Jia-Sin Ong et al. |
概要 |
| 安全に使用されてきた長い歴史のおかげで、典型的にはLactobacillus属に由来するプロバイオティック微生物は、特に伝統的な発酵食品産業で長い間認識されてきました。従来、乳製品、肉、野菜の発酵に使用されていましたが、健康食品、サプリメント、栄養補助食品でのプロバイオティクスの使用が徐々に増加しています。.過去 20 年間で、腸の健康と免疫を改善し、代謝性疾患を緩和する上でのプロバイオティクスの重要性が認識されてきました。腸 - 心 - 脳軸の新しい概念は、食品や食事へのプロバイオティクスの導入に関連するさまざまな革新と戦略の開発につながりました。プロバイオティクスは、腸内微生物叢のプロファイル、炎症、障害に影響を与え、脳の神経伝達物質経路に直接影響を与えます。脳の健康は年齢とともに低下することが多いため、プロバイオティクスが老化した脳に有益であるという概念は、研究と製品開発の両方で勢いを増し、強調されています。このレビューでは、脳の老化の概念、さまざまな in vivo 老化モデル、プロバイオティクスの老化に関連するさまざまな利点について説明します。 |
| 目次(クリックして記事にアクセスできます) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.老化する脳 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.老化の原因 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1.テロメア短縮 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.2.酸化ストレス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.3.エネルギー恒常性の不均衡 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.老化研究で使用されるモデル | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.1.ヒト以外の霊長類 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.2.げっ歯類 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.3.ショウジョウバエ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.4.単細胞真核生物 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.プロバイオティクス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.腸-脳軸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7.腸内微生物叢と腸-脳軸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.神経化学レベルの変化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9.免疫調節反応の変化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.加齢に伴う生理的変化とプロバイオティクスの役割 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.1.腸内細菌叢 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.2.炎症 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.3.酸化ストレス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 11.サイコバイオティクス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1.老化の序説 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 老化は、時間の経過に伴う機能的能力の直線的かつ進行性の喪失 (暦年齢) として定義されます。加齢に伴うパフォーマンスや生産性などのフィットネス要素の低下を引き起こす複雑なプロセスが特徴です (Lipsky and King, 2015)。細菌から線虫、脊椎動物に至るまで、地球上のほぼすべての生物が老化の影響を受けています。加齢は身体能力の低下と関連しているだけでなく、メンタルヘルスや細胞プロセスにも影響を与え、最終的には罹患率と死亡率につながります (Ferrucci et al., 2008)。世界保健機関 (WHO) によると、暦年齢が 65 歳の人は、高齢者(elderly)および老齢者(old)と呼ばれます (Kowal and Dowd、2001 年)。 2017 年には、全世界で約 9 億 6,200 万人が 60 歳以上で、世界の総人口の 13% に相当します (国立衛生研究所、2016 年)。加齢に伴い、体力や協調性の低下、視力や聴力の低下などの加齢に伴う病気、糖尿病や心血管疾患などの代謝障害、アルツハイマー病やパーキンソン病などの神経障害を経験することがあります (Ferrucci et al., 2008)。 .人口の高齢化は、医療費の増加、労働参加率の低下、年金費用、および貧困による社会経済的負担にも寄与しています (国立衛生研究所、2016 年)。したがって、老化を遅らせたり、健康的な老化を促進したりする治療法を開発するには、老化の根底にある生物学的プロセスを理解することが不可欠です。今日まで、老化は世界中の科学者の間で最も議論されているトピックの 1 つです。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.老化する脳 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 加齢は、認知機能の低下や、不安などの行動障害と密接に関連しています。 中国、フランス、イタリア、韓国、米国で実施された 10 の人口調査によると、軽度認知障害の有病率は 3% から 42% の範囲です (Ward et al., 2012)。 高齢者によくみられる軽度認知障害は、特定の年齢や教育レベルで予想される以上の記憶、注意、および認知機能の低下を特徴としています (Eshkoor et al., 2015)。 記憶障害は、アルツハイマー病および認知症のマーカーと見なされています (Ritchie and Touchon, 2000)。 高齢者集団における認知症の有病率は、年間 1% から 2% です (Eshkoor et al., 2015)。 60 歳以上の人口における認知症の症例数は、9.4% (2000 年) から 23.5% (2050 年) に増加すると仮定されています (Eshkoor et al., 2015)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 52 歳から 88 歳までの 1,248 人の成人を対象とした研究では、聴覚言語学習テスト (学習能力のテスト) の成績が 58 歳を過ぎると低下し、処理速度と実行機能が 50 歳を過ぎると低下することがわかりました (Li et al., 2014)。老化が脳と認知に及ぼす影響は広範囲に及び、遺伝、神経伝達物質経路、ホルモン、経験など複数の原因が考えられます (Peters, 2006)。脳の構造的結合の変化、神経新生の減少、脂質過酸化、酸化ストレス、ミトコンドリアの機能不全、神経伝達物質レベルの低下、およびベータアミロイド (Aβ) の過剰産生も、脳の老化および老化の病因において主要な役割を果たすことが示唆されています。関連する神経障害 (Luo et al., 2009; Ali et al., 2015; Li et al., 2015)。これらの病因のほとんどは、加齢による活性酸素種 (ROS) の増加に関連しており、活性酸素種が脳の老化の主要なメディエーターであることを示唆しています。年齢に伴う DNA 修復メカニズムの障害も、加齢に伴う変性疾患の発症に寄与することが提案されています (Langie et al., 2017)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.老化の原因 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 老化に関する数十年にわたる研究を通じて、老化の原因に関する多くの理論が提唱され、老化が多因子プロセスであることは否定できなくなりました。 ほとんどの理論は、テロメア短縮、酸化ストレス、およびエネルギー恒常性の不均衡 (Weinert and Timiras,2003; Lipsky and King,2015; Sergiev et al.,2015) を中心に展開しており、これらはすべてこのレビューで説明されています。 全体として、老化は、細胞が生成された日に細胞にプログラムされているか、または成長に伴う損傷の蓄積を通じて発生するという 2 つの概念が浮上しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1.テロメア短縮 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| テロメアは、すべての線状染色体の末端に位置する高度に保存された反復 DNA 配列のセットです。それらは、靴ひものアグレットと同様に染色体の完全性を保護します (Weinert and Timiras, 2003; Lipsky and King, 2015)。テロメアは、細胞分裂の数を制限する主な要因です。すべての細胞は、細胞分裂の数が事前に決定され制限されている自然淘汰プロセスである複製老化を受けます。染色体の末端にあるテロメア DNA の一部は、細胞分裂のたびに消失し、テロメアの長さが非常に短くなると、細胞は複製老化を起こします (Weinert and Timiras, 2003; Heidinger et al., 2012)。短縮されたテロメアは、テロメア DNA を伸長することによって機能するテロメラーゼ酵素によって修復できます。しかし、テロメラーゼは、生殖細胞、幹細胞、がん細胞など、無限に増殖する能力を持つ細胞にしか見られません。細胞分裂のサイクルが限られている細胞の大部分は、テロメラーゼ活性を欠いており、その結果、テロメアの短縮を示しています (Sergiev et al., 2015)。したがって、テロメアの長さは、生物の寿命を決定するための体内時計としてよく使用されます。 Steenstrup et al (2017) による研究では、ヒトでは、テロメアが個人の自然な寿命限界の主要な決定要因である可能性があることがわかりました。 この研究は、2010 年のヒト死亡データベースによると、男性よりも約 5 年長く生きる女性は、通常、男性よりも長いテロメアを持っていることを証明しました (150 bp の差)。ゼブラフィッシュに関する別の研究でも、生後 25 日でテロメアが長い魚は長生きする傾向があり、生後 25 日で測定されたテロメアの長さが寿命と相関していました (Heidinger et al., 2012)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.2.酸化ストレス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 酸化ストレスまたはフリーラジカル説は、注意が必要な老化について最も議論されている説です。酸化ストレスは、老化を加速する最も破壊的なプロセスと考えられています。これは、反応性の高いスーパーオキシド アニオン、過酸化水素、ヒドロキシル ラジカルなどの活性酸素種によって顕著に引き起こされます (Cui et al., 2012)。活性酸素種は通常、代謝反応の副産物として生成されます。細胞の発電所であるミトコンドリアは、アデノシン三リン酸の形でエネルギーを生成し、電子伝達系を介して酸素を生成する役割を担っています。ミトコンドリアの呼吸中に、酸素が消費されて還元され、スーパーオキシドラジカルと過酸化水素が形成されます (Cui et al., 2012; Sergiev et al., 2015)。活性酸素種は、ストレス、放射線、化学酸化剤、炎症、毒素などの環境要因にも反応して生成されます。活性酸素種の有害な影響と老化との関連は、活性酸素種によって誘発されるミトコンドリア DNA の有害な変異、DNA 損傷、脂質とタンパク質の急速な酸化に起因すると考えられており、これらは老化した組織で一般的に観察されます (Cui et al., 2012)。活性酸素種の蓄積は、パーキンソン病、糖尿病、がんなど、いくつかの加齢に伴う病状にも関連していました (Sanz, 2016)。通常の状況下では、細胞はスーパーオキシドジスムターゼ (SOD)、カタラーゼ (CAT)、グルタチオンペルオキシダーゼなどの活性酸素種代謝酵素 (抗酸化物質) を生成して、活性酸素種の影響を打ち消します (Sergiev et al., 2015)。酸化ストレスと老化の関連性は、Caenorhabditis elegans を使用した研究によってさらに確認されています。ある研究では、酸化ストレスに対する耐性が増加した変異株が寿命を延ばしたことが報告されており、別の研究では、スーパーオキシドジスムターゼ/カタラーゼ模倣薬による治療が生物の寿命を延ばしたことが報告されています (Melov et al., 2000; Cui et al., 2012)。さらに、酸化ストレスのレベルが低いトランスジェニックキイロショウジョウバエの平均最大寿命は、最大 3 分の 1 まで延長され、歩行速度の増加などの加齢に伴う破壊的な変化が減少しました (Sohal and Weindruch, 1996)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.3.エネルギー恒常性の不均衡 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| エネルギー恒常性の不均衡は、老化の最新の提案された原因です。エネルギー恒常性不均衡理論は、エネルギー消費が寿命に反比例することを示唆しています。心拍数の速い小型の哺乳類は、線虫や昆虫よりも多くの酸素を利用するため、寿命が短くなるという自然現象が存在します。カロリー制限 (CR) は、依然として最も強力なアンチエイジング介入の 1 つであり、エネルギー恒常性の不均衡理論を最もよく実証しています (Anderson et al., 2009; Lipsky and King, 2015)。 カロリー制限は、栄養失調を誘発することなくカロリー摂取量を減らすことによって達成されます。サル、ラット、虫、酵母のモデルを含む複数のモデルで、カロリー制限は老化を遅らせ、寿命を延ばすことが示されています (Lipsky and King, 2015)。たとえば、げっ歯類では、55% から 65% の カロリー制限で、平均寿命と最大寿命が最大 65% 増加することを示すのに十分でした (Anton and Leeuwenburgh, 2013)。 カロリー制限は、グルコースとインスリンのレベルを下げることによって作用すると仮定されています。これにより、アデノシン一リン酸活性化プロテインキナーゼ (AMPK) やサーチュインなどのエネルギー感知ネットワーク制御因子が活性化されます (Anderson et al., 2009; Aliper et al., 2017)。カロリー制限とサーチュインは、オートファジー (細胞質成分が分解されて再利用されるプロセス) を上方制御することにより、細胞エネルギーの恒常性を維持します (Ulgherait et al., 2014)。その後、不要な老廃物の除去は細胞の健康を促進し、酸化ストレスを防ぎ、寿命を延ばします (Madeo et al., 2015)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.老化研究で使用されるモデル | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 人間は平均寿命が長いため、老化プロセスを調査することは困難です。 さらに、研究に影響を与える可能性のある食事やライフスタイルなどの要因を制御することは不可能であり、非倫理的です. このように、老化研究を行う上で欠かせないツールとして、哺乳類や他の脊椎動物、真核微生物を用いた様々な老化モデルが開発されています。 モデルが異なれば寿命も異なり、目的も異なります。 したがって、単純な遺伝子研究であろうと、神経老化研究などのより機構的な生物学的研究であろうと、研究者は研究の範囲に応じて適切なモデルを選択することが重要です (Mitchell et al., 2015)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.1.ヒト以外の霊長類 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 加齢研究では、ヒト以外の霊長類とげっ歯類が最も広く使用されている脊椎動物です。ヒト以外の霊長類は、ヒトに最も近い動物であり、解剖学、遺伝的同一性、および生理学の点でヒトに非常に似ています。彼らは、代謝障害、筋骨格の弱体化、生殖能力の低下などの加齢に伴う病気を経験しています。老化研究において他の哺乳類よりもヒト以外の霊長類を使用することの最大の利点は、それらが人間と同様の脳機能と思考スキルを持っているため、神経老化研究の効果的なモデルであることです (Sprott、2011; American Federation for Aging Research、2016) )。類人猿、チンパンジー、マーモセットなどの霊長類の中でも、アカゲザルは人間の 3 倍の速さで老化し、調査期間が大幅に短縮されるため、特に有用です (American Federation for Aging Research, 2016)。アカゲザルに関する最も有名な老化研究の 1 つは、カロリー制限がどのように病気の発症と死亡率を遅らせるかを評価したものです。ウィスコンシン国立霊長類研究センターで実施された 20 年間にわたる縦断的研究で、Colman らは次のように述べています(2009) 。 中等度のカロリー制限が老化に関連する死亡の発生率を改善することを発見しました。通常の食事をしているサルは、15.5 歳の若さでサルコペニア (60 歳以上の人間に一般的に見られる加齢に伴う筋肉量の減少) を発症しました (Colman et al., 2009)。同じ研究から得られたもう 1 つの興味深い発見は、脳の萎縮に関するもので、これは小さな動物では正確に調べることができません。ヒトの加齢に伴い、脳の白質が 1 年に 2 mL の割合で減少することが観察されています (Double et al., 1996)。 Colman et al.の脳萎縮に関する研究 (2009) は、アルツハイマー病の早期発症の検出に価値があります。 カロリー制限は、運動機能を制御する領域の加齢に伴う脳萎縮を軽減することが示されました。人間に非常に似ているにもかかわらず、霊長類を研究に使用することにはいくつかの欠点があります。たとえば、飼育関連の問題や、毎日のライブモニタリングを行うための高度な機器の必要性などです。霊長類は非常に知的な動物であり、潜在的に危険な病気を伝染させる可能性があるため、それらを適切なケアの下に置くことは非常に労力と費用がかかります (Mitchell et al., 2015)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.2.げっ歯類 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 霊長類とは異なり、げっ歯類はより小さな収容施設を必要とするため、扱いが簡単です。寿命が短く繁殖しやすいことから、最も人気のある実験動物にもなっています (実験動物の 80% はマウスとラットです)。げっ歯類、特に実験用ラットの平均余命は 3 年で、これは人間の 90 歳に相当し、2 歳のラットは 60 歳の人間に匹敵します。これらの相関データが利用できるため、加齢研究の実験を設計することができます (Andreollo et al., 2012)。近交系マウスは、その遺伝的均一性から、加齢や加齢に伴う疾患の研究に広く使用されています。近親交配によって、同じ遺伝子系統の動物間の差異が少なくなります。それらの遺伝学、生理学、および行動に関する広範なデータは、2001 年に Jackson Laboratories によって開始されたマウス フェノーム データベース (MPD) でも利用できます。機構的および細胞老化研究の実施に役立ちます (Mitchell et al., 2015)。 さらに、1970 年頃に日本で開発された有名な老化促進マウス (SAM) 系統 (Takeda et al., 1991) などの変異マウス系統も容易に入手できます。 現在、老化促進マウス (SAM-P)/1、-P/2、-P/3、-P/6、-P/7、-P/8、-P/9、および -P/10 および老化抵抗マウス(SAM-R)/1 および -R/2の全部で12の変異株があります。 老化促進マウスおよび老化抵抗マウスは、系統固有の表現型を示します。たとえば、SAM-P/1、-P/2、および -P/7 における老人性アミロイドーシス、 SAM-P/3 における変性関節疾患、 SAM-P/8 における学習および記憶障害などです (Takeda et al., 1991)。特定の病状のモデルとして機能するように開発された他の遺伝子改変マウスも表 1 にリストされています (Kuro-o et al., 1997; Blüher et al., 2003; Yan et al., 2007; Yuan et al., 2011; Mitchell et al., 2015)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| げっ歯類では、D-ガラクトース (D-gal) などの化学物質を使用して老化を加速させることができます。マウスとラットを使用したさまざまな研究では、D-ガラクトースの慢性的な低用量投与は、酸化ストレスレベルと最終糖化産物の量を増加させ、最終的に加齢に似た症状を引き起こします。たとえば、D-ガラクトースを使用して老化させたげっ歯類は、神経障害、神経筋活動の低下、および免疫系の障害を示すことが判明しました (Song et al., 1999)。老化に関連する遺伝的および生化学的変化だけでなく、認識に対するD-ガラクトースの破壊的な影響も、ステップスルー法や自発的な運動活動の評価などの行動ツールを使用してげっ歯類で評価できます。ケージは、動きを検出して記録するために使用されます。慢性的なD-ガラクトース治療を受けたマウスは、自然に老化した対照マウスと比較して、自発的な運動活動が低く、潜伏期間が長く、ステップスルー法でのエラー率が高かった (Song et al., 1999)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.3.ショウジョウバエ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ショウジョウバエ(fruit fly)としても知られるキイロショウジョウバエ(D. melanogaster)は、老化および長寿研究のモデルとして広く使用されています (2,000 件を超える出版物) (Hoxha、2012)。寿命が短く (平均寿命は 2 ~ 3 か月)、メンテナンスが容易で、遺伝的に扱いやすいため、魅力的な遺伝モデルです (Sun et al., 2013; He and Jasper, 2014; Heintz and Mair, 2014)。その上、キイロショウジョウバエは、人間の加齢に伴う生理学的、行動的、および解剖学的変化を調査するための関連モデルです (Iliadi et al., 2012)。一般に、キイロショウジョウバエの研究で一般的に採用されている基本的なアプローチには、寿命、食事組成、食物摂取量、生涯生殖出力、生理的および行動的変化の分析が含まれます (Sun et al., 2013)。キイロショウジョウバエの加齢に伴う生理的および行動的変化を評価するには、高速反復負走性 (RING) アッセイなどの自発運動アッセイが信頼できます (Iliadi et al., 2012; Sun et al., 2013)。高速反復負走性アッセイは、機械的刺激に対するクライミング速度、自発運動、逃避反射を測定します。 キイロショウジョウバエでは、登る速度が年齢に依存することが実証されています (Iliadi et al., 2012; Sun et al., 2013)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
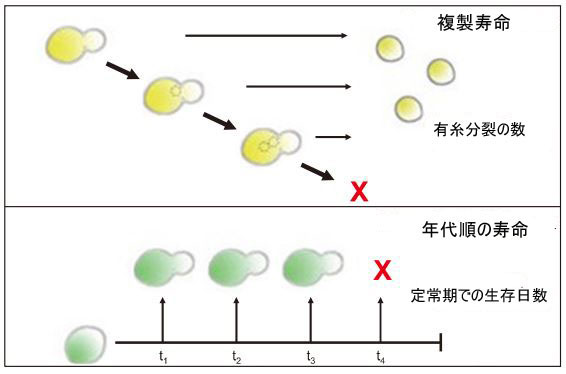
| さらに、カロリー制限はキイロショウジョウバエで十分に確立されており (Lee and Min, 2013)、老化のプロセスと加齢関連疾患の発症を遅らせることに成功しています (Taormina and Mirisola, 2014; López-Lluch and Navas, 2016)。 キイロショウジョウバエでは、カロリー制限は通常、栄養素、通常はタンパク質と炭水化物の比率を希釈することによって達成されます (Lee and Min, 2013; Taormina and Mirisola, 2014)。さらに、キイロショウジョウバエの遺伝子の 50% 以上がヒトの遺伝子と相同であり、ヒトの疾患に関与する遺伝子の 75% 以上がハエの相同体を持っています (Hoxha, 2012; Sun et al., 2013)。 P エレメント挿入による突然変異誘発、Gal4-無人航空機システム (UAS) および GeneSwitch Gal4-UAS による遺伝子発現、RNA 干渉による遺伝子ノックダウンなどの技術は、キイロショウジョウバエを使用した老化研究で一般的に使用され、寿命における単一遺伝子の関与を決定します。変調または疾患の発症 (Sun et al., 2013)。さらに、キイロショウジョウバエの目は、ハンチントン病、アルツハイマー病、およびパーキンソン病を調査するための神経変性疾患モデルとして広く使用されています (Sang and Jackson, 2005)。成虫のハエの眼の表現型は、基本的な生物学的プロセスの遺伝的破壊に耐性があるため、容易に検出されます (Sang and Jackson, 2005)。 4.4.単細胞真核生物 このレビューで説明する最後の老化モデルは、真核生物の出芽酵母Saccharomyces cerevisiae です。 老化研究における酵母の使用は、モーティマーとジョンストンが酵母細胞の有限の複製能力について最初に発表した 60 年前にさかのぼることができます(Mortimer and Johnston, 1959)。 酵母の老化を研究するには、複製寿命 (RLS) と年代的寿命 (CLS) の 2 つのアプローチがあります (図 1)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 図1 酵母老化モデルの複製寿命と年代順寿命の模式図 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 複製寿命は老化に達する前に母細胞によって生成される娘細胞の数を指し、年代的寿命は酵母細胞が非分裂状態で生存する期間として定義されます (Kaeberlein et al., 2007)。研究された最も単純なモデルであるため、酵母の老化は、サーチュインとラパマイシンの標的 (TOR) シグナル伝達経路という 2 つの主要な経路のみによって影響を受けます。これらの発見により、老化のメカニズムを理解し、他のモデル生物におけるアンチエイジング治療介入を発見することができました (Longo et al., 2012)。酵母の長寿プロモーターとしてのサーチュインの役割は、サーチュインファミリーのメンバーであるSIR2の欠失によって引き起こされる複製寿命短縮によって実証されました。逆に、SIR2 の過剰発現は複製寿命の増加につながりました。 TOR シグナル伝達経路は複製寿命と年代的寿命の両方を調節し、TOR 活性を阻害すると長寿が促進されます。通常の増殖条件下では、酵母は高グルコース (2%) と十分なアミノ酸濃度を添加した培地で維持されます。いくつかの研究では、グルコースまたはアミノ酸のいずれかの濃度を下げると複製寿命と年代的寿命が改善されることが報告されています (Kaeberlein et al., 2007)。老化モデルとして酵母を使用することの欠点は、複製寿命を拡張する酵母遺伝子のホモログが、他の複雑な多細胞真核生物では同様の効果を示さない可能性があることです。それにもかかわらず、他の動物モデルよりも酵母の in vitro 分析の変数を調整および監視する方が簡単です。酵母は、最も費用対効果の高いモデルでもあります。したがって、少なくともいくつかの重要な保存された遺伝子の同定のために、老化の仮説を検証するのに役立ちます. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.プロバイオティクス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内微生物叢と中枢神経系との複雑な関係は、機能性食品や栄養補助食品をプロバイオティクスとして使用して、脳の健康を高めることを期待して腸内微生物叢を調節することに大きな関心を集めています (Liu et al., 2018)。プロバイオティクスの定義は、過去数十年にわたって何度か改訂されてきました。現在、一般的に受け入れられている定義は、「適切な量を投与すると、宿主に健康上の利益をもたらす生きた微生物」です (国連食糧農業機関、世界保健機関、2006 年)。最も広く研究されているプロバイオティクスである乳酸菌 (LAB) は、非運動性および非胞子形成性のグラム陽性菌のグループです。乳酸菌ファミリーのメンバーには、Lactobacillus, Lactococcus, Enterococcus, Leuconostoc, Pediococcus, および Streptococcus.が含まれます。それらは炭水化物を発酵させ、主な最終産物として乳酸を生成します (Nair and Suredran, 2005)。主にグラム陽性、非胞子形成性、非運動性、およびカタラーゼ陰性の嫌気性細菌であるbifidobacteria(ビフィズス菌)のプロバイオティクス効果も研究されています (Tham et al., 2012)。 過去 20 年間で、さまざまな疾患における治療特性により、プロバイオティクスの使用が増加しました。 プロバイオティクスが下痢、痛み、便秘、および炎症の発生率を低下させる腸関連の病気から (Hor et al., 2018)、高血圧 (Yeo et al., 2009; Fung and Liong, 2010)、高コレステロール血症 (Lye et al., 2017)、などの代謝性疾患および呼吸器の健康 (Hor et al., 2018) を改善します。 しかし、腸-脳軸という新たな概念により、プロバイオティクスの健康上の利点は、これらの従来の利点を超えてさらに拡大しました (Liu et al., 2018)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.腸-脳軸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
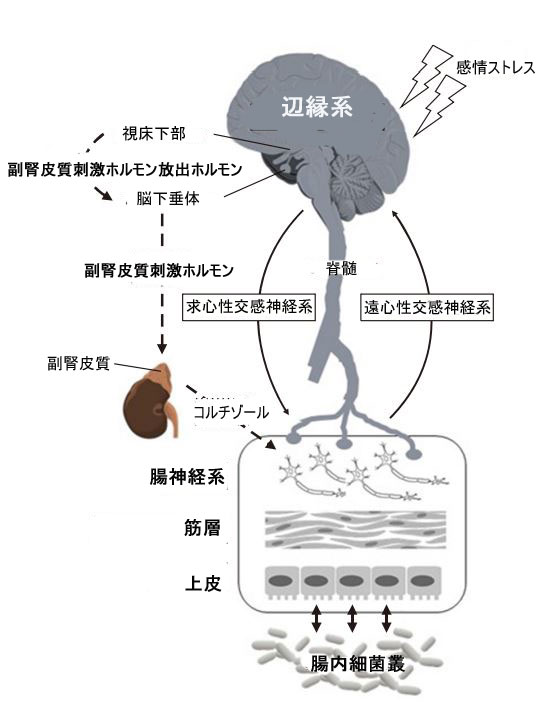
| 腸-脳軸は、中枢神経系、交感神経系、腸神経系、視床下部-下垂体-副腎 (HPA) 軸、および腸内微生物叢からなる双方向のコミュニケーション ネットワークです (図 2)。これは比較的新しい概念であり、腸の健康と脳の健康との関連性と影響を説明するために科学界によって最近採用されました (Arck et al., 2010)。脳と腸の間の双方向通信は、19 世紀初頭の研究によってすでに認識されていました。代表的な例の 1 つは、1833 年に患者の胃の瘻孔を通して胃分泌物を監視することにより、気分と腸の機能との関連性を指摘したボーモントの研究です。彼の調査結果は、個人の感情状態が胃腸 (GI) 管の機能に影響を与え、逆もまた同様であることを示しました (Beaumont, 1977)。腸内微生物叢は、ヒトの消化管全体に分布しており、宿主にとって重要な代謝および生理学的機能を果たしています。腸-脳軸の双方向性により、拘束条件や食物不足などのさまざまな環境ストレッサーが、宿主の腸内微生物叢に望ましくない変化を引き起こす可能性があります。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 図2 腸-脳軸構造 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 視床下部-下垂体-副腎 (HPA) 軸 (破線) は、感情やストレスに反応して活性化されます。 視床下部からの副腎皮質刺激ホルモン放出ホルモン (CRH) の分泌は、下垂体からの副腎皮質刺激ホルモン (ACTH) の分泌を刺激します。視床下部-下垂体-副腎軸は、副腎からのコルチゾールの放出で終わります。 並行して、中枢神経系は求心性交感神経系と遠心性交感神経系 (SNS) の両方を介して、腸神経系 (ENS)、腸の筋肉層、腸上皮などのさまざまな腸の標的と通信します。 腸内微生物叢とこれらの腸の標的との間に双方向のコミュニケーションがあり、腸の運動性、免疫、透過性、粘液の分泌などの胃腸機能を調節します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7.腸内微生物叢と腸-脳軸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内微生物叢としても知られる腸内の複雑な微生物生態系には、約 100 兆の微生物が含まれています。それは腸内層を確立する機能を果たし、その維持に貢献します (Mangiola et al., 2016)。腸内微生物叢は何千もの異なる微生物種で構成されていますが、最も優勢な微生物は Firmicutes 門と Bacteriodetes 門に属しています (Sartor, 2008)。腸にコロニーを形成する微生物は、哺乳類の健康を維持するために不可欠であり (Jarchum and Pamer, 2011)、腸細胞および腸神経系と局所的に相互作用することにより、宿主と共生関係にあることが報告されています (Hooper and Macpherson, 2010)。神経内分泌および代謝経路を介して中枢神経系に直接伝達されます (Carabotti et al., 2015)。まだ初期段階ですが、臨床的および実験的調査により、人間の脳の発達、行動、および気分における腸内微生物叢の重要な役割が示されています (Tillisch et al., 2013; Mayer et al., 2014)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 加齢は、腸の機能と組成の変化と密接に関連しています。実際、高齢者では腸内細菌叢の多様性の低下と加齢に伴う疾患の発生率の増加が報告されています (Hor et al., 2019)。 D-gal を使用してラットの老化を誘導したところ、Hor et al (2019) は、Firmicutes/Bacteroidetesの比率が老化中に大幅に低下することを発見しました。しかし、この効果は、Lactobacillus helveticus OFS 1515 や Lactobacillus fermentum DR9 などのプロバイオティクスの投与によって防止されました (Lew et al., 2020)。さらに、Lactobacillus paracasei OFS 0291 および L. helveticus OFS 1515 は、日和見病原体である Bacteroides の量を減少させますが、L. fermentum DR9 投与は Lactobacillus の増殖を促進し、その結果、D-gal 処理ラットの糞便酢酸レベルを増加させます。老化はしばしば大きな代謝変化を引き起こします。プロバイオティクスはこれらの障害を改善しますが、変化の大きさと特異性はしばしば菌株に依存します (Lew et al., 2020)。 L. paracasei OFS 0291 および L. helveticus OFS 1515 は、アラビノースやリボースなどの糖レベルを若いラットに存在する量と同様の量に戻します。Lactobacillus plantarum DR7、Lactobacillus reuteri 8513d は、トリプトファン、ロイシン、チロシン、システイン、メチオニン、バリン、リジンなどのアミノ酸の糞便含有量を増加させました。 一方、L. fermentum DR9 の投与により、エリスリトール、キシリトール、アラビトールなどの炭水化物代謝に関連する化合物の有病率が高くなりました。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.神経化学レベルの変化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸の感覚運動機能の変化に加えて、微生物のコロニー形成の欠如は、腸と中枢神経系の両方における神経伝達物質の異常と関連しています (Diaz Heijtz et al., 2011; Asano et al., 2012; Clarke et al., 2013)。ニューロトロフィン ファミリーのメンバーである脳由来神経栄養因子 (BDNF) は、学習および記憶機能における極めて重要な役割のため、特に興味深いものです。脳由来神経栄養因子の発現が老化中に減少することは十分に確立されています (Chapman et al., 2012)。興味深いことに、Bifidobacterium longumNCC3001 などのプロバイオティクスは、海馬の脳由来神経栄養因子mRNA 発現を変化させることによって有益な効果をもたらします (Bercik et al., 2011)。プロバイオティクスと腸-脳軸との関係を証明するために、迷走神経切断マウスに B. longum NCC3001 を与えました。驚いたことに、B. longum NCC3001 は迷走神経切断マウスでは治療効果がなく、プロバイオティクスと腸-脳軸との関係についての洞察が得られました。他の多くの研究は、おそらく迷走神経を介して、プロバイオティクスと消化管内の受容体との直接的な相互作用を示しています。Lactobacilliによって産生される神経活性化合物、特に神経伝達物質の産生に関与する化合物に関する証拠が増えています。中枢神経系における神経伝達物質シグナル伝達の調節におけるプロバイオティクスの効果には、直接的および間接的なものがあります (Wang et al., 2016)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 最近、Liu et al (2020) は、L. plantarum DR7 を 12 週間投与された成人は、プラセボ群の成人と比較してストレスと不安が軽減されたことを報告しました。 L. plantarum DR7 の投与は、セロトニン、ドーパミン、ノルエピネフリンなどの脳神経伝達物質経路の変化も誘発しました (Liu et al., 2020)。 12 週間の投与中、DR7 グループの腸内細菌叢のアルファとベータの多様性は、クラスおよび順序レベルでプラセボ グループの多様性とは異なっていました。 特定の細菌グループの違いが特定され、Bacteroidetes門とFirmicutes門、およびDelta-proteobacteriaとActinobacteriaのクラスを含むさまざまな分類レベルで一貫性が示されました。 プラセボ群で存在量が減少したBacteroidetes、Bacteroidia、および Bacteroidales目は、ドーパミン経路におけるドーパミンベータヒドロラーゼ(DBH)遺伝子発現と負の相関がありましたが、Bacteroidia と Bacteroidales目はセロトニン経路でトリプトファンヒドロキシラーゼ-II(TPH2)遺伝子発現と正の相関がありました。 DBH と、プラセボ群で存在量が増加した Firmicutes、Clostridia、Clostridiales、Blautia、および Romboutsia との間に相関関係が観察されました。 セロトニン経路に関して、Blautiaはトリプトファン 2,3-ジオキシゲナーゼと関連していましたが、Romboutsia は TPH2 と負の相関がありました。 プラセボ群で存在量が減少したDeltaproteobacteria とDesulfovibrionalesは、DBHと負の相関がありましたが、BilophilaはTPH2と関連していました。 これは、プロバイオティクスによって誘発される生理学的変化だけでなく、腸内細菌叢と、脳のセロトニン作動性およびドーパミン作動性経路に沿った分類群特異的な変化との関連も示した、腸-脳軸の先駆的な研究でした。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9.免疫調節反応の変化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| インターフェロン-γ (IFN-γ) やインターロイキン (IL) などの炎症誘発性サイトカインの発現増加は、加齢や加齢に伴う疾患に関与しています (Michaud et al., 2013)。炎症の増加は、最終的に視床下部-下垂体-副腎軸の過剰活性化につながり、神経伝達物質の活動と代謝を変化させます。多数のサイトカイン受容体が末梢神経に位置しており、神経炎症の開始とその後の認知機能低下の原因への関与を示唆しています (Goehler, 2008; Luo et al., 2014)。プロバイオティクスは、ホストの胃腸の完全性を改善し、腸の炎症を減少させることにより、視床下部-下垂体-副腎軸と神経伝達物質の活性の調節を間接的に改善することが提案されています。プロバイオティクスを与えられた早期ストレス (ELS) ラットのフォース スイミング テストでの成績が向上し、炎症性サイトカイン IL-6 の濃度が低下しました (Desbonnet et al., 2010)。コントロール ラットと比較して、プロバイオティクスを与えられた早期ストレスラットでは両方のパラメーターが正常化され、脳の健康の改善におけるプロバイオティクスと抗炎症反応との密接な関係が示唆されました。さらに、Luo et al (2014) は、L. helveticus NS8 の投与が高アンモニア血症による神経炎症を防ぎ、その後ラットの認知障害を軽減することを実証しました。これらの研究は、プロバイオティクスが免疫調節経路を介して脳の健康を間接的に調節することを示唆しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hor et al (2019) は、加齢が化学的に (D-ガラクトースによって) 誘発されたラットを使用して、加齢が結腸の免疫学的変化を引き起こすことを示しました。 これらの変化は、 L. paracasei OFS 0291, L. helveticus OFS 1515, およびL. fermentum DR9などのプロバイオティクス株によって、通常は免疫調節特性を介して首尾よく逆転されました。 D-ガラクトース処置ラットでは結腸の炎症誘発性サイトカインである腫瘍壊死因子-α (TNF-α)、IFN-γ、IL-1β、および IL-4 のレベルを低下させますが、自然に老化したコントロールでは低下させません。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| プロバイオティクス L. plantarum DR7 は、成人のストレスと不安を軽減します。 また、成人の IFN-γ や TNF-α などの血漿炎症誘発性サイトカインを抑制し、IL-4 や IL-10 などの抗炎症性サイトカインのレベルを高めます。L. plantarum DR7の補給は、プラセボと比較して血漿過酸化および酸化ストレスレベルの低下も伴いました(Chong et al., 2019)。 これらの抗炎症効果も L. plantarum DR7 に起因するものであり、血漿分化クラスター (CD)44、CD117、CD4、および CD8 のより高い発現が見られたプラセボ群で必要とされるT 細胞の活性化をそれほど必要とせずに非休息および成熟したナチュラル キラー細胞の活性を高めました (Chong et al., 2019)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.加齢に伴う生理的変化とプロバイオティクスの役割 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 老化の間に、さまざまな生理学的パラメーターが影響を受け、脳機能の変化につながります。 このセクションでは、マイクロバイオーム、炎症、酸化ストレスを含む加齢に伴う変化について説明します。 これらの変化が脳機能に及ぼす影響と、加齢に伴う生理学的影響の緩和および脳機能の改善におけるプロバイオティクスの役割も提示されています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.1.腸内細菌叢 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 老化はさまざまな生理学的パラメーターに影響を与え、それが脳機能に影響を与えます。腸内細菌叢の組成の変化は、脳機能に影響を与える要因の 1 つとして示唆されています。消化管での微生物のコロニー形成は、出生時に始まり、発達を続け、生後 1 年間で劇的な変化を遂げます (Bäckhed et al., 2015)。最初の低い多様性と複雑さから、腸内細菌叢は多様で複雑な集団を含むように進化します。老化は、おそらく高齢者の腸の運動性の低下が原因で、腸内毒素症に関連していると報告されることがよくあります。これは消化に悪影響を及ぼし、腸内細菌叢を変化させる可能性があります. Hopkins と Macfarlane (2002) は、 Bacteroides の多様性は健康な高齢者の糞便で増加するのに対し、Bifidobacterium の多様性は年齢とともに減少すると報告しました。 Claesson et al (2011) は、高齢者の中核となる微生物叢は若い成人のものとは異なり、高齢者ではBacteroides spp. の割合が高く、Clostridium 群の存在量パターンが異なることを報告しました。 Bacteroides の数が減少したのに対し、 Escherichia coli と Enterococci spp.は高齢者で増加したことも示されました (Enck et al., 2009)。 さらに、Ruminococcus 属のグループが高齢者で最も優勢なグループであり、総微生物叢の約 9.6% に寄与していることが発見されました。 一般に、高齢者の腸内細菌叢は、細菌の多様性の減少、優勢な種の変化、有益な微生物の減少、および通性嫌気性細菌の増加によって特徴付けられます (Salazar et al., 2017)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| プロバイオティクスが定着し、腸内細菌叢の組成を変化させる能力は、一般的に報告されています (Zhang et al., 2014; Kato-Kataoka et al., 2016; Zhang et al., 2018)。脳機能の調節における腸内細菌叢の重要性を考慮して、腸の生態系を調節し、ひいては脳機能を改善するためのプロバイオティクスの使用がますます研究されています。腸内細菌叢組成のプロバイオティクス媒介調節の証拠は、いくつかの研究によって提供されています (Oh et al., 2016; Harata et al., 2017; Xue et al., 2017)。腸内細菌叢のプロバイオティクスを介した調節の根底にあるいくつかのメカニズムも提案されています。たとえば、プロバイオティクスは抗菌活性を発揮したり、粘膜結合部位を他の微生物と競合したりして、最終的に腸内病原体の増殖を抑制する可能性があります (Collado et al., 2007; Spinler et al., 2008)。プロバイオティクスは、腸の免疫とバリア機能を調節することも報告されており、腸内の微生物に対する腸上皮と免疫細胞の応答性の変化につながる可能性があります (Hemarajata and Versalovic, 2013)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.2.炎症 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 老化プロセスのもう 1 つの主な特徴は、「炎症」と呼ばれる炎症誘発性状態の慢性的な進行性増加です (Franceschi et al.,2000)。増加する証拠は、末梢炎症と神経炎症の両方を、高齢者集団における認知機能の低下や神経変性などの脳機能の障害に関連付けています。炎症のマーカーと、アルツハイマー病、パーキンソン病、および軽度認知障害との間の相関関係が観察されています (Simen et al., 2011)。炎症性サイトカインと認知機能との関連性は、ベースラインで 15,828 人の参加者を含むメタアナリシス研究によって裏付けられました。循環 IL-6 レベルが高い被験者は、循環 IL-6 レベルが低い被験者と比較して、2 ~ 7 年の追跡期間中に全体的な認知機能低下を経験する可能性が 1.42 倍高かった (オッズ比: 1.42、95% 信頼区間: 1.18~1.70; P<0.001) (Bradburn et al., 2018)。大うつ病性障害の高齢患者を対象とした症例対照研究でも、IL-6 の血清レベルがうつ病および認知機能低下の臨床的特徴と関連していることが示されました (Ali et al., 2018)。さらに、炎症誘発性サイトカイン IL-17A および TNF-α のレベルの上昇と IL-10 のレベルの低下は、多発性硬化症患者で観察される認知障害と関連していました (Trenova et al., 2018)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| プロバイオティクスは、加齢に伴う炎症の特徴の一部を改善する可能性のある潜在的な免疫系アジュバントとして提案されているため、高齢者の脳認知機能を改善する潜在的な治療薬として機能します。プロバイオティクスが免疫調節活性を発揮する能力は、重度の敗血症の子供 100 人を対象としたランダム化二重盲検プラセボ対照試験で示されました。 プロバイオティクスは7日間子供たちに投与した結果、炎症誘発性サイトカイン (IL-6、IL-12p70、IL-17、および TNF-α) レベルが大幅に低下し、抗炎症性サイトカイン (IL-10 およびトランスフォーミング増殖因子-β1) レベルが上昇しました (Angurana ら、2018)。 これは、最近のメタアナリシスによってさらに裏付けられており、プロバイオティクスの介入により、慢性的な軽度の炎症を伴う中年および高齢者の IL-6 および C 反応性タンパク質のレベルが低下することが示されています (Custodero et al., 2018)。プロバイオティクスは、腸のバリアの完全性を改善し、腸内のタイトジャンクションタンパク質を回復することにより、免疫調節活性を発揮すると仮定されました (Zaylaa et al., 2018)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.3.酸化ストレス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 老化における酸化ストレスの役割は、1956 年に最初に提唱され、現在、老化が生化学的および分子レベルでどのように発生するかについて最も確立された説明の 1 つです (Bokov et al., 2004)。酸化ストレスは、酸化還元の不均衡に起因する損傷として定義され、活性酸素種などの破壊的なフリーラジカルが増加し、抗酸化物質が減少します (Birch-Machin and Bowman, 2016)。活性酸素種はミトコンドリア呼吸の副産物です。したがって、ミトコンドリアは酸化的損傷の主要な標的であることが示唆されています (Cui et al., 2012)。研究は現在、ミトコンドリアの機能不全と、脂質、DNA、およびタンパク質の酸化的損傷の増加を示しており、それらを代謝疾患や脳機能障害などのさまざまな加齢に伴う疾患と相関させています。酸化ストレスは、アルツハイマー病の発症に寄与する脳内のアミロイドベータ形成を誘発することが報告されています。酸化ストレスは、うつ病、アルツハイマー病、統合失調症などの神経精神疾患の病態生理学における重要な要因である神経炎症を引き起こすことがよくあります (Popa-Wagner et al., 2013)。高レベルの活性酸素種は、パーキンソン病やアルツハイマー病などのさまざまな神経疾患における神経細胞死とも頻繁に相関しています (Guglielmotto et al., 2009)。酸化ストレスは最終的に高齢者集団の認知障害の割合を増加させることを考慮して、研究は現在、抗酸化防御の改善に焦点を当てています。たとえば、多価不飽和脂肪酸、ドコサヘキサエン酸、または n-3 脂肪酸を投与すると、活性酸素種の蓄積が減少し、神経保護が強化されます (Kang and Gleason, 2013; Tan et al., 2016)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| プロバイオティクスの抗酸化活性も多くの研究で報告されています。 たとえば、Lactobacillus acidophilusを含むプロバイオティクス ヨーグルトを 9 週間摂取すると、酸化ストレス マーカーのレベルが低下しました (Mikelsaar and Zilmer, 2009)。 無作為二重盲検プラセボ対照臨床試験では、2 型糖尿病患者の血漿総抗酸化能や総グルタチオン (GSH) などの酸化ストレス バイオマーカーのレベルが、複数種のプロバイオティクスの投与によって改善されることが示されました (Asemi et al.ら、2013)。高脂肪食を与えられたラットへの L. plantarum P-8 の投与は、肝臓脂質の蓄積の減少と肝機能の保護によって反映されるように、高脂肪食によって誘発される酸化ストレスに対して有益な効果をもたらします (Bao et al., 2012 )。 プロバイオティクスの抗酸化活性のいくつかのメカニズムも提案されています。 Wang et al (2017) によって議論およびレビューされたように、プロバイオティクスは金属イオンをキレート化し、金属イオンが酸化を触媒するのを防ぐことによって抗酸化活性を発揮します。 プロバイオティクスはまた、スーパーオキシドイオンの過酸化水素と水への分解を触媒するスーパーオキシドジスムターゼなどの抗酸化酵素の活性を高め、したがって活性酸素種レベルの中心的な調節因子です (Landis and Tower, 2005)。 プロバイオティクスは、総グルタチオンや葉酸などの抗酸化代謝産物も生成し (Kullisaar et al., 2002)、抗酸化活性に関連するいくつかの宿主経路を刺激します (Wang et al., 2017)。 最後に、プロバイオティクスは腸内細菌叢の組成を変えることで抗酸化の役割を果たし、酸化ストレスの一因となる有害な細菌の過剰な増殖を防ぐ可能性があります (Wang et al., 2017)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 11.サイコバイオティクス | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 近年、脳の健康に有益な影響を与えるために、腸内細菌叢を標的とする研究への関心が高まっています。有望な戦略は、サイコバイオティクスの使用です。サイコバイオティクスは有益な細菌、特にプロバイオティクス、またはこれらの細菌のサポート (プレバイオティクス) であり、十分な量を摂取すると細菌と脳の関係に影響を与えます (Dinan et al., 2013)。 サイコバイオティクスの治療効果は、多くの場合、次のように分類されます。
(1) ストレス応答に対する全身的な影響、特に視床下部-下垂体-副腎軸に対する影響。 (2) 神経伝達物質と神経タンパク質の生理学的変化。 (3) 感情および認知機能の変化を含む心理的変化 (Sarkar et al., 2016)。 げっ歯類で観察されたプロバイオティクスの向精神特性を人間に翻訳することは、驚くほど有望です。過去 10 年間に、高齢者による推定サイコバイオティクスとしてのプロバイオティクスの使用に関する多数の臨床研究が実施されました (表 2) (Rao et al., 2009; Messaoudi et al., 2011; Tillisch et al., 2013; Chung et al., 2014; Akbari et al., 2016; Akkasheh et al., 2016)。 たとえば、L. helveticus IDCC3801 発酵乳で治療された健康な高齢者では、認知疲労が改善されました (Chung et al., 2014)。別の最近の研究では、アルツハイマー病患者の認知機能を改善するプロバイオティクスの能力が示されました (Akbari et al., 2016)。プロバイオティクス ミルクを 12 週間という短期間提供された患者は、Mini-Mental State Examination スコアによって評価されるように、認知機能の顕著な 35% の改善を示しました。 これらの研究は、特に老化時の認知に対するプロバイオティクスの有望な治療効果の証拠を提供します。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 12.結論 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 栄養補助食品や健康食品に対する消費者の需要の高まりは、脳の健康と腸-脳軸との関連性に関する新しい視点の開発を含む、プロバイオティクスの分野で多くの革新と開発を促しました。 さらに重要なことに、脳の健康状態の低下は老化にも関連していることが多く、健康な脳は高齢者が自立して生活できることを保証します。 ここでは、健康的な老化と脳と腸の健康維持におけるプロバイオティクスの役割に関する重要な新しい発見を紹介します。 さらに、自然なアンチエイジングの食事戦略としてのプロバイオティクスの可能性を実証しました。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| この文献は、Prev Nutr Food Sci. 2022 Mar 31; 27(1): 1–13.に掲載されたProbiotics: The Next Dietary Strategy against Brain Aging.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |