Anjana and Santosh Kumar Tiwari |
概要 |
| 乳酸菌のいくつかの株は強力なプロバイオティクスであり、さまざまな作用機序を使用してさまざまな病気を治すことができます。 これらの細菌は抗菌ペプチド、バクテリオシンを産生し、これは一般的に密接に関連する細菌株や、Listeria, Clostridium, および Salmonellaなどの他の病原性細菌を阻害または殺します。 バクテリオシンは、細孔形成とサイトゾル内容物の散逸によって標的細胞を殺し、細胞死に導くカチオン性ペプチドです。 バクテリオシンは、宿主のいくつかの健康増進機能に影響を与え、ネイティブ微生物叢と宿主免疫を調節することも知られています。 この総説では、バクテリオシンを産生するプロバイオティクス乳酸菌が腸内細菌叢を調節して腸内毒素症(dysbiosis:腸内細菌叢の異常)を是正し、慢性感染症、炎症性腸疾患、肥満、がんなどのいくつかの重要な人間の障害の治療/維持を行う能力について説明しました。 |
| 目次(クリックして記事にアクセスできます) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.乳酸菌 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.バクテリオシン | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.バクテリオシン産生乳酸菌 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.腸内細菌叢と免疫調節 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.腸内細菌叢と腸-脳軸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7.バクテリオシン産生乳酸菌とその疾患における役割 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.結腸感染症 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9.炎症性腸疾患 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 11.肥満 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1.はじめに | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内微生物叢は、宿主防御システムと恒常性を維持し、病原体から保護し、腸の完全性を強化する上で重要です。 腸内細菌の大部分は、Bacteroidetes (例、 Porphyromonas, Prevotella), Firmicutes (例、 Enterococcus, Lactobacillus, Streptococcus, Ruminococcus, Clostridium), Actinobacteria (例、 Bifidobacteria),およびProteobacteria (例、 Escherichia coli) に属します (Zhang et al. 、2015)。 共生腸内微生物叢の乱れは、不均衡な食事、感染、または抗生物質の使用などのいくつかの要因によって起こる腸内毒素症を引き起こし、腸内共生微生物叢の長期的な変化を引き起こし、多数の致命的な病気を促進する可能性があります (Lange et al., 2016)。 ウイルス感染、炎症性腸疾患 (IBD)、クローン病 (CD)、結腸直腸がん、肥満など、腸内毒素症に関連するいくつかの疾患があります (Kim et al., 2019)。 腸内毒素症は、アレルギーや自己免疫疾患、炎症性疾患などの免疫調節不全に関連する疾患の発症をもたらします (D'amelio and Sassi, 2017)。 バクテリオシンを産生するプロバイオティクス株による腸のコロニー形成は、競合、ニッチの除去、および空間的分離を通じて、病原体の腸上皮細胞への接着を阻害します (Heilbronner et al., 2021)。 プロバイオティクスは生きた微生物であり、適切な量を摂取すると、腸内細菌叢を改善することで宿主に健康上の利益をもたらします (Binda et al., 2020)。 バクテリオシンは、これらの細菌によって産生される抗菌ペプチドであり、一般に腸内の病原菌を阻害/殺菌し、マウス、ブタ、ニワトリなどの動物モデルの腸内微生物叢の組成を変化させます (Gillor et al., 2008; Yang et al., 2014 )。 バクテリオシンの作用機序は、孔形成と膜破壊によって標的細胞を殺す抗生物質とは異なります。 さらに、リボソームで合成されたタンパク質であるバクテリオシンは、タンパク質分解酵素によって分解されるため、病原体は腸内で耐性を発達させることができません (Epand et al., 2016; Umu et al., 2017)。 さらに、バクテリオシンは、従来の抗生物質に比べて生合成メカニズムが単純であり、生物工学の助けを借りて標的微生物に対する活性を容易に高めることができます。 さらに、多剤耐性病原体に対するより高い比活性は、治療への応用に利点を提供します (Perez et al., 2014)。 したがって、バクテリオシンおよび/またはバクテリオシン産生プロバイオティクスの使用は、腸管感染症を含むいくつかの疾患の治療および健康促進微生物群集の回復のための新しいアプローチです (Fong et al., 2020)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.乳酸菌 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| プロバイオティクス乳酸菌 (LAB) は、カタラーゼ陰性、グラム陽性、非胞子形成性細菌の非病原性異種グループです。 それらは、グルコースと、バクテリオシン、バクテリオシン様阻害物質(BLIS)、過酸化水素、ジアセチル、および二酸化炭素などのいくつかの成長阻害物質から主な生成物として乳酸を生成します. これらの細菌は、成長のためにアミノ酸、ペプチド、ヌクレオチド塩基、ビタミン、脂肪酸、炭水化物などの複雑な栄養物質を必要とします (Mokoena, 2017)。 それらは、乳製品、発酵肉、魚、飲料、漬物、穀物、および人間や動物の空洞に含まれています. 重要な属には、 Lactococcus, Enterococcus, Streptococcus, Pediococcus, Aerococcus, Alliococcus, Carnobacterium, Dolosigranulum, Oenococcus, Tetragenococcus, Vagococcus, Weissella, および最大の属である Lactobacillusが含まれます (Bintsis、2018 年)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lactobacilli(乳酸桿菌)は、人間や他の動物に見られる最も一般的なプロバイオティクスです。 腸 内で見つかったlactobacilliの主な種は、L. gasseri, L. crispatus, Limnosilactobacillus reuteri, Ligilactobacillus salivarius, および L. ruminis です (Walter, 2008; Zheng et al., 2020)。 メタゲノム解析は、異なる種のBifidobacteriumとLactobacillusの組み合わせを使用した治療が、マウスの腸内微生物叢の組成を著しく変化させることを示唆しています (Azad et al., 2018)。 乳酸菌以外では、Bifidobacterium(ビフィズス菌)は、宿主に健康上の利益をもたらす最初の腸内コロニー形成微生物と見なされています (O'Callaghan and Sinderen, 2016)。 母乳育児中の乳児では、Bifidobacteriumの種が広範囲に存在し、年齢とともに徐々に変化します。 B. longum、B. bifidum、および B. breve は一般に乳児の腸で優勢ですが、B. catenulatum、B. adolescents、および B. longum は成人に存在します。 それらは、さまざまな胃腸 (GI) 疾患の治療/維持のための治療薬として使用され、有害な微生物を制限し、胃腸フェンスを強化し、炎症性サイトカインを阻害します (Xue et al., 2017)。 乳酸菌以外の細菌も腸を支配し、重要な機能を果たします。 たとえば、腸内に見られる無害なE. coli Nissleは、腸内微生物叢のバランスをとるために広く利用されているプロバイオティクスです。 病原性E. coli (大腸菌)による付着と捕捉に対抗して腸のハードルを救うことができるヒト β-ディフェンシン 2 の産生を回復できることが明らかになりました(Schlee et al., 2007; Liu et al., 2017)。 したがって、乳酸菌は腸内微生物叢の主要な構成要素であり、微生物群集全体のバランスを維持する上で重要な役割を果たします。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.バクテリオシン | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| バクテリオシンは、さまざまな細菌によって産生される多機能の抗菌ペプチドであり、低濃度で作用し、一般的に密接に関連する種の成長を阻害します (狭いスペクトル) が、最近の発見は、広いスペクトルのバクテリオシンの存在も示唆しています (Chi and Holo, 2018; Goyal et al. ら、2018)。 バクテリオシン産生細胞は、産生菌の細胞膜に免疫タンパク質が存在するため、これらの抗菌ペプチドに耐性があります。 Lactococcus lactisのいくつかの株によって産生されるバクテリオシンであるナイシンは、米国食品医薬品局 (FDA) によって GRAS (一般に安全とみなされる) ステータスを受けており、一般的に食品の安全性に使用されています (Negash and Tsehai, 2020)。 Zacharof と Lovitt (2012) によると、バクテリオシンは生化学的および遺伝的特徴に基づいて 3 つのクラスに分類されています。 クラス I バクテリオシンは、分子量が 5 kDa 未満のランチビオティックです。 それらは翻訳後修飾され、メチルランチオニンとランチオニンの形成につながります。 クラス II バクテリオシンは、分子量が 10 kDa 未満の非ランチビオティックで、修飾されておらず、熱安定性があり、さらに 3 つのサブクラスに分類されます。 Pediococcus種由来の ペディオシンPA1/AcH などの抗リステリア活性を有するクラス IIa ペプチドには、N 末端保存配列 YGNGVXC が含まれています (Nishei et al., 2012)。 クラス IIb は、L. lactis 由来のラクトコシン G などの 2 つのペプチド バクテリオシンで構成されます。 クラス IIc では、N 末端と C 末端がペプチド結合によって連結され、環状バクテリオシン、たとえばエンテロシン AS-48 を形成します (Van Belkum et al., 2011)。 クラス III は、熱安定性があり分子量が 30 kDa を超える大型のバクテリオシン (エンテロリシンやヘルビチシンなど) で構成されています (Yang et al., 2014)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| バクテリオシンは、いくつかの病原体に対する有効性により、さまざまなヒト感染症に対して有効です。 例えば、Streptococcus pneumonia (肺炎球菌)による肺炎、髄膜炎、敗血症はナイシンで治療できる(Goldstein et al., 1998)。 環状バクテリオシンのグリセリマイシンは、マウスの結核を治すことができました (Kling et al., 2015)。 ペディオシン PA-1 やラクトシン AL705 などのいくつかのバクテリオシンが、抗がん作用と抗炎症作用を示すことが報告されています (Huang et al., 2021)。 例えば、ナイシンは、細胞膜上にイオンチャネルを形成し、乳酸脱水素酵素を放出し、活性酸素種の数を増やし、がん細胞のミトコンドリア呼吸を妨害することにより、がん細胞の増殖を阻害します。 ナイシンは、抗がん剤と組み合わせて、腫瘍のクリアランスにおいて相乗作用を示すことも報告されています (Preet et al., 2015)。 バクテリオシンは、マイトジェン活性化プロテインキナーゼや Toll 様受容体などのさまざまなシグナル伝達経路によって、抗炎症性サイトカインのレベルを上昇させ、炎症誘発性サイトカインのレベルを低下させます (Sassone-Corsi et al., 2016)。 したがって、バクテリオシンは、宿主のさまざまな健康増進活動に使用できる乳酸菌の重要なプロバイオティック代謝産物です。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.バクテリオシン産生乳酸菌 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 動物およびヒトの腸内微生物叢の変化に対するバクテリオシン産生乳酸菌の影響を説明する十分な研究があります (Yang et al., 2014; Hernandez-Gonzalez et al., 2021)。 例えば、ヒト腸の末端回腸から分離された L. salivarius UCC118 によって産生されるバクテリオシン Abp118 は、マウスとブタの腸で抗リステリア活性を示します (Riboult-Bisson et al., 2012)。 L. plantarum P-8 によるヒトの糞便細菌群集の変化は、プランタリシンの産生によるものです (Kwok et al., 2015)。 別の研究では、L. lactis ssp. lactis F10 によって生成されたナイシン F を腹腔内に注射すると、マウスの腸内の細菌群集に対する安定化効果が示されました (Umu et al., 2017)。 別のバクテリオシンである Trnα と Trnβ の 2 つのペプチドから構成されるツリシン クローン病 (CD) は、Bacillus thuringiensis DPC6431 によって分泌され、遠位結腸モデルの共生微生物叢に影響を与えることなく、広範囲の C. difficile分離株を殺します (Rea et al., 2010)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| バクテリオシンを産生する乳酸菌は、ヒトの腸に存在するListeria monocytogenesやいくつかのenterococci (腸球菌)などの食品媒介病原体によって引き起こされる感染症に対して効果的です (Harris et al., 1989; Millete et al., 2008)。 Pediococcus acidilactici UL5 はペディオシンPA-1 を産生し、ネイティブの腸内細菌叢に影響を与えることなく、マウスモデルで抗リステリア活性を示しました (Dabour et al., 2009)。 E. mundtii CRL35 によって産生されたエンテロシン CRL35 を妊娠マウスに投与すると、L. monocytogenes の生体臓器への移行が阻害されました (Salvucci et al., 2012)。 ラットの腸から分離された L. helveticus PJ4 によって生成されたプランタリシン PJ4 は、肥満マウスの体重を減少させる強力な結果を示しました (Bai et al., 2020)。 同様に、L. plantarum NCMIB8826 によって産生されるプランタリシン EF は、食事誘発性肥満マウスで有益な効果を示します (Heeney et al., 2019)。 ナイシン (市販の製剤の形態) を含む食事の免疫調節および免疫刺激効果が評価され、CD4 および CD8 T リンパ球が増加し、B リンパ球細胞数が減少しました (Pablo et al., 1999)。 別の in vivo 研究では、マウスモデルの腸内でのバンコマイシン耐性腸球菌 (VRE) のコロニー形成の減少が、バクテリオシン産生菌である L. lactis MM19 および P. acidilactici MM33 のヒト糞便サンプルから分離されたものを投与することによって報告されました (Millete et al., 2008)。 ラットにS. aureus K(黄色ブドウ球菌K)を投与した後、鼻腔内にナイシン F を投与すると、免疫抑制されたラットは肺炎の症状を示した 一方、S. aureus Kが定着し、ナイシン F で処理されたラットは、健康な気管と肺を示しました (De Kwaadsteniet et al., 2009)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Umu et al (2016) は、腸内細菌叢の調節に対するバクテリオシン産生乳酸菌とその同質遺伝子変異体の効果を実証しました。 この研究で使用されたバクテリオシンは、サカシン A、ペディオシン PA-1、エンテロシン P、Q、および L50 でした。 彼らは、バクテリオシン産生物質の経口投与が全体的な構造を変えないことを実証しました しかし、いくつかの有益な変化は、マウスの腸内のより低い分類学的レベルで発生しますが、いくつかの変化は治療後に元に戻りました. それぞれの株の同質遺伝子変異体がそのような変化を引き起こさなかったことを知ることは興味深いことであり、微生物叢の調節におけるバクテリオシンの役割を示唆しています (Umu et al., 2016)。 Lacticaseibacillus casei、L. acidophilus、L. plantarum、Streptococcus thermophiles などのプロバイオティクスを用量依存的に経口投与すると、Ig-A および Ig-G 産生細胞の数が増加しました (Azad et al., 2018)。 ナイシン Z とペディオシン AcH を投与すると、バンコマイシン耐性Enterococcus に感染する 8 日前に投与すると、病原体のコロニー形成が減少しました (Millette et al., 2008)。 高脂肪食を与えられたマウスに、 L. ramnosus, L. acidophilus, およびB. bifidum,などのプロバイオティクスを組み合わせて投与すると、腸内微生物叢の組成が変化します (Azad et al., 2018)。 したがって、腸内微生物叢の調節と宿主の健康維持におけるバクテリオシン産生プロバイオティクス乳酸菌の役割を示唆する十分な証拠があります。 いくつかの疾患の治療に使用されるバクテリオシン産生乳酸菌を表 1 に示します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.腸内細菌叢と免疫調節 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粘膜免疫系は、消化管を病原体の回避から保護します。 粘膜関連リンパ組織、上皮層、および固有層は、免疫系の主要部分です。 L. fermentum、L. crispatus、および L. gasseri は、ヒト消化管の樹状細胞、腸細胞、および Treg 細胞と相互作用することが知られており、適応免疫が活性化されて、炎症性サイトカインおよび抗炎症性サイトカインが放出されます (Azad et al., 2018)。 プロバイオティクス株の抗原性フラグメントは、腸上皮細胞およびパイエル板の M 細胞に侵入できるため、自然免疫および適応免疫システムに調節があります。 インターロイキン (IL)、腫瘍壊死因子 (TNF)、インターフェロン (IFN) などのサイトカインは、自然免疫系を調節します。 同様に、CD8+ T リンパ球細胞の細胞傷害性 T リンパ球への分化は、ウイルス感染細胞を殺し、ナチュラル キラー細胞とマクロファージを活性化し、病原体を破壊します (Singh and Rao, 2021)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Bifidobacterium breve C-50 の発酵産物が樹状細胞の成熟を引き起こし、抗炎症反応を示す樹状細胞 (DC) と IL-10 の生存を促進することが報告されました。 樹状細胞の長期生存は、抗アポトーシスタンパク質のレベルの上昇によって引き起こされます。 BCL-xl は PBK/Akt のリン酸化を引き起こし、CD86 および CD83 成熟マーカーの効果を高めることで成熟を引き起こします (Hoaru et al., 2006)。 樹状細胞は実行可能な腸内細菌叢を保護し、微生物を「腸間膜リンパ節」に派遣し、粘膜浸潤の反対を防御するIgA抗体の産生をもたらします(Macpherson and Uhr、2004; Macpherson et al。、2005)。 ナイーブ T 細胞から TH-17、TH-2、TH-1、CD8+ リプレッサー、制御性 T 細胞などのさまざまな種類の細胞株への分化は、樹状細胞と特定のパターン認識因子との相互作用に依存します。 ある研究では、サイトファージ-bacteroidesが固有層における TH17 細胞の発生に必要であると述べられており、これにより、制御性 T 細胞集団と TH-17 の間のバランスが維持されます (Foligne et al., 2007; Delcenserie et al., 2008; Zeuthenet al.,2008)。 無菌マウスに Bacteroides fragilis NCTC9343 を定着させると、CD4+ CD45Rb T 細胞集団の数が大幅に回復することが証明されました。 この回復は、B. fragilis 多糖類 A によって引き起こされました (Mazmanian et al., 2008)。 興味深いことに、この多糖類 A を欠く変異株は、CD+ CD45Rb T 細胞集団の数を回復できなかったことが観察されました。 大腸炎のモデルでは、L. paracasei を胃内に投与すると、疾患の重症度を軽減し、進行を遅らせることで保護効果をもたらしました (Mileti et al., 2009)。 樹状細胞をこれらのプロバイオティクスのいずれかで処理すると、ナイーブT細胞がサプレッサー機能を獲得することが観察されました。 Streptococcus thermophilus DN-001 621、Bacteroides adolescentesis DN-150 017、および Bifidobacterium animalis DN173 016.の抑制効果は、分化した T 細胞の増殖および CD4+ エフェクター T 細胞による IFNγ 産生の減少によって引き起こされました (Baba et al., 2008 )。 ナイシンはマウスモデルで免疫調節効果を示し、その結果、CD4+ および CD8 T リンパ球が増加し、B リンパ球が減少し、ナイシンを長期間投与すると、B リンパ球と T リンパ球のレベルのバランスがとれる可能性があります。 ナイシン Z は、ヒト末梢血単核細胞 (PBNC) の炎症誘発性サイトカインのレベルを低下させることにより、自然免疫応答の調節に効果的でした。 好中球の初期バーストがある歯周病に使用でき、その後期段階では、免疫系によって B 細胞および T 細胞関連の免疫応答が示されました (Shin et al., 2016)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.腸内細菌叢と腸-脳軸 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内微生物は一般に、化学伝達物質の中枢神経系 (CNS) パターン (Sanders et al., 2011; Briguglio et al., 2018) に応答して、神経節細胞と相互作用するアミノ酸を分泌します (Dinan and Cryan, 2017)。 腸と中枢神経系の中間における相互作用のさまざまな経路が研究されています (Dinan and Cryan, 2017)。 迷走神経は、腸と脊髄の中間で関係を形成し、脳幹核で終了し、逸脱した繊維に触覚します (Bonaz et al,. 2018)。 したがって、脳幹核は多くの腸の役割に影響を与え、中脳や大脳皮質など、より多くの中枢神経系ゾーンにジェスチャーを伝える可能性があります (Wang and Wang, 2016)。 腸と中枢神経系の中間での交換は、全身の血流を介して行われることもあります (Gibson and Mehler, 2019)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 微生物叢 - 腸 - 脳軸は、さまざまな機能を備えたウェブとして注目することができます。ここでは、免疫系と内分泌系が二重伝達に関与しています (Borre et al., 2014)。 最初に、微生物は、酢酸、プロピオン酸、酪酸ヒスタミン、他のピリミジン、グルタチオンなどのトランスモジュレーターとともに、神経伝達物質を代用、結合、および破壊することができます (Dinan and Cryan, 2017)。 これらの物質は脳内で神経伝達物質として働き、神経細胞を安定させます。 ただし、直接的な効果を示すには詳細な研究が必要です (Angelucci et al., 2019)。 さらに、腸内微生物叢は、中枢神経系の有害な代替物である他のタンパク質を生成します。これらは、宿主内の強力な刺激性サイトカインおよび先天性応答活性化因子 B 細胞です (Alam et al., 2017)。 したがって、天然の微生物叢は、さまざまな経路とともに、抗体を介した神経系および内分泌系を通じて、微生物叢 - 腸 - 脳の軸に影響を与えることができます (Gibson and Mehler, 2019)。 脳内のこれらの神経変化の結果は、破壊、高血圧、およびその他の一貫した疾患につながる可能性があります (Saunders et al., 2002; Galland, 2014; Johnson and Foster, 2018; Angelucci et al., 2019)。 腸内細菌叢の変化は、いくつかの神経疾患に関連しています (Cox and Weiner, 2018)。 これには、高血圧とストレス (Nagpal et al., 2018) だけでなく、神経変性疾患 (Quigley, 2017) と難治性てんかん (Braakman and Van Ingen, 2018) も含まれます。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 今日まで、腸-脳軸におけるバクテリオシンまたはバクテリオシン産生乳酸菌の役割を示唆する直接的な証拠はありません。 ただし、腸内微生物叢は産生株によって調節される可能性があり、腸 - 脳軸に間接的に影響を与える可能性があります。 in silico 研究では、神経伝達物質、アクアポリン、共生腸内細菌叢に対するナイシンの有益な効果が、ハイスループットシーケンスを使用して分析され、腸内細菌叢と腸脳軸で使用される神経化学物質との関係が明らかになりました。 ナイシンは、対照群およびシプロフロキサシン治療群と比較して、脳内でノルエピネフリンの最高の発現を示しました。 さらに、ナイシンで処理されたマウスは、盲腸サンプル中のLactobacillus, Bacteroides, およびBifidobacterium のレベルの増加と、病原性E. coli (大腸菌)およびenterococci(腸球菌)の減少を示したことがわかりました。 したがって、ナイシン、腸内細菌叢、およびマウスモデルにおける大腸菌によって引き起こされるストレスの減少の間には強い関係があります (Jia et al., 2018)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7.バクテリオシン産生乳酸菌とその疾患における役割 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸の正常な微生物叢の変化は、関節痛、免疫関連疾患、代謝障害、肝臓疾患、さまざまな胃腸疾患など、いくつかの慢性疾患を引き起こします (Carding et al., 2015)。 バクテリオシンは、宿主の微生物叢を形作る役割を果たしている可能性があり、間接的に、腸内毒素症の修正と宿主の健康の改善に重要な役割を果たしている可能性があります。 ここでは、腸内毒素症の形成によって発生するいくつかの重要な疾患と、バクテリオシンを産生するプロバイオティック乳酸菌を使用したそれらの治療の可能性について説明しました. 明確にするために、図 1 に図解します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
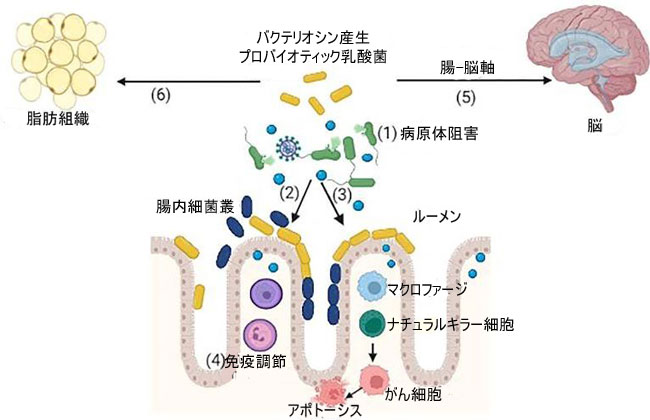
| 図 1 バクテリオシンを産生するプロバイオティクス乳酸菌は、いくつかの潜在的な機能を示しています | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| (1) 病原体の阻害、(2) 競合的排除によるプロバイオティクス細菌のコロニー形成、(3) マクロファージの活性化、ナチュラル キラー (NK) 細胞はさらにがん細胞と相互作用してアポトーシスを引き起こす、(4) 免疫調節、(5)腸内微生物叢のバランスをとる腸-脳軸、(6) 脂肪組織を減らすことによる抗肥満活性 (BioRender で作成)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
8.結腸感染症 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| バクテリオシンおよび/または乳酸菌のバクテリオシン産生株は、重度の感染症を引き起こすいくつかの食品由来および臨床病原体の阻害について文書化されています。 乳酸菌バクテリオシンのほとんどは細孔形成剤であり、細胞膜と相互作用して、膜電位の散逸とATP流出を介して標的細菌を殺し、細胞死に至ります。 したがって、乳酸菌のバクテリオシンは、臨床抗生物質の代替として機能する可能性があり、細菌感染症の治療に適用できます (Pérez-Ramos et al., 2021; Sheoran and Tiwari, 2021; Li et al., 2022)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Clostridium difficileは結腸感染症の主な原因菌です。 ex vivo モデルを使用して、精製されたナイシンは、本来の腸内微生物叢に影響を与えることなく、糞便微生物環境で C. difficile を選択的に枯渇させることができることがわかりました。 他の病原性細菌は、E. coli(大腸菌), Salmonella typhi,(チフス菌)Campylobacter jejuni, Shigella(赤痢菌), および Yersinia enterocolitica です (Papaconstantinou および Thomas、2007 年)。 このような感染は、共生プロバイオティクス L. acidophilus の個体数を増やすことで最小限に抑えられることがわかっています (Yun et al., 2014)。 ラクトシンとキレート剤であるエチレンジアミン四酢酸の組み合わせは、E. coliに対して有効であることが判明しました (Belfiore et al., 2007)。 L. salivarius NRRLB は、ヒト消化管の腸内病原体 C. jejuni に対して活性なバクテリオシン OR-7 を生成します (Ilinskaya et al., 2017)。 エンテロシン P は、Staphylococcus(ブドウ球菌), Clostridium, L. monocytogenes, Enterococcus faecium, および E. faecalis を阻害します (De Kwaadsteniet et al., 2006)。 E. coli および L. monocytogenes 腸に対するバクテリオシン産生菌L. casei の阻害効果は、マウスモデルで発見されました (Soltani et al., 2021)。 L. citreum GJ7 によって生成されたキムチシン GJ7 は、in vitro で S. typhi を阻害しました (Chang and Chang, 2011)。 L. crustorum MN047 によって産生されたバクテリオシン BM1829 は、G1/S チェックポイントで細胞周期を停止させるか、膜の完全性を破壊することにより、 E. coli, S. typhi, およびS. aureus (黄色ブドウ球菌)を阻害しました (Yan et al., 2021)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 進行中のパンデミックの間、ウイルス感染は深刻な病気と死亡率を引き起こしています。 このような感染症の治療に成功したことが最近証明された、提案および試験された多くの抗ウイルス剤があります。 しかし、これらの治療法は毒性を示し、症状を完全に軽減することはできませんでした. したがって、ウイルス感染症の安全な治療法を見つけることが重要です (Lehtoranta et al., 2020; Villena et al., 2020)。 プロバイオティクス乳酸菌は、ウイルスとの直接的な相互作用、バクテリオシンの生成、または宿主の自然免疫の増強によって抗ウイルス効果を発揮します (Al Kassaa et al., 2014; Tiwari et al., 2020)。 バクテリオシンは、ウイルス複製の後期段階で糖タンパク質の合成をブロックすることにより、多くのウイルスに対して抗ウイルス活性を示します (Huang et al., 2021)。 急性胃腸炎は、主にロタウイルス、ノロウイルス、アデノウイルスが原因で5歳未満の乳幼児に発症します。 ロタウイルスは、乳児の上皮細胞内層の破壊を引き起こし、下痢を引き起こす二本鎖 RNA 欠損エンベロープです (Li et al., 2021)。 Lacticaseibacillus rhamnosus GG は、オートファジータンパク質、p-mTor、および VPS34 陽性細胞、Beclin 1 および ATG16L1 の量を低下させることにより、子豚の腸内でヒト ロタウイルスおよび誘発されたオートファジーを抑制します。 L. rhamnosus GG はまた、p53 タンパク質のレベルを上昇させ、感染した腸細胞のアポトーシスを誘導します (Wu et al., 2013)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ウイルス感染に重要ないくつかの酵素反応は、バクテリオシンまたはバクテリオシン様物質によって阻害されます (Salman et al., 2020)。 ナイシンの経口投与により、マウスの CD4+ および CD8+ T リンパ球のレベルが上昇し、B 細胞が減少することが観察されました (Dey et al., 2021)。 Enterococcus faecium CRL35 によって産生されるエンテロシン CRL35 は、ヒトの腸関連潰瘍性疾患を引き起こす単純ヘルペス ウイルスの複製を阻害します (Wachsman et al., 1999)。 Erwinia carotovora NA4 が産生するエルウィニアオシンNA4 はコリファージ HSA を死滅させ、Enterococcus faecium NKR-5-3 が産生するエンテロシン NKR-5-3C は HSV type 1 に対して拮抗活性を示します (Dey et al., 2021)。 ナイシンとサカシン A は非エンベロープ マウス ノロウイルスに対して有効であり、Enterococcus faecium ST5Ha によって生成されるバクテリオシン ST5Ha は、単純ヘルペス ウイルスに対して抗ウイルス活性を示します (Todorov et al., 2010; Lange-stark et al., 2014)。 ノロウイルスは、エンベロープを持たない一本鎖 RNA を持つ腸内ウイルスで、Calciviridae科に属します。 ノロウイルス感染者では、プロテオバクテリアの数が増加し、バクテロイデスが減少します。 糞便から単離されたプロテオバクテリアへのノロウイルスの直接結合は、腸内細菌叢の調節を示しています。 さらに、ウイルス上に存在する P 粒子の上皮細胞への付着は、 L. casei BL23 によって阻害される可能性があります (Salman et al., 2020)。 この証拠は、乳酸菌のバクテリオシンおよび/またはバクテリオシン産生株がウイルス感染を予防する可能性があることを示唆しており (Cavicchioli et al., 2018)、したがって、治療に応用する前に正確な作用機序を解明するには、さらなる研究が不可欠です。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9.炎症性腸疾患 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 炎症性腸疾患(IBD) は、潰瘍性大腸炎 (UC) および クローン病 (CD)を含む胃腸管の長期の赤血球形成疾患です(Bjarnason et al., 2019)。 炎症性腸疾患の病因はまだ決定されていません。 一般に、不適切な食事、微生物叢の破壊、うつ病によって悪化します。 ただし、E. coli、C. concisus、および Mycobacterium avium などのいくつかの腸内微生物も炎症性腸疾患の病態生理学に関与しています (Ryma et al., 2021)。 潰瘍性大腸炎およびクローン病の病気には、遺伝的感受性、遺伝的傾向、生態学的トリガー、免疫系の変化、腸内微生物叢の異常な反応など、さまざまな要素を伴う病因の一定の挑発的な状態があります。 これらの疾患では、微生物の不均衡が患者に発生し、それは腸内毒素症(dysbiosis)を特徴としています (Sidhu and Vander Poorten, 2017)。 糞便微生物叢移植は炎症性腸疾患の潜在的な治療法ですが、その成功率は低いです (Colman and Rubin, 2014)。 アジュバントとしてのLactobacillus GG の有効性は、潰瘍性大腸炎患者の寛解維持において研究されています (Scaldaferri et al., 2013)。 プロバイオティクスは効果があり、炎症性腸疾患患者の忍容性が高いようですが、バクテリオシンの役割とそのメカニズムはまだわかっていません (Ryma et al., 2021)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 炎症性腸疾患に関連する患者の微生物叢は、健康な個人とは異なります (Shadnoush et al., 2015)。 最も豊富な腸内細菌の 1 つである Faecalibacterium prausnitziのような Firmicutesや Bacteroidetesの数が炎症性腸疾患中に減少し、Proteobacteria と Actinobacteria が増加することが観察されました。 したがって、これらの病気を克服するには、腸内細菌叢を安定させる必要があります。 ここで、バクテリオシンを生成するプロバイオティクスは、健康な微生物叢の成長を促進することによって重要な役割を果たすことができます (Furrie et al., 2005)。ただし、ある研究では潰瘍性大腸炎におけるプロバイオティクスの補給は潰瘍性大腸炎の治癒に有利であるが、クローン病には有利ではないことが示されています (Bjarnason et al. ら、2019)。 腸内微生物叢の変化は粘液層の欠陥を引き起こし、病原体に対する腸の透過性を高め、免疫応答を引き起こし、腸の炎症を引き起こします (Michielan, and D'Incà, 2015)。 バクテリオシンは、病原体を直接阻害/殺すことによって腸上皮の完全性を維持したり、乳酸菌が腸のニッチを占有するのを促進するコロニー形成ペプチドとして機能したりできます。 L. reuteri は、腸分泌ロイテリンの共生細菌であり、酵母、真菌、原生動物、ウイルスなどのいくつかの腸内病原体を阻害し、有益なグラム陽性菌の増殖を促進します (Liu et al., 2020)。 プロバイオティクスと、短鎖脂肪酸などのその代謝物が、腸内細菌叢異常症および炎症性腸疾患の免疫病因において重要な役割を果たすことが判明しました (Ryma et al., 2021)。 最近の研究では、母乳から分離されたL. casei, L. plantarum, L. rhamnosus, および L. acidophilusのバクテリオシン産生菌株が腸内病原体と競合し、ヒト結腸直腸腺癌細胞株 (HT- 29)、コレステロール値を下げ、マウスモデルで炎症性腸疾患を改善した (Abdi et al., 2021)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10.大腸がん | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 結腸直腸がんは、大腸の直腸と結腸に影響を及ぼし、血便と体重減少の主な症状を伴います。 食事、ライフスタイル、加齢などのさまざまな要因に左右されます (Center et al., 2009)。 強力な乳酸菌の有効性は、著名な臨床調査と動物モデル実験を通じて実証されました (Krebs、2016 年)。 マウスモデルに関する研究は、 Bifidobacteriumとバクテリオシン産生プロバイオティクスの組み合わせが結腸直腸癌の可能性を減らすという証拠を提供します (O'Callaghan and Sinderen, 2016)。 L. acidophilusは、単独または組み合わせて、結腸直腸癌に対する免疫を高めることがわかった (Zhong et al., 2014) プロバイオティクス細菌は、バクテリオシン、毒素、酵素など、抗がん作用を持つ多数の物質を分泌します。 L. lactis によって生成されるナイシン A は、腫瘍細胞の増殖を阻害し、肝細胞癌 (HepG2) の膜の完全性を変化させます。 ナイシンは細胞膜に細孔を形成し、内因性経路を介してアポトーシスを誘導し、メラノーマ細胞の増殖を低下させることによって抗転移剤としても機能します (Norouzi et al., 2018)。 さらに、P. acidolactici K2a2-3 によって産生されるペディオシンは、ヒト結腸腺癌細胞 (HT29) の増殖を阻害します (Soleimanpour et al., 2020)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HT-29 細胞株に対するE. coli によって産生されたコリシン E7 の in vitro 効果は、p53 の発現について評価され、bcl-2 は bcl-2 の減少と p53 遺伝子発現の増加を示します (Taherikalani and Ghafourian, 2021 )。 マイクロシンは、細胞膜の脱分極、DNA の断片化、ホスファチジルセリンの放出、およびカスパーゼ活性を引き起こします (Baindara et al., 2018)。 in vitro 研究では、Pediococcus acidilactici K2a2-3 によって産生されるペディオシンは、用量依存的に HT-29 および DLD-1 細胞株に対して抗がん活性を示します (Kaur and Kaur, 2015)。 この証拠は、結腸直腸癌の治癒に対するバクテリオシンの直接的または間接的な役割を示唆しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 11.肥満 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 肥満は、腸内細菌叢の異常に密接に関連する代謝障害です。 プロバイオティクスは、腸内微生物叢を調節してこのような障害と闘うのに役立ちます. 腸内微生物叢は、腸ペプチドのシグナル伝達や神経系の変化を通じて、エネルギー摂取と満腹感のバランスをとることに関与しています。 腸内微生物叢に変化があると、調節シグナル伝達ペプチドのバランスが変化します。 したがって、腸内細菌叢を回復させることで肥満を治すことができます。 肥満の人では、Firmicutes/Bacteroidetesの比率に変化があります (Mazloom et al., 2019)。 不均衡は、グラム陰性の好気性菌と嫌気性菌のBacteroidetesの数の減少と、グラム陽性のFirmicutesの増加によって特定されました (Sze and Schloss, 2016)。 しかし、マウスの体重増加とフィットの代謝生理機能は、糞便/大便の微生物叢移植によって受け継がれる可能性があります (Turnbaugh et al., 2008; Liou et al., 2013)。 バクテリオシン産生プロバイオティクスは、脂肪酸の吸収を減少させ、脂肪細胞のサイズを縮小し、脂肪酸の酸化に関連する遺伝子の発現も増加させます (Wicinski et al., 2020)。 L. plantarum は TNFα の産生を刺激し、レプチン ホルモンの産生も調節します (Behrouz et al., 2017)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| プロバイオティクスは、細菌含有量を調節するバクテリオシンの産生によって間接的に肥満に影響を与えます (Million et al., 2013)。 ケフィアから分離されたL. mali APSによる治療は、マウスモデルの肥満を軽減します。 L. helveticus によって産生されるバクテリオシン PJ4 は、マウスモデルの炎症と体重の減少に効果的であることが証明されています (Bai et al., 2020)。 Heeney et al. (2019) は、プランタリシン EF 産生 L. plantarum NCMIB8826 を与えられたマウスが高脂肪食の消費を減らし、体重増加が約 10% 減少したことを調査しました。 アイソジェニック(ΔplnEFI)変異株LM0419を補充したグループにも同じことがありませんでした。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 12.結論と将来展望 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| プロバイオティクス乳酸菌によって産生されるバクテリオシンは、一般に、細孔形成によって標的細胞を殺す小さなカチオン性ペプチドです。 これらのペプチドは、Salmonella, Staphylococcus, Listeria, Clostridium, および Enterococcusなどの関連株や病原菌に対して抗菌活性を示します。 バクテリオシンは、ロタウイルス、ノロウイルス、アデノウイルスなどによるウイルス感染にも効果があります。 腸内微生物叢は人体の重要な部分であり、いくつかの身体機能を安定させる上で重要な役割を果たしています。 プロバイオティクスとそのバクテリオシンは、抗菌作用と免疫調節を通じて腸内微生物叢を調節する可能性があり、腸内のバランスの取れた微生物群集と宿主免疫を回復するのに役立ちます。 さらに、バクテリオシンの役割は、結腸直腸癌、炎症性腸疾患、および肥満においても実証されています。 したがって、メタゲノミクス、プロテオミクス、メタボロミクスなどの高度な技術を使用して、バクテリオシンのプロファイリングと、腸の生態学的ニッチの確立におけるそれらの役割について、腸内のプロバイオティクス細菌を特徴付ける必要があります。 そのような発明は、いくつかの慢性疾患の治療のための自然由来の新しい製品と戦略の発見につながります。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| この文献は、Front Cell Infect Microbiol. 2022; 12: 851140.に掲載されたBacteriocin-Producing Probiotic Lactic Acid Bacteria in Controlling Dysbiosis of the Gut Microbiota.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |