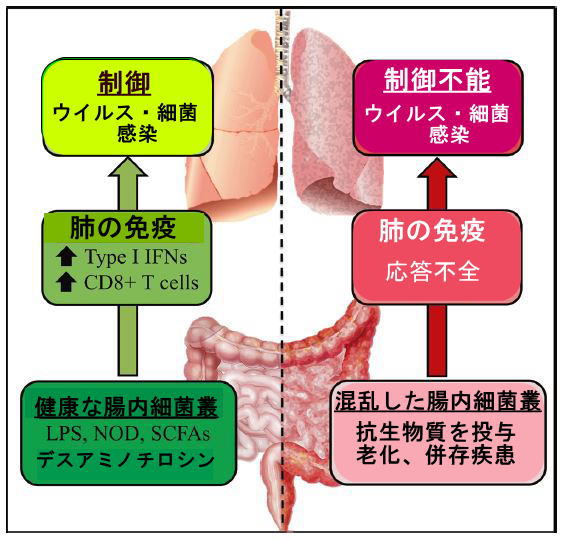 |
図1ウイルス性呼吸器感染症における 腸内細菌叢の役割 |
| 健康な腸内細菌叢によって放出される因子は、ウイルス性呼吸器感染症に対して肺を武装させます。 対照的に、抗生物質治療は腸内の生態学的平衡を乱し、肺の防御を変化させます。 加齢と併存疾患は、腸内細菌叢の機能の変化と呼吸器感染症への感受性の向上にも関連しています。 |
| 腸内細菌叢の変化に関与する要因と状態の中には、抗生物質の使用、急性および慢性疾患、および老化があります。 肥満、糖尿病、慢性呼吸器疾患などの併存疾患、および加齢は、腸内細菌叢の組成と機能の混乱(多様性の低下など)に関連しています(6,36,37)。 これらの状況では、機能不全の腸内細菌叢が呼吸器ウイルスに対する肺の防御に遠位に影響を与える可能性があります。 これを実証するには、糞便微生物叢移植実験(例えば、老化した腸内細菌叢の若いマウスへの移植)が必要になります。 |
| 「腸-肺軸」は双方向であり、肺疾患(感染症を含む)は腸内細菌叢の組成に影響を与える可能性があります。 過去数年にわたって、多くの研究者が肺感染症の動物モデルを使用して、腸内細菌叢に対する呼吸器ウイルス(主にA型インフルエンザウイルスや呼吸器合胞体ウイルス)の影響を分析してきました。 ウイルス性呼吸器感染症が肺自身の微生物叢に及ぼす影響は、本レビューの範囲外です。 |
| インフルエンザ感染と腸内細菌叢 |
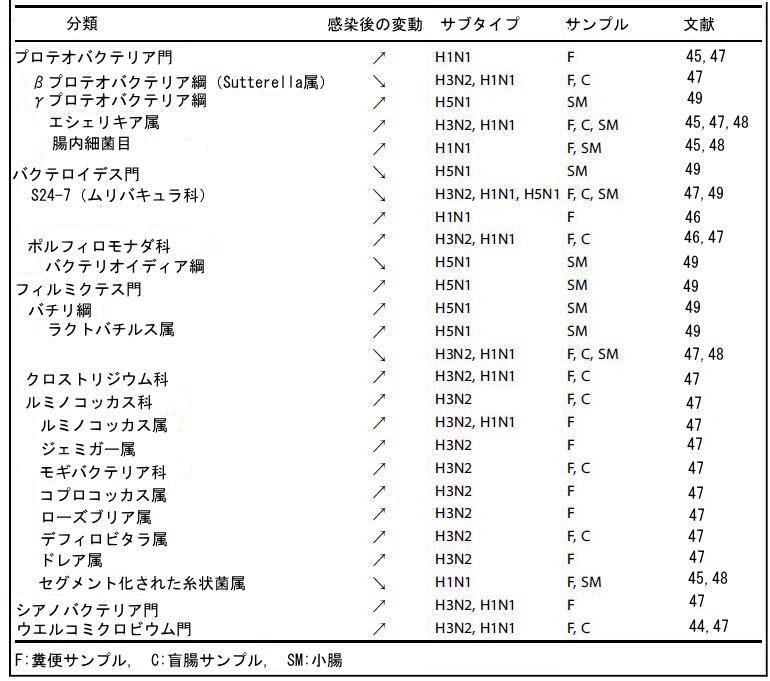
| インフルエンザ感染症は重大な罹患率と死亡率の原因であり、世界中で毎年300万から500万人の重症例と、29万人から65万人の死亡があります(38)。 インフルエンザはまた、壊滅的なパンデミックを引き起こす可能性があります。 1918年と2009年のパンデミックにより、それぞれ5,000万人と250,000〜500,000人が死亡しました(39–41)。 インフルエンザ感染症は、軽度から重度の肺炎、急性呼吸不全として臨床的に定義される急性呼吸窮迫症候群(ARDS)、および多臓器不全を引き起こす可能性があります(38–40,42)。 腸内にウイルスが見られないにもかかわらず、腹痛、吐き気、嘔吐、下痢などの胃腸炎のような症状を示す患者もいます(43)。 これは、インフルエンザ感染時に肺と腸の軸が潜在的に重要であることを示唆しています。 いくつかの研究グループは、主に16S rRNAシーケンスによって、マウスのA型インフルエンザウイルス感染の状況における微生物の変化の性質を研究しました(35,44–49)。 小腸、盲腸、結腸など、いくつかのA型インフルエンザウイルスサブタイプ(H1N1、H3N2、H5N1、H7N9)と部位が研究されています。 データは収束しています。つまり、腸内細菌叢の組成の変化は、感染後7〜9日でピークに達します。 腸内でウイルスは検出されず、弱毒化ウイルスの接種後に腸内細菌叢の組成に変化は観察されませんでした(46)。 後者の発見は、腸内細菌叢の変化が生きたウイルス感染によって特異的に引き起こされ、信号が局所的な複製によるものではないことを示唆しました。 腸内細菌叢の組成の変化は一過性であり、違いはまだ存在していましたが、組成は感染後14日までに全体的に初期状態に戻りました(47,49)。 運用分類単位の数によって評価される細菌負荷とアルファ多様性は、A型インフルエンザウイルス感染の影響を強く受けませんでした(47,49)。 門レベルでは、ほとんど変化は観察されませんでした。 A型インフルエンザウイルスのH1N1サブタイプによる感染は、バクテロイデス門/ファーミキューテス門の比率の低下につながりました。 これは主に、S24-7(ムリバキュラ科としても知られている)とポルフィロモナダ科の増加によるものでした(46)(表1)。 対照的に、H3N2およびH5N1感染は、S24-7科の減少を引き起こしました(47,49)。 ウェルコミクロビウム(主にアッケルマンシア属によって構成される)の増加は、H3N2およびH1N1サブタイプによる感染中にも観察されました(44,47)。 これらの細菌は腸の粘液層を分解することが知られており、ルミノコッカスも同様に、A型インフルエンザウイルス感染中にその数が増加しました。 放線菌の減少(主にビフィドバクテリア属の減少による)もA型インフルエンザウイルス感染後に観察されています(47)。 より広い変化は、より低い分類学的レベルで観察されています。 簡単に言えば、ほとんどの研究で、ガンマプロテオバクテリア(病原性大腸菌の可能性がある)が増加し、まれにラクトバチルス属(バチリ綱)が減少することがわかりました(45,47–49)(表1)。 微生物叢分析はまた、ファーミキューテス門内の変化を浮き彫りにしました。 要約すると、クロストリジウム(非関連)、ルミノコッカス科、およびモギバクテリア科、およびコプロコッカス、ローズブリア、デフルビタレア、ドレア、ルミノコッカス、およびジェミガー属は、感染中に比較的豊富でした。 さらに、マウスのA型インフルエンザウイルス感染は、セグメント化された糸状菌(クロストリジウム科)の割合の低下と関連していた(45,48)。 この細菌は腸上皮と密接に相互作用し、シトロバクター・ロデンチウムなどの腸内病原菌に対する宿主の耐性に重要です(50)。 全体として、インフルエンザのマウスモデルからのデータは、感染がガンマプロテオバクテリアのメンバーや粘液分解細菌などの潜在的に有害な細菌種の出現を促進することを示しました。 逆に、感染は、乳酸桿菌、ビフィズス菌、セグメント細菌などの健康促進細菌の増殖を鈍らせるように見えます(48)。 |
| 興味深いことに、最近の研究では、A型インフルエンザウイルス(H7N9)感染による腸内細菌叢の変化の性質が、感染に屈したマウスと感染を生き延びたマウスで異なることが示されました(35)。 特に、ビフィドバクテリウム・シュドロンガムおよびビフィドバクテリウム・アニマルスのレベルは、死亡したマウスと比較した場合、生存しているマウスで有意に上昇しました。 ビフィズス菌は、バリンや補酵素Aを含むいくつかの特定の代謝分子を介して抗インフルエンザ効果を媒介することが示唆されました(35)。 |
 |
| 驚くべきことに、人間の環境でのウイルス性呼吸器感染症の間の腸内細菌叢の変化の性質を分析した研究はほとんどありません。 Quinらは、H9N2鳥インフルエンザに感染した40人の患者のサンプルを分析しました(51)。 多様性の減少と大腸菌とエンテロコッカス・フェシウムの異常増殖が観察されました。 ユーバクテリウム、ルミノコッカス、ビフィズス菌、およびローズブリアはすべて、感染した患者ではそれほど豊富ではありませんでした。24人のインフルエンザA(H1N1)患者のコホートで、Guらは、放線菌、エリシペロトリキア、クロストリジウム、および有益な酪酸生産菌(ラクノスピラ科およびルミノコッカス科)の相対的な存在量の減少を報告しました(52)。 一方、赤痢菌やエシェリキア属などの日和見病原菌が発生しました。 大規模なヒトコホートの腸内細菌叢に対するインフルエンザ感染の影響の研究は現在保証されています。 |
| 腸内細菌叢の代謝物は健康に重要です。 A型インフルエンザウイルス感染中の腸内細菌叢の組成の変化が腸の代謝出力を変化させるかどうかが最近取り上げられました。 マウスシステムでは、A型インフルエンザウイルスの感染により腸(盲腸)メタボロームが変化し、短鎖脂肪酸の産生が大幅に低下しました(47)。 酢酸塩、プロピオン酸塩、酪酸塩の濃度はすべて、模擬感染した対照よりも低かった。 A型インフルエンザウイルスの感染は、糖脂質代謝の変化ももたらしました。 興味深いことに、A型インフルエンザウイルス感染は、不変のナチュラルキラーT細胞のリガンドとして知られており、げっ歯類やヒトの免疫系で重要な役割を果たすα-ガラクトシルセラミド53の合成を変化させました(54)。 これらの代謝変化が腸の恒常性と機能に影響を与えるかどうかは明らかではありません。 しかし、α-ガラクトシルセラミドの利用可能性の低下は、腸の不変ナチュラルキラーT細胞の免疫調節機能を妨げる可能性があると推測できます(55)。 (不変の)ナチュラルキラーT細胞のリガンドに加えて、粘膜関連不変のT(MAIT)細胞のリガンドの合成もウイルス性呼吸器感染中に変化する可能性があります。 実際、バクテロイデス門およびプロテオバクテリア門に属する種を含む細菌共生菌、ならびに病原性細菌は、MAIT細胞のアゴニストまたはアンタゴニストとして作用することが知られているビタミンB2およびビタミンB9代謝産物を産生することができる。 粘膜免疫におけるMAIT細胞の重要な役割に関しては、ウイルス性呼吸器感染症の際のMAIT細胞リガンドの合成の変化は、疾患の転帰に重要な結果をもたらす可能性があり(56-58)、この仮説を調査する必要があります。 |
| 呼吸器合胞体ウイルスと腸内細菌叢 |
| 呼吸器合胞体ウイルスは、2歳未満の小児における細気管支炎および肺炎の最も一般的な原因です。 小児下気道感染症の80%以上を占めています(59)。 この病原体は、高齢者の呼吸器疾患の重要な原因でもあります 。A型インフルエンザウイルス感染で観察されたように、呼吸器合胞体ウイルスは季節的な発生を引き起こし、世界中で毎年最大118,000人が死亡しています(60)。 マウスモデルでは、呼吸器合胞体ウイルスにより、感染後7日目に腸内細菌叢の多様性が大幅に変化し(バクテロイデス門が増加し、ファーミキューテス門が減少しました)(46)。 バクテロイデス門のこの増加は、主にバクテロイデス門とS24-7科の増加によるものでしたが、ファーミキューテス門の数の減少は、ラクノスピラ科と乳酸桿菌科の両方の弱毒化に関連していました。 腸内細菌叢の代謝活動に対する呼吸器合胞体ウイルス感染の影響はめったに扱われていません。 ある研究では、呼吸器合胞体ウイルス感染は主に、スフィンゴ脂質、多価不飽和脂肪酸、短鎖脂肪酸、吉草酸などの脂質の代謝を増加させました(61)。 |
| コロナウイルスと腸内細菌叢 |
| コロナウイルスは人間の集団を継続的に循環し、一般的に風邪を含む軽度の呼吸器疾患を引き起こします。 対照的に、SARS-CoVと中東呼吸器症候群コロナウイルスはどちらも人獣共通感染症であり、重症呼吸器疾患を引き起こし、高い死亡率を示す可能性があります(62)。 SARS-CoVおよび中東呼吸器症候群コロナウイルスに対する特定の抗ウイルス薬または承認されたワクチンはまだ利用できません。 2019年後半、中国の武漢で新たな感染性呼吸器疾患が発生しました(63,64)。 この病気(現在はSARS-CoV-2によって引き起こされるCOVID-19と呼ばれています)は、中国や世界中の他の多くの国々に急速に広がりました(64)。 |
| 驚くべきことに、腸内細菌叢に対するコロナウイルス感染の影響に取り組んだ研究はほとんどなく、それらのほとんどは鶏と豚のモデル(微生物の不均衡の誘発)に焦点を当てていました(65–69)。 最近の研究では、SARS-CoV-2感染がヒトの腸内細菌叢に及ぼす影響が報告されています(52,70–73)。 簡単に言えば、SARS-CoV-2感染は、ルミノコッカス科およびラクノスピラ科(ローズブリア)科のいくつかの属などの酪酸生産菌の存在量を低下させました(71)。 ストレプトコッカス(クラス バチリ)、ロチア、およびアクチノマイセスを含む日和見細菌性病原体の有意に高い相対的存在量も観察されました。 別の最近の研究では、RNAショットガンメタゲノミクスシーケンシングにより、SARS-CoV-2感染性しているCOVID-19の高い患者の糞便サンプルに、コリンセラ・アエロファシエンスやモーガネラ・モーガニ、ストレプトコッカス・インファンチス(上気道に豊富なコロニー形成菌)などの日和見細菌性病原体が存在することが明らかになりました(73)。 対照的に、短鎖脂肪酸とトリプトファン生産菌は糞便サンプルに富んでおり、SARS-CoV-2感染性が低いかゼロかを示しています。 興味深いことに、SARS-CoV-2の感染力が高い糞便は、ヌクレオチドのデノボ生合成、アミノ酸生合成、解糖に対してより高いミクロビオーム機能能力を持っていました(73)。 注目すべきことに、日和見真菌病原体(アスペルギルスおよびカンジダ属)の異常増殖もCOVID-19患者で報告されました(72)。 Guの研究では、インフルエンザ(H1N1)患者とSARS-CoV-2患者の類似点と相違点が説明されていることに注目するのは興味深いことです(52)。 中核となる腸内微生物の特徴、および関連する代謝物の発見は、潜在的な診断マーカーとして役立つ可能性があります。 |
| 展望 |
| 動物モデルと臨床データは、急性ウイルス性呼吸器感染症が、バランスの取れた腸内微生物生態系の一時的な破壊につながり、有益な細菌が少なく、病原性細菌が多いことを明確に示しています。 ただし、より包括的な説明を取得する必要があります。 さらなる研究には、ショットガンメタゲノミクスシーケンスなどのメタゲノミクス分析を含める必要があります。 後者のアプローチは、真菌、ウイルス、寄生虫の組成の潜在的な変化を分析し、微生物の代謝経路の主要な変化を評価する可能性を提供します(74)。 並行して、大規模な人間のコホートとより関連性の高い動物モデル(たとえば、人間以外の霊長類)が必要です。 これらのアプローチは、調査結果の潜在的な臨床的関連性を判断するのに役立ちます。 同じ方針に沿って、腸内細菌叢の代謝出力に対する急性ウイルス性呼吸器感染症の全体的な影響をよりよく理解することが緊急に必要とされています。 この分野の研究は、ウイルス性呼吸器感染症との関連でまだ始まったばかりです。 メタボロミクス研究は、腸内細菌叢の変化が疾患の転帰に及ぼす影響のより詳細な理解を提供する可能性があり、また、疾患の重症度のバイオマーカーの発見につながる可能性があります。 ゲノムスケールの代謝モデリングは、腸内細菌叢の代謝能力を推定するのにも役立ちます。 腸内細菌叢の組成、微生物代謝物、炎症性サイトカインのレベル、および疾患の重症度の変化の間の相関分析は非常に有益です。 このアプローチは、特にウイルス性呼吸器感染症に非常にかかりやすい患者の間で、介入の予防/治療標的として役立つ可能性があります。 |
ウイルス性呼吸器感染症中の 腸内細菌叢組成の変化につながるメカニズム |
| 急性ウイルス性呼吸器感染症の際の腸内細菌叢の変化にはいくつかの原因があります。 これらには、炎症性サイトカインの放出や食物摂取量の減少が含まれる可能性があります(図2)。 | 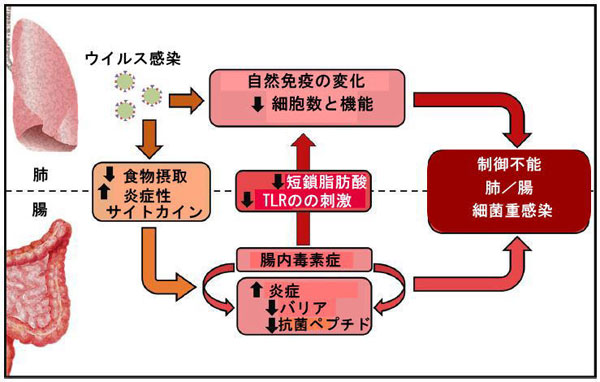 |
| インフルエンザなどのウイルス性呼吸器感染症は、肺の免疫細胞と間質(上皮)細胞の機能を変化させ、二次的な細菌感染症を引き起こします。 一方、全身性炎症性サイトカイン、食欲不振(食物繊維の減少)、酸素レベルの変化、および上皮代謝の変化は、腸内細菌叢の組成と機能を破壊します。 これらの変化は、腸の炎症、上皮バリアの破壊、および抗菌ペプチド(AMP)の産生の減少に寄与します。 その後、二次腸感染症が発症します。 上皮漏出は、細菌の移動を促進し、全身性炎症および臓器機能障害を引き起こす可能性があります。 ウイルス性呼吸器感染症による腸内毒素症は、Toll様受容体(TLR)やヌクレオチドオリゴマー化ドメイン(NOD)様アゴニスト、短鎖脂肪酸(SCFA)などの微生物代謝物などの微生物関連分子パターンの産生を低下させ、抗菌性肺免疫を低下させます。 したがって、腸の恒常性を変えることにより、呼吸器ウイルス感染症は細菌の重感染につながります。 肺と腸の相互接続性は、SARS-CoV-2感染時に特に関係がある可能性があります。 |
マウスでは、A型インフルエンザウイルスまたは呼吸器合胞体ウイルスによる感染は、初期体重の10〜20%の範囲で、感染後5〜7日でピークに達する大幅な体重減少を引き起こします(47,61,75,76)。 この体重減少は主に食欲不振(食欲不振)によるものです(77,78)。 食物とカロリー摂取量の減少は腸内細菌叢を混乱させることが知られています(79)。 ペアフィーディング実験の結果は、次のことを明確に示しています。 食物摂取量の急激な減少は、感染中に観察された腸内細菌叢の変化を模倣します(すなわち、ウェルコミクロビウム門、アルファプロテオバクテリアクラス、およびパラバクテロイデス属の存在量の増加とラクノスピラ科、ルミノコッカス、およびラクトバチルス属の存在量の減少)も観察された(47,61)。 食物繊維摂取量の減少と一致して、ペア飼育マウスは短鎖脂肪酸の濃度が低かった(47)。 それらの中で、酪酸は結腸上皮細胞に燃料を供給し、上皮の酸素消費を促進するため、腸管腔での酸素欠乏を可能にします。 食物(繊維)摂取量の減少による短鎖脂肪酸の低下は、上皮細胞の代謝を変化させる可能性があります。 ウイルス感染時の食欲不振の主な説明の1つは、腫瘍壊死因子α-80を含む炎症性サイトカインの過剰産生です。 呼吸器合胞体ウイルス感染中のこのサイトカインの中和は、体重減少を減らし、腸内細菌叢の摂動を部分的に弱めました(61)。 同じ方針に沿って、呼吸器合胞体ウイルス感染中にサイトカイン産生CD8 + T細胞を枯渇させると、食欲不振が減少し、腸内細菌叢の変化が逆転しました(61)。 I型およびII型インターフェロンは、特にインフルエンザの際に、宿主の抗ウイルス反応に不可欠な要素です(81)。 これらのサイトカインは腸内細菌叢を強力に妨害しました(45,48)。 野生型マウスと比較して、I型インターフェロン受容体を欠損したマウスは、プロテオバクテリア(エッシェリシャ属)の開花の減少を示し、逆に、セグメント化された糸状菌の割合が増加しました(45)。 同じ方針に沿って、A型インフルエンザウイルス感染中のインタフェロン-γの枯渇は、セグメント化された糸状菌(インターフェロン-γコンピテントマウスで減少)、ラクトバチルス属(減少)、および腸内細菌科(増加)の変化を回復させました(48)。 したがって、炎症性サイトカインと食欲不振は、少なくともマウスのシステムでは、ウイルス性呼吸器感染症の間に腸内毒素症を引き起こす可能性があります。 他のメカニズムも関係している可能性があります。たとえば、浸潤したCD4 + T細胞または全身のインターフェロン放出による腸の炎症は、上皮細胞の代謝を変化させることが知られており(45,48)、その結果、腸管腔に生息する微生物が競合する新しい栄養素のセットが蓄積されます。 酸素の利用可能性の増加(以下を参照)とともに、この現象は、偏性嫌気性菌からプロテオバクテリア腸内細菌科などの通性嫌気性菌への移行を説明しているようで(82–86)。 低酸素症は、COVID-19患者を含む、呼吸器ウイルス感染の急性期における主要な臨床症状です(87)。また、慢性的な腸の損傷にも関連しています。 マイクロバイオータの組成と機能を含む腸内恒常性における酸素の役割に関して、例えば短鎖脂肪酸の利用可能性による酸素勾配の崩壊は、呼吸器ウイルス感染時の腸内毒素症と胃腸障害に関与している可能性があります(88)。 呼吸器ウイルス感染中の腸(上皮)免疫機能の欠陥(抗菌ペプチドの産生の低下)も腸内毒素症に関与している可能性があります。 |
| A型インフルエンザウイルスおよび呼吸器合胞体ウイルスの状況とは対照的に、ウイルスRNAは、SARS-CoV-2感染を含むコロナウイルス感染中に腸内で検出されます(89–92)。 COVID-19患者の約半数は、気道にもはや見られない場合でも、便中にSARS-CoV-2 RNAを持っています(90–92)。 重要なことに、感染性ウイルスがCOVID-19患者の糞便サンプルで検出され、消化管がウイルスの複製と活動の部位である可能性があることを示唆しています(93,94)。 並んで、ヒト小腸オルガノイドを使用したいくつかの最近の研究は、SARS-CoV-2が腸細胞で複製することを示しました(93,95,96)。 したがって、局所的なウイルス複製は、腸内細菌叢の組成と機能の変化につながる局所的な生態系を乱す可能性があります。 さらに、ウイルススパイクタンパク質が細胞表面受容体アンジオテンシン変換酵素IIに結合すると、後者の発現がダウンレギュレーションされます(97)。 腸の微生物生態学の維持におけるアンジオテンシン変換酵素IIの重要な役割に関して(アミノ酸輸送の機能不全、トリプトファン欠乏、および抗菌ペプチドの産生の減少を介して)(98)。 SARS-CoV-2感染時に利用可能なアンジオテンシン変換酵素IIが不足していることが、腸内毒素症に重大な役割を果たしている可能性があります。 |
呼吸器ウイルスによって誘発された 腸内細菌叢の変化が疾患の転帰に及ぼす影響 |
糞便移動実験は、インフルエンザ(H7N9)感染を生き延びたマウスから収集された腸内細菌叢が、A型インフルエンザウイルス.35でチャレンジされたレシピエント(ナイーブ)マウスの保護を移動できることを示しました。 この保護効果が他のA型インフルエンザウイルスサブタイプや他の呼吸器ウイルスにまで及ぶかどうかはまだ不明であり、今後の研究に値します。 したがって、呼吸器ウイルス感染による腸内細菌叢の組成と機能的活動の変化は、有益な効果につながる可能性があります。 ただし、以下で説明するように、急性呼吸器ウイルス感染による腸内毒素症も有害な影響をもたらし、感染の結果に関与します。 |
| 腸の恒常性への影響 |
腸内細菌叢は、上皮の完全性の維持と制御性T細胞の発達に重要です(1–8)。 この重要な役割は、主に微生物叢の代謝物に依存しています。 恒常性の間、腸は比較的抗炎症性の免疫状態を発揮します(99)。 腸内細菌叢の混乱は、炎症性腸疾患、過敏性腸症候群、セリアック病、結腸直腸癌などのいくつかの腸疾患の一因となります(100,101)。 これらの観察に基づいて、研究者らは、ウイルス性呼吸器感染症の間に観察された腸内毒素症が腸の恒常性と機能に及ぼす可能性のある悪影響を調査しました。 |
| 上記のように、A型インフルエンザウイルス、呼吸器合胞体ウイルス、コロナウイルス感染症などのウイルス性呼吸器感染症は、腹痛、吐き気、嘔吐、下痢などの胃腸炎のような症状を引き起こす可能性があります(43,102,103)。 A型インフルエンザウイルスに感染したマウスは、結腸の長さの減少(炎症のマーカー)、小腸の粘膜層の除去、軽度の下痢などの腸の損傷を示しました(48,49)。 並行して、炎症性サイトカインとインターフェロン刺激遺伝子のmRNA発現の増強が観察されました(45,48)。 腸の重要な特徴の1つは、そのバリア機能です。健康な条件下では、これは微生物成分の過度の拡散を防ぎます。 多くの病気はバリア機能の崩壊に関連しており、したがって細菌の転座、全身性炎症、およびショックを引き起こします。 腸のバリア特性に対するA型インフルエンザウイルスの影響は、詳細には特徴付けられておらず(104)、調査に値します。 呼吸器合胞体ウイルスの場合、腸の炎症のマーカーであるリポカリン-2の糞便レベルの上昇を除いて、結腸の有意な炎症の組織学的証拠はマウスモデルで観察されませんでした(46)。 |
| ウイルス性呼吸器感染症の状況において、変化した微生物叢が腸の炎症を引き起こすのか、それとも炎症が腸内毒素症を引き起こすのかを疑問視することができます。 これは「鶏が先か卵が先か」の状況です。 上記のように、炎症は腸内細菌叢の組成に影響を与える可能性があります。 一方、異生物性微生物叢は、腸内障害を直接開始し、維持する可能性があります。 実際、A型インフルエンザウイルス感染中の抗生物質治療(腸内細菌科の増加を制限するため)は腸の炎症を軽減しました(48)。 さらに、糞便移動実験は、A型インフルエンザウイルスを経験した微生物叢がTヘルパー17細胞の局所蓄積を引き起こし、その後腸の炎症を引き起こすことを示しました(48)。 腸内細菌科に加えて、粘液分解細菌(ルミノコッカスやアッケルマンシアなど)が腸の炎症の維持および/または増幅における重要な要因である可能性もあります。 実際、これらの細菌は結腸粘膜を侵食し、管腔細菌と腸上皮の間の相互作用を促進することが知られています。 次に、これは炎症とバリア機能の障害につながります(105,106)。 |
| 全体として、ウイルス性呼吸器感染症は、おそらく共生の平衡の変化を介して、腸の恒常性に影響を及ぼします。 腸の障害の性質と、腸の生理機能(摂取した食物や液体の消化と吸収など)、代謝、免疫恒常性に対する感染の影響については、詳細に分析する必要があります。 ウイルス性呼吸器感染症後の腸機能の変化は、健康に即時および長期の両方の結果をもたらす可能性があります。 たとえば、腸のバリアは恒常性の維持に不可欠であり、腸内細菌叢の混乱は細菌成分の全身的な拡散、ひいては有害な健康上の結果につながる可能性があります。 |
| 二次腸内感染の結果 |
腸内細菌叢は、直接的な微生物拮抗作用と宿主のエフェクター応答(抗菌ペプチドなど)の刺激を通じて、病原性感染症の局所制御に不可欠です(107,108)。 腸内毒素症は、腸内細菌感染症を発症するリスクを高める可能性があります(109,110)。 動物モデルでの実験は、A型インフルエンザウイルスがサルモネラ菌血清型ティフィムリウムによる二次腸内感染に対する感受性を高めることを示しました。 この増強は、少なくとも部分的には、I型インターフェロンとそれに関連する抗菌ペプチドおよび炎症性サイトカイン放出の減少によるものでした(45)。 腸内細菌叢の多様性の低下は、二次腸内感染にも関与している可能性があります(49)。 たとえば(マウスシステムでは)、セグメント化された糸状菌(Tヘルパー17を介した免疫応答で重要であることが知られている)の割合は、A型インフルエンザウイルス感染中に低下しました(50)。 並行して、短鎖脂肪酸生産菌の割合の低下は、二次腸内感染において重要である可能性があります(47)。 これらの脂肪酸は、腸内細菌叢の平衡を制御し、エンテロコッカス・フェカリスやサルモネラ菌血清型ティフィムリムなどの腸内病原菌の発生を防ぐことが知られています(111,112)。 A型インフルエンザウイルス感染中の短鎖脂肪酸の経口補給が二次腸感染を低下させることができるかどうかはまだ分からない。 |
| 肺細菌の重感染の結果 |
呼吸器ウイルス感染症の主な合併症の1つは、二次細菌感染症(主に肺炎球菌、黄色ブドウ球菌、インフルエンザ菌によって誘発される感染症)に対する感受性の増加です。 この合併症は主に子供と高齢者に影響を及ぼし(113,114)、エピデミックとパンデミックの間に観察される過剰な罹患率と死亡率の一因となります(42)。 動物モデルでの実験は、感受性の増強が、抗菌性の自然免疫応答の障害に部分的に起因することを示しました(図2)(42,115)。 |
| 健康な腸内細菌叢は、肺炎球菌や黄色ブドウ球菌などによる日和見細菌の発生を制御する積極的な役割を含め、肺免疫において有益で生理学的な役割を果たします(19,21,23,47,116)。 最近、インフルエンザ感染時の腸内細菌叢の混乱が肺の細菌性重感染の発生率を増加させる可能性を調査しました。 実際、糞便移動実験は、A型インフルエンザウイルスで調整された微生物叢が肺炎球菌感染症に対する肺の防御を危うくすることを示しました(図2)(47)。 メカニズムの用語では、酢酸塩(微生物叢によって生成される主な短鎖脂肪酸)の生成の低下は、肺胞マクロファージの殺菌活性に影響を与えました。 インフルエンザと肺炎球菌の二重感染の状況では、酢酸塩の補給は細菌の負荷を減らし、肺疾患を減らし、生存率を高めました。 したがって、インフルエンザ中の腸内細菌叢の変化(および短鎖脂肪酸産生の除去)は、細菌の重感染に関連しています。 この分野でのさらなる研究は、予測マーカー(例えば、全身性短鎖脂肪酸)を定義し、および/または、例えばプレバイオティクスおよび/またはプロバイオティクスを介して腸内細菌叢の力を利用することにより、細菌の重感染に対する治療アプローチを開発するのに役立つ可能性があります。 注目すべきことに、腸内細菌叢とともに、急性ウイルス性呼吸器感染症(レビュー117)に関連する気道の微生物組成と機能の変化が細菌の重感染において重要である可能性もあります。 この局所的な腸内毒素症は、微生物間の相互作用のダイナミクスを変化させ、それによって潜在的に病原性の細菌種の増殖を促進する可能性があります。 局所的な微生物代謝出力の変化も、二次細菌感染に対する肺の防御を低下させる可能性があります。 まとめると、ウイルス性呼吸器感染症(少なくともインフルエンザ)中の腸内毒素症は、肺の細菌性重感染に影響を及ぼします。 微生物の変化が、急性呼吸窮迫症候群(ARDS)、敗血症、多臓器不全などのウイルス性呼吸器感染症の際に他の疾患の転帰に影響を与えるかどうかは、依然として未解決の問題です。 |
| SARS-CoVの場合 |
COVID-19の進行と重症度に対する腸内毒素症の結果はまだ完全に特徴付けられていません。 胃腸症状のあるCOVID-19患者は、全体的に重症/重篤な疾患を患っており、この状況での肺-腸軸の重要性を示しています(118,119)。 入手可能な証拠は、SARS-CoV-2感染が腸のバリアを変化させ、細菌、内毒素、および微生物代謝物の全身拡散を引き起こすことを示唆しています(63,120,121)。 これは、COVID-19感染に対する宿主の反応に影響を与える可能性があり、多系統機能障害、敗血症性ショック、およびSARS-CoV-2感染の第2段階で発生し、疾患の死亡率の一部の原因となる全身性炎症性ストームの一因となる可能性があります。 SARS-CoV-2感染中の腸障害は、重症COVID-19患者に発症する、付随的または二次的な細菌感染にも関与している可能性があります(122–124)。 COVID-19患者の胃腸障害は、急性呼吸窮迫症候群、肝障害、体温上昇、ショックなど、より積極的な臨床経過と関連しています(118)。 腸の障害の性質、特に微生物叢の機能と腸のバリア特性は、標的療法を開発するために緊急に調査する必要があります。 COVID-19の危険因子(例えば、加齢、および肥満や糖尿病などの代謝性疾患)は特に重要である可能性があります(125)。 実際、肥満と糖尿病は、腸内細菌叢の障害と腸のバリア機能の障害に関連していることが知られています(6,36,37)。 腸の恒常性に対するウイルスの影響と組み合わせると、これらの慢性的な障害はCOVID-19の重症度を増幅させる可能性があります。 |
結論 |
腸内細菌叢は、A型インフルエンザウイルスおよび呼吸器合胞体ウイルス感染症の場合に例示されるように、呼吸器感染症に対する肺の防御に不可欠です。 これがSARS-CoV-2などの他の危険な呼吸器ウイルスにも当てはまるかどうかはまだ分からない。 関連する動物モデルを使用すると、ウイルス性病原体に対して肺を強化できる介入戦略を設計するための重要な情報が迅速に生成されます。 これらの戦略には、食物繊維が豊富な食事など、短鎖脂肪酸を生成するプロバイオティクスやプレバイオティクスの使用が含まれる場合があります。 これらの手順は、免疫系を予防的に武装させる腸内細菌叢の能力を強化し、ウイルス感染に関連する腸内細菌叢の混乱から微生物叢を保護する可能性があります。 腸内細菌叢の変化は、腸の障害に関与するだけでなく、全身の損傷や細菌の重感染にも有利に働きます。 この分野の研究は、ウイルス性呼吸器感染症との闘いを改善するための予防的および治療的アプローチにつながる可能性があります。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
| この文献は、Mucosal Immunology (2021) 14:296–304に掲載されたThe lung–gut axis during viral respiratory infections: the impact of gut dysbiosis on secondary disease outcomes. を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |