Sonakshi Rastogi and Aditi Singh |
概要 |
| 微生物の密度が最も高いのは、「腸内細菌叢」として知られる人間の消化管です。 注目すべきは、Firmicutes 門に属する Lactobacillus (乳酸桿菌:以下Lactobacillusと表示)属のメンバーは、腸内細菌叢の最も重要なプロバイオティクス細菌です。 これらの腸内に存在するLactobacillusは、互いに通信するだけでなく、腸のバリアの完全性、粘膜のバリア防御のバランスを取り、宿主の免疫応答を改善するために、腸の上皮内層とも通信します。 人体は、腸、肺、心臓、骨、または神経組織に影響を与えるいくつかの炎症性疾患に苦しんでいます. 主に胃腸管と離れた臓器との間の免疫学的シグナルの交換を通じて、免疫応答の調節におけるLactobacillus種とその成分 (代謝物、ペプチドグリカン、および/または表面タンパク質など) の重要な役割を裏付ける証拠が増えています。 Lactobacillusによって媒介されるこの双方向のクロストークは、抗炎症反応を促進し、喘息、慢性閉塞性肺疾患 (COPD)、神経炎症性疾患 (多発性硬化症、アルツハイマー病、パーキンソン病など)、心血管疾患、炎症性腸疾患(IBD)および患者の慢性感染症に関連する症状の改善をサポートします。 代謝障害、肥満、糖尿病は軽度の炎症を特徴としています。Lactobacillusは、酸化ストレス応答と炎症経路を調節することにより、代謝障害を緩和します。 骨粗鬆症は、骨の炎症および吸収にも関連しています。 Lactobacillusとその代謝産物は、強力な免疫細胞コントローラーとして機能し、骨の吸収と形成において調節的な役割を示し、骨の健康をサポートします。 したがって、このレビューではメカニズムを実証し、動物および臨床試験からのさまざまな臓器に関連する炎症性疾患の緩和におけるLactobacillusの利点の証拠を要約しました。 現在の物語のレビューでは、宿主の健康を促進するために、腸内に生息するLactobacillusと遠隔臓器の免疫成分との間の複雑な相互作用を詳細に調査しています。 |
| 目次(クリックして記事にアクセスできます) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2. 腸内細菌叢の一員としての乳酸菌属 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1. 消化管バリアの完全性 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1.1. 腸上皮バリア | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1.2. 粘液産生 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1.3. 抗菌ペプチド/因子 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.2 .胃腸の炎症性疾患 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.3. 呼吸器炎症性疾患(腸-肺軸) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.4. 神経炎症性および神経変性疾患 (腸-脳軸) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.5. 循環器疾患(腸-心軸) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.6. 代謝障害 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.6.1. 肥満 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.6.2. 糖尿病 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.7. 骨の健康(腸-骨軸) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4. 今後の展望 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5. 結論 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1. はじめに | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 人体には、「マイクロバイオーム」として一般に知られている、宿主の生理機能を調節する可能性のある何兆もの動的で多様な微生物群集が生息しています (Malard et al., 2021)。 ヒトマイクロバイオームは、免疫系の恒常性、宿主代謝の調節、病原体の侵入の防止、上皮バリア機能の改善などの必須の生物学的機能を果たします (Malard et al.,2021)。 微生物の密度が最も高いのは、「腸内細菌叢」として知られるヒト消化管です (Dwivedi et al.,2021. 2019)。 注目すべきは、Firmicutes門に属するLactobacillus属のメンバーは、腸内細菌叢の最も重要なプロバイオティクス細菌です。 これらの腸内に存在するLactobacillus種は、互いに通信するだけでなく、腸のバリアの完全性、粘膜のバリア防御のバランスを取り、宿主の免疫応答を改善するために、腸の上皮内層とも通信します (Martín et al., 2019)。 さらに、Lactobacillus種は、腸内の機能的ニッチに生息する日和見病原体を競合的に排除し、上皮への病原体の付着を抑制し、乳酸、酢酸、プロピオン酸、バクテリオシン、および活性酸素種( ROS)を生成することによって病原体を直接殺すことにより、微生物の役割を示します (Dempsey and Corr, 2022)。 微生物の役割に加えて、共生Lactobacillus種は、T 細胞、ナチュラルキラー (NK) 細胞、マクロファージ分化、サイトカイン産生、刺激性Toll 様受容体 (TLR) を誘導することにより、獲得免疫応答と自然免疫応答の両方を調節します。 それらはまた、免疫グロブリン-A (IgA) 産生 B 細胞の発現を粘膜固有層のパイエル板で増加させることによって免疫調節効果を示し、そこで腸上皮への病原体の接着をブロックします (Cristofori et al., 2021)。 人体は、腸、肺、または神経組織に影響を与えるいくつかの炎症性疾患に苦しんでいます。 T-ヘルパー17 (Th17) 炎症細胞とその特徴的なサイトカインである IL-17F、および腫瘍壊死因子-アルファ (TNF-α) の発現をダウンレギュレートすることにより、炎症反応を抑制する乳酸桿菌種の重要な役割を裏付ける証拠が増えています。 心血管関連疾患(CVD)、骨粗鬆症、糖尿病や肥満などの代謝性疾患の症状を改善する上でのこれらの共生生物の有益な役割を強調する数多くの文献が利用可能です(Zaiss et al.,2019; Archer et al.,2021; Archer et al.,2021; Companys et al., 2021)。 この属は、コレステロール代謝や、トリメチルアミン-N-オキシド (TMAO)、短鎖脂肪酸 (SCFA)、リポ多糖 (LPS)、胆汁酸 (BA) などの腸由来の代謝産物の生成も調節することが報告されています。 骨粗鬆症は、骨の炎症と吸収に関連しています (Britton et al., 2014)。 Lactobacillus属とその代謝産物は、強力な免疫細胞コントローラーとしても機能し、骨の吸収と形成において調節的な役割を示し、骨の健康をサポートします (Zaiss et al., 2019)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| これらの考慮事項を考慮すると、ヒト疾患の緩和および治療におけるLactobacillus種の重要な役割は、魅力的な治療の可能性を示しています。 したがって、このレビューの目的は、宿主の健康に影響を与える免疫応答の媒介におけるこれらの微生物とその成分の強力な使用を示唆する現在の研究成果を分析することです。 PubMed は、キーワード「乳酸菌」または「腸内細菌叢」および「炎症」を使用して、過去 15 年間に発表されたすべての研究を検索するために使用されました。 550 以上の論文が見つかりましたが、英語で出版され、人間の病気に関連する側面に関するデータを提供するものだけが評価に含まれていました。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2. 腸内細菌叢の一員としての乳酸菌属 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 今日、腸内細菌叢は、人間の健康と病理学において不可欠な役割を担っているため、科学研究の重要な分野です。 注目すべきは、Lactobacillus属のメンバーが最も重要なプロバイオティック細菌です。 Lactobacillus属は、管腔の pH の調節、粘液産生の増加によるバリア機能の強化、抗菌ペプチドの分泌、および腸内細菌組成の変化によって作用します (Dempsey and Corr, 2022)。 それらの説得力のある機能的属性には、消化の改善、消化管バリアの完全性の維持、日和見病原体との競合、神経調節、初期の免疫系の成熟への参加、生涯にわたる免疫恒常性の維持、代謝産物、ビタミンおよびその他の成分の産生が含まれます ( Dempsey と Corr、2022 年)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細な分類プロファイリングにより、Lactobacillus属は Firmicutes門、Bacilli綱、Lactobacillales目(order)およびLactobacillaceae 科 に属することが明らかになりました。 それらは、グラム陽性、カタラーゼ陰性、非胞子形成性、ゲノムの GC (グアニンおよびシトシン) 含有量が低い偏性糖分解桿菌または球桿菌です (Zheng et al., 2020)。 Lactobacillus属のメンバーは、口腔、胃腸、および膣管に持続する敵対的な環境によく適応しています。 健康な成人の糞便では、さまざまな乳酸菌種の濃度が最大 105 ~ 108 CFU/g になります。 ヒトの腸内に生息する微生物種の中で、 Lactobacillus crispatus, Lactobacillus gasseri, Lactobacillus ultunensis, Ligilactobacillus ruminis, Limosilactobacillus reuteri, Lactobacillus kalixensis, Lactocaseibacillus casei, Limosilactobacillus gastricus, Limosilactobacillus antri, Lactobacillus rhamnosus, Ligilactobacillus salivarius, Limosilactobacillus fermentumなど永続的であり、不可欠な部分を形成することがわかっています (Zheng et al., 2020)。 乳酸菌種によるプロバイオティクスの補給は、腸内細菌叢の組成を変化させるのに役立ち、それによって腸内細菌叢を減らし、より豊富な有用な細菌で微生物バランスを維持します。 L. rhamnosus GG, L. acidophilus, L. plantarum, L. paracasei, および L. delbrueckii を含むプロバイオティック製剤は、経口投与すると、ラットで抗炎症活性を示したPrevotella や Oscillibacterなど他の腸内微生物種を刺激する傾向があります (Zeng et al., 2021)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.乳酸桿菌と人の健康 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| プロバイオティクスとして、Lactobacillusはさまざまな腸内細菌叢由来の代謝産物を介して重要な役割を果たし、慢性疾患の緩和、免疫系の刺激、病原体の保護、および栄養生理学など、それらに共通するいくつかの健康改善属性を持っています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1. 消化管バリアの完全性 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸のバリアが機能不全になると、過敏性腸症候群 (IBS)、炎症性腸疾患 (IBD)、肥満などのいくつかの炎症状態につながります。 腸内乳酸菌 は、マウスの炎症モデルにおいて直接的または間接的に胃腸のバリア機能を回復させることがわかった (Martín et al., 2019)。 それらが免疫系と相互作用する間接的なメカニズムには、腸内細菌叢の調節、腸上皮バリアの完全性の調節、粘弾性ムチン層の特性、拮抗ペプチド/因子の産生、および病原体の競合的排除が含まれます (Cristofori et al., 2021)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1.1. 腸上皮バリア | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸上皮バリアは、腸管内環境を管腔環境から分離し、それによって栄養素や他の分子の透過性を確認し、微生物や有毒化合物の侵入を阻止することによって保護的な役割を果たします。 共生Lactobacillus種は、タイトジャンクション、ギャップジャンクション、接着剤、デスモソームなどの多タンパク質複合体に依存する腸バリアの完全性を維持することが研究で報告されています (Kocot et al., 2022)。 これらの複合体は、主に膜貫通タンパク質、すなわちクローディン、オクルディン、および小帯オクルーデン (ZO) およびアクチン線維を介して隣接する細胞と相互作用する結合接着分子で構成されています。 慢性炎症性疾患または腸感染症の場合、腸のバリアの完全性が損なわれます。 研究では、腸管出血性大腸菌 O 157:H7 感染 Caco-2 細胞株における L. rhamnosus CNCM-I による ZO-1、オクルディン、およびクローディンタンパク質の発現増加が報告されています (Laval et al., 2015)。 同様に、L. caseDN-114 001 は、Caco-2 細胞で Toll 様受容体 (TLR)-2 の活性化を介して ZO-1 タンパク質の発現をアップレギュレートします。 TLR-2 結合はプロテインキナーゼ C (PKC) の活性化につながり、タイトジャンクションタンパク質の移動を引き起こし、それによって腸のバリア機能が強化されます (Kocot et al., 2022)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1.2. 粘液産生 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸上皮は、高分子量の糖タンパク質、主に杯細胞によって産生されるムチンで覆われています (Wuet al., 2020)。 粘弾性ムチン層は、消化酵素に対する保護を提供し、食物の通過をサポートし、粘膜固有層を横切る微生物の移動を防ぎ、腸の恒常性を付与します (Wu et al., 2020)。 研究では、特定のLactobacillus株がムチン遺伝子発現を調節し、それによって粘液層の属性を変化させ、腸の免疫応答に間接的に影響を与えることが報告されています。 注目すべきことに、L. rhamnosus CNCM I-3690 は、マウス炎症モデルの粘液産生杯細胞における Muc2 および Muc3 遺伝子発現を調節し、腸バリア機能障害および炎症を防止しました (Martín et al., 2019)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.1.3. 抗菌ペプチド/因子 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 乳酸菌株の重要な機能は、腸内病原体を相乗的に阻害し、宿主の免疫系を刺激することにより、病原性の増殖を防ぐ能力にあります。 歯周および腸内病原体に対してin vitroで拮抗活性を示した乳酸桿菌は、 L. oris, L. paracasei, L. crispatus, L. gasseri, L. salivarius, L. plantarum, L. delbrueckii, L. rhamnosus, L. acidophilus, および L. fermentum, 等で、それによって腸の恒常性が改善されます (NĘdzi-GÓra et al., 2020; Dempsey and Corr, 2022)。 Lactobacillus種の抗菌能力に寄与する既知の要因の中で。 有機酸、過酸化水素、一酸化窒素、短鎖脂肪酸、バクテリオシンなど、病原体の増殖を妨げる幅広い代謝物を生成する傾向があります (Sulijaya et al., 2020)。 有機酸、特に乳酸は、細胞質の pH を酸性にし、グラム陰性細菌の外膜を透過性にするため、強力な阻害因子です (Sulijaya et al., 2020)。 たとえば、L. acidophilus 4,356 は、乳酸を豊富に産生することにより、H. pyloriの増殖を抑制しました (Modiri et al., 2021)。 一酸化窒素 (NO) は、別の阻害性微生物代謝産物です。 プロバイオティクス乳酸菌は、一酸化窒素合成を効果的に上昇させるか、宿主マクロファージを刺激して一酸化窒素産生を促します。 たとえば、 L. fermentum LF1 は、l-アルギニンの l-シトルリンへの酸化を介した一酸化窒素シンターゼ経路を介して一酸化窒素を生成します (Dempsey and Corr, 2022)。 食品由来の病原菌や腐敗菌に対して広域スペクトルの阻害活性を持つバクテリオシンとして知られる短いペプチドをリボソームで合成するものさえあります。 たとえば、L. plantarum LPL-1 は 4,347 Da のプランタリシン LPL-1 (Wang et al., 2018) を生成し、クルバシリンA はL. curvatusに由来します (Ahsan et al., 2022)。 他のいくつかの乳酸菌のゲノムでは、ペディオシンとプランタリシンをコードする遺伝子が見つかっています (Ahsan et al., 2022)。 過酸化水素を産生する乳酸菌は、病原体の成長の停滞も引き起こします。 L. johnsonii UBLJ01 ゲノム解析により、H2O2 合成に関与する NADH オキシダーゼ、乳酸オキシダーゼ、およびピルビン酸オキシダーゼ遺伝子が明らかになりましたが、in vitro 試験ではそれらの強力な拮抗可能性が報告されました (Ahire et al., 2021)。 阻害物質の合成に加えて、乳酸菌は微生物毒素の分解、特に毒素の発現を阻害するか、病原体の外膜に結合することにより、毒性を低下させることができます。 たとえば、L. rhamnosus JB3 は、脂質ラフトを形成するか、病原性遺伝子の発現を下方制御することにより、腸内の H. pylori の感染を減少させました (Do et al., 2021)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 最近、宿主において発癌および免疫抑制を引き起こすことが知られているマイコトキシンの解毒における乳酸桿菌種の能力を強調する文献が利用可能です。 注目すべきことに、L. coryniformis BCH-4 と L. plantarum MiLAB 393 は、殺菌剤化合物であるシクロ (L-ロイシル-L-プロリル) と 3-フェニル乳酸シクロ (L-Phe-L-Pro) をそれぞれ生成しました。 これにより、アスペルギルスとカンジダ種の生存率が大幅に低下しました (Ström et al., 2002; Salman et al., 2022)。 さらに、Sunmola et al., 2019 は、一次複製部位が消化管にあるエコーウイルスなどのエンテロウイルスに対する L. plantarum および L. amylovorus AA099 の抗ウイルス活性を報告しました。 同様に、Kawahara et al., 2022 は、ヒト腸上皮 Caco-2 細胞におけるロタウイルスタンパク質 6 (VP6) 発現の増幅を抑制することにより、L. crispatusKT-11 株の S 層タンパク質の抗ウイルス活性を報告しました。 また、Mousavi et al., 2018 は、細胞表面での L. crispatusのマイクロコロニー形成が、単純ヘルペス ウイルス 2 粒子の侵入をブロックする傾向があることを示し、したがって、一次感染段階の阻害に役立ちます。 ロタウイルスは乳児に重度の反復性下痢を引き起こすウイルスの 1 つであり、単純ヘルペスウイルス I は成人に口唇ヘルペスを引き起こすウイルスの 1 つです (Kawahara et al., 2022)。 正確なメカニズムの理解はまだ限られていますが、そのうちのいくつかには、シアル酸とバクテリオシンの合成、免疫刺激、およびウイルスの結合の妨害が含まれます (Mousavi et al., 2018; Kawahara et al., 2022)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.2 .胃腸の炎症性疾患 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 炎症は、単球、B 細胞、T 細胞、樹状細胞 (DC)、ナチュラルキラー (NK) 細胞、およびマクロファージによって産生される炎症誘発性サイトカインによって誘発される、組織の損傷および/または感染に対する生理学的反応です。 大腸炎誘発マウスモデル研究からの実質的な証拠は、腸の炎症におけるLactobacillus種の抗炎症および免疫調節作用を主張しています (Cristofori et al., 2021)。 これらの効果は、サイトカインの産生と免疫細胞の増殖に大きく依存します。 IL-8 などの炎症誘発性サイトカインは、炎症反応中の免疫細胞の動員において重要な役割を果たします。 特に、 L. acidophilusは、TLR-2媒介マイトジェン活性化プロテインキナーゼ(MAPK)シグナル伝達経路の調節を通じて、IL-8産生を抑制し、Toll様受容体-2(TLR-2)および腸の炎症性上皮細胞における核因子カッパ軽鎖活性化 B 細胞 (NF-κB)の発現を増強することができます(Li et al., 2021)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 同様に、いくつかの乳酸菌株 ( L. acidophilus CCFM137、 L. fermentum CCFM381、L. plantarum CCFM634、CCFM734 など) も、TLR-2/TLR-6 ヘテロ二量体受容体の発現をアップレギュレートすることにより、抗炎症の可能性を示しました。 これは、炎症性細胞内シグナル伝達ネットワークとして機能します (Ren et al., 2016)。 炎症性腸疾患 (IBD) は、クローン病 (CD)、潰瘍性大腸炎 (UC)、顕微鏡的大腸炎、および嚢炎の 4 つの病理学的状態を引き起こす、消化管の腸上皮層を著しく混乱させる全身性疾患です。 Compare et al., 2017 は、 L. casei DG が炎症誘発性 IL-6、IL-8、TLR-4、および IL-1a を低下させ、感染後の クローン病被験者の結腸粘膜における IL-10 サイトカインレベルを上昇させることを報告しました。 さらに、L. casei とL. bulgaricusは、クローン病患者の結腸粘膜サンプルで炎症誘発性サイトカイン TNF-α を有意に低下させました (De Conno et al., 2022)。 Treg 細胞は、免疫調節と寛容に関与するもう 1 つの免疫学的プレーヤーです。 L. casei M2S01 は、Treg 細胞の活性化、IL-10 レベルを高め、腸内細菌叢を回復させることにより、クローン病や顕微鏡的大腸炎などの疾患において抗炎症作用を示しました (Liu Y et al., 2021)。 壊死性腸炎 (NEC) は、特に未熟児に影響を与える別の深刻な胃腸の炎症状態です。 L. rhamnosus HN001 で処理した壊死性腸炎乳児のイレウス由来のヒト腸細胞に対して実施された ex-vivo 研究では、TLR-4 の阻害による NF-kB 炎症経路の活性化の減少が示されました (Good et al., 2014)。 さらに、共生乳酸菌は IgA 抗体を増加させることで粘膜免疫を活性化し、病原体の固定化と凝集を引き起こします。 乳酸菌の用量依存的な消費は、特にL. plantarum, L. acidophilus, L. casei, L. delbrueckii subsp. bulgaricus, L. rhamnosusは、粘膜固有層に結合したIgA産生免疫細胞の数を増加させ、腸の上皮細胞に存在する受容体免疫グロブリンを刺激しました (Chen et al., 2022)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
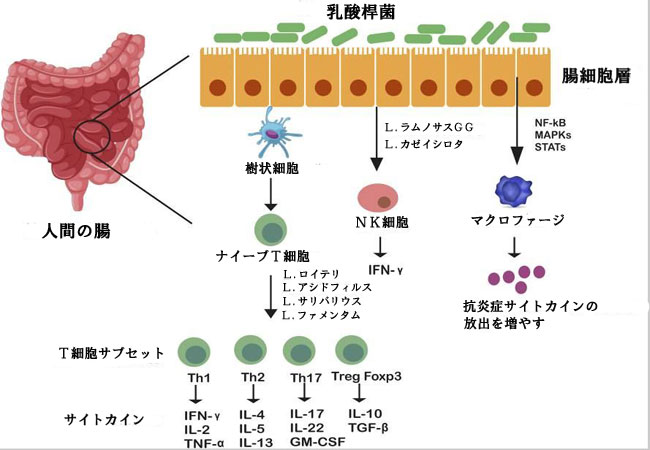
| 宿主の回腸管に付着する乳酸菌は、別の重要な側面であるアポトーシスとがんを調節します。これらはどちらも粘膜炎に関連しています。 たとえば、L. rhamnosus GG は抗アポトーシス Akt/プロテイン キナーゼ B を積極的に誘導し、p36 MAPK 経路を介してアポトーシス促進因子を阻害しますが、リポテイコ酸などの一連の乳酸菌の細胞壁成分は 一酸化窒素シンターゼを刺激する傾向があります。 それにより、病原体に感染した細胞死をもたらすイベントのカスケードが開始されます (Banna et al., 2017)。 重要な免疫イベントには、TNF-α サイトカイン産生によるマクロファージの活性化と、2 つの表面食作用受容体 (FcγRIII および TLR-2) の 一酸化窒素介在アップレギュレーションが含まれます。 Sun et al., 2020 は、抗炎症性サイトカイン遺伝子 (IL10、IL4、トランスフォーミング増殖因子-β3 (TGF-b3 および IFN-γ)) のアップレギュレーションとTLR-2およびNOD様受容体遺伝子のより高い発現とともに、炎症誘発性サイトカイン(IL6、IL1B、IL8、およびTNF-α)のダウンレギュレーションによるCaco-2 細胞におけるリポ多糖誘発性炎症反応を報告しました。 また、ゲノム解析により、L. gasseri JM1 による Caco-2 細胞のホスファチジルイノシトール 3-キナーゼ (PI3K)/プロテインキナーゼ B (Akt) シグナル伝達経路の活性化が明らかになりました。 別の in vivo 研究では、BALB/c 近交系マウスおよび Fisher-344 近交系ラットに対する L. acidophilus NCFB 1748 および L. paracasei DC412 による自然免疫応答および適応免疫応答の誘導が、多形核 (PMN) 細胞の動員、TNF-α 分泌および食作用を介して実証されました ( Azad et al., 2018)。 乳酸桿菌は、特に B 細胞やマクロファージで発現する TLR によって認識されるメチル化されていない CpG モチーフなど、ゲノム内の特定のモチーフをコードしています (Xiao et al., 2022)。 免疫応答の調節における乳酸桿菌種の役割を図 1 に示します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 図 1 乳酸桿菌属が腸内で媒介する免疫調節 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 細菌全体またはその成分としてのLactobacillus 属は、マクロファージおよび樹状細胞に発現する膜受容体 TLR-6 および TLR-2 を介して通信し、T 細胞サブセットの分化を刺激します。 これらの T 細胞 (Th1、Th2、Th17、Treg) は、炎症のマスターと見なされます。 乳酸菌属はまた、転写因子、STAT、NF-kB を活性化または抑制する MAP キナーゼを介して免疫細胞 (マクロファージなど) の細胞内経路を変更し、抗炎症性サイトカインの放出を増加させることによって、その機能を発揮します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 最近の研究では、痛みの管理とアレルギー症状の治療におけるLactobacillus属の免疫調節の役割を含む研究が勢いを増しています. 今日まで、TNF-α、IL-6、IL-1βなどの炎症誘発性サイトカイン、および宿主の腸上皮におけるCOX IIの発現を調節することにより、炎症性疼痛を緩和する乳酸桿菌種の能力を実証した研究者はほとんどいません(Santoni et al.,2021)。 炎症性疼痛の緩和に関与する正確な経路とメカニズムはまだ詳細に調査されていませんが. L. reuteri DSM17938 は抗侵害受容活性を示し、機能性腹痛のある子供の痛みを軽減しました (Jadrešin et al., 2020)。 考えられるメカニズムには、神経終末および/またはタイトジャンクションの感受性を引き起こす粘膜下免疫細胞の活性化が含まれます。 Martoni et al,2020による別の無作為化プラセボ対照二重盲検多中心研究 では、B. lactis UABla-12 およびL. acidophilus DDS-1 の投与後6週間以上の成人被験者の腹痛および炎症性腸疾患症状の改善が報告されました。 Maixent et al., 2020 は、炎症性腸疾患患者の疼痛緩和における 2 つのL. acidophilus 株のカクテルの有効性も報告しました。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.3. 呼吸器炎症性疾患(腸-肺軸) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 細菌全体またはその成分 (ペプチドグリカン、代謝物、表面タンパク質など) としてのLactobacillus は、慢性呼吸器疾患の患者の治療において免疫調節の役割を果たすことが示されています。 Lactobacillus が肺の免疫を調節し、呼吸器の健康を促進するメカニズムは、双方向の「腸-肺軸」によるものと考えられます (Du et al., 2022)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸粘膜では、免疫細胞に存在するパターン認識受容体 (NLR や TLR などの PRR) が n乳酸桿菌種またはその成分を認識し、リンパ循環を通じて肺組織に到達する傾向がある自然免疫細胞の活性化をもたらします (Stavropoulou et al., 2021; Du et al., 2022)。 解明するために、 L. paracasei CNCM I-1518 の経口投与により、先天性リンパ系グループ 3 細胞 (ILC3) が腸から肺に移動し、そこで ILC3 が肺炎に対する耐性を提供します (Gray et al., 2017)。 さらに、L. rhamnosus GG および L. murinusの経口補給は、Treg 細胞の肺への移動を促進し、それによって肺の炎症を増強します (Zhang et al., 2018; Han et al., 2021)。 Treg 細胞は、強力な抗炎症能を有するだけでなく、宿主の Th2 型免疫応答をブロックすることもわかっています。 これに加えて、胃粘膜とのLactobacillusの相互作用は、免疫細胞によるサイトカインの分泌をもたらし、循環を通じて肺組織に到達し、そこで免疫応答を変化させます。 Kollinget al., 2018 は、Lactobacillus CRL1505 を経口摂取すると、肺の TNF-α、IFN-β、IFN-α、および IFN-ɣ サイトカインのレベルが高くなり、呼吸器の炎症が大幅に減少すると報告しました。 注目すべきことに、特定のLactobacillus株は代謝産物、特に宿主の肺免疫応答を調節できる酢酸、プロピオン酸、酪酸などの短鎖脂肪酸 (SCFA) を分泌します。 短鎖脂肪酸は 2 つの方法で免疫応答に影響を与えます。 1 つは、代謝されていない短鎖脂肪酸が循環を介して肺に直接移動し、G タンパク質共役受容体 (GPCR) の活性化またはヒストン脱アセチル化酵素の阻害を促進する場合です (Koh et al., 2016)。 もう 1 つは、短鎖脂肪酸が循環によって骨髄に移動し、そこでマクロファージと樹状前駆細胞 (MDP) の分化を促進し、それらを Ly6C-単球に変換する場所です。 これらは次に肺に到達し、抗炎症性代替活性化マクロファージ (AAM) に分化します。 これらの抗炎症性代替活性化マクロファージは、好中球の動員を減らし、Treg 細胞を刺激して抗炎症性サイトカイン (IL-10、TGF-β) を生成させ、肺の損傷と炎症を軽減します (Anand and Mande, 2018; Du et al., 2022)。 動物研究では、より高い酪酸産生が、GPR109A受容体活性化によって媒介されるTreg細胞およびIL-10産生の増加、ならびにヒストン脱アセチル化酵素の阻害による肺組織におけるIL-10の回復につながることが報告されています(Vieira et al., 2019)。 Spacova et al., 2020 は、L. rhamnosus GR-1 が、Th2 を介した免疫応答を調節し、アレルギー性喘息モデルの腸内細菌叢の組成を変化させることで、気道の炎症と活動亢進の重症度を有意に防止し、腸-肺軸の存在を支持することを示しました。 同様に、Li et al., 2019 は、腸内細菌叢およびハウスダストの気道炎症に対する 6 つのlactobacillus種 (L. fermentum, L. salivarius, L. rhamnosus, L. casei, L. reuteri および L. gasseri) のダニ (HDM) 治療喘息マウス モデルで有効性を調査しました。。 これらの中で、L. reuteri の菌株は、腸内細菌叢の変化を伴う総 HDM-IgG1、IgE、および Th2 関連の炎症誘発性サイトカインの減少により、改善された気道炎症を示しました。 要約すると、乳酸桿菌のメンバーまたはその代謝産物は強力な抗炎症反応を示し、それによって患者の喘息、呼吸器感染症、および慢性閉塞性肺疾患 (COPD) の症状の緩和をサポートします。 さまざまな臓器系の炎症性疾患におけるLactobacillus種に関連する動物および臨床試験を表 1 (訳者注:表が大きいため1部割愛)にまとめます。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.4. 神経炎症性および神経変性疾患 (腸-脳軸) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 人間の脳の発達と機能における腸内常在菌種の役割も報告されています。 このメカニズムには、胃腸管と中枢神経系 (CNS) との間の神経、ホルモン、および免疫学的シグナルの交換が含まれ、主に「腸-脳軸」として知られています。 この双方向通信は、トリプトファン前駆体と、γ-アミノ酪酸 (GABA)、ヒスタミン、5-ヒドロキシトリプタミン (5-HT)、グルタミン、リポ多糖、分岐鎖アミノ酸 (BCAA)、胆汁酸、短鎖脂肪酸およびカテコールアミン などの微生物代謝産物によって媒介されます。 これらは、神経膠細胞機能、神経新生、髄鞘形成、血液-脳関門機能、およびシナプス剪定に関与する強力な経路を調節します (Suganya and Koo, 2020)。 注目すべきは、ヒトの腸内分離株である乳酸菌が、腸神経系 (ENS) でγ-アミノ酪酸を生成することができたことです。 研究では、L. rhamnosusJB-1 の補給による腸内細菌叢の調節が、γ-アミノ酪酸受容体の発現を活性化し、その結果、迷走神経を介したマウスの認知反応が著しく改善されたことが報告されています (Breit et al., 2018)。 Lactobacillusのいくつかの種は、神経調節物質および神経伝達物質 (モノアミン、セロトニン、脳由来神経栄養因子など) として顕著な効果を示しています (Suganya and Koo, 2020)。 主に腸内の乳酸桿菌によって産生される短鎖脂肪酸などの微生物代謝産物は、循環に入るか血液-脳関門 (BBB) を通過することにより、迷走神経、内分泌、液性および免疫経路を介して脳の神経機能に直接影響を与えます。 短鎖脂肪酸は、脳機能を維持する調節性サイトカインのレベルを高めるために、Treg 細胞、内分泌細胞、神経細胞を直接活性化する傾向があります (Silva et al., 2020)。 リポ多糖 は、TLR 刺激、特にミクログリア細胞とアストロ サイトの TLR4 を介して、神経変性および神経炎症性疾患を誘発します。 ミクログリア細胞上の リポ多糖/TLR4 シグナル伝達は、特に自閉症スペクトラム障害 (ASD) 患者の腸または中枢神経系における炎症性サイトカインのレベルを高めることによって、中枢神経系に影響を与えることが研究で報告されています。 Liu et al., 2019 は、 L. plantarum PS128 を 1 ヵ月間経口補給すると、二重盲検無作為化プラセボ対照試験で、プラセボ群と比較して自閉症スペクトラム障害関連の症状が劇的に改善したと報告しました。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
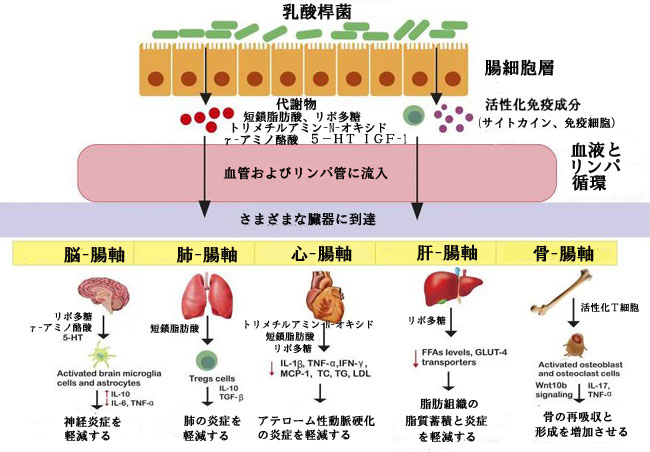
| 既存の証拠は、実験モデルまたは臨床設定における神経炎症性および神経変性疾患の緩和における乳酸桿菌種とその有益な代謝産物の重要な役割を支持しています。 多発性硬化症 (MS) は、ミエリン鞘の分解と軸索損傷を特徴とする神経炎症性自己免疫疾患です (Suganya and Koo, 2020)。 Kwon et al., 2013 は、L. casei, L. acidophilusおよびL. reuteniを実験用マウスに経口投与すると、Foxp3+ および IL10 + Treg の発現が増強され、末梢免疫系および炎症部位において炎症誘発性 Th1/Th17 分極が減少することにより、多発性硬化症の進行が遅延したことが報告されました。 別の同様の研究では、L. paracasei DSM 13434 および L.plantarum PDSM 15312 を補給すると、特にマウスで CD4+CD25+Treg を産生する IL-10 によって抑制された多発性硬化症症状が抑制されることが示されました (Lavasani et al., 2010)。 これに加えて、ヒト腸分離株 L. reuteri NK33、L. mucosae NK41、B. longum NK46、および B. adolescentis NK98 は、マウスのストレス誘発性不安/うつ病を軽減することが報告されています。 考えられるメカニズムには、NF-κB 経路を遮断し、血清中の リポ多糖、コルチコステロン、IL-6、および TNF-α レベルを低下させることによる、腸内細菌組成および炎症性免疫応答の調節が含まれます (Jang et al., 2019)。 アルツハイマー病 (AD) は神経変性疾患であり、高齢者では記憶、活動、認知能力、および思考の大幅な低下が観察されます (Suganya and Koo, 2020)。 研究によると、L. acidophilus, B. bifidum および B. longum をラットに 12 週間摂取させると、空間学習と記憶、長期増強 (LTP)、ペアパルス促進 (PPF) 比、および脂質プロファイルが大幅に増強されることが示されました (Rezaei Asl et al., 2019)。 さらに、Castelli et al., 2020 は、pTrK、P13K/Akt、p-CREB、pERK5 経路を活性化し、腸内細菌叢を回復させることにより、ヒト神経芽細胞腫細胞におけるLactobacillus種の神経保護的役割を支持しました。 パーキンソン病 (PD) は、中枢神経系と末梢神経系の両方が関与する、気分の変化、認知障害、安静時振戦、動きの鈍さ、自律神経機能不全、感覚と睡眠の交替を特徴とする、もう 1 つの一般的な神経変性疾患です (Suganya and Koo, 2020)。 研究は、炎症誘発性サイトカイン (TNF-α、IL-6、IL-17A) および活性酸素種 (ROS) レベルの低下と、末梢血単核細胞 (PBMC) の抗炎症性サイトカイン (IL-4、IL-10) を増加させ、パーキンソン病 (PD) 関連の症状を緩和における L. acidophilus (LA02) および L. salivarius, (LS01) の役割を支持しています (Magistrelli et al., 2019)。 腸と離れた臓器の間の複雑なクロストークを図 2 に示します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 図 2 宿主の健康に影響を与える乳酸桿菌属媒介の双方向クロストーク | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 代謝物および/または活性化された免疫成分を介してLactobacillus属は循環に入り、炎症反応を促進または抑制するさまざまな経路に入る遠隔臓器に到達します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.5. 循環器疾患(腸-心軸) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
心血管疾患(CVD)の緩和における腸内の乳酸桿菌種の役割を記録した大量の文献が入手可能です。具体的には、うっ血性心不全、心筋梗塞、アテローム性動脈硬化症、冠動脈疾患、狭心症、末梢血管疾患、動脈瘤などの心臓および血管の疾患が含まれます (Companys et al., 2021)。 高血圧と高血清コレステロールは、心血管疾患の主要な素因の 1 つです。 腸内の乳酸桿菌のコロニーは、アテローム性炎症反応、コレステロール代謝、酸化ストレス反応、および トリメチルアミン-N-オキシド、短鎖脂肪酸、リポ多糖、胆汁酸 などの腸由来の機能的代謝産物の産生を調節することにより、心臓に保護効果を発揮します (Companys et al., 2021; Papadopoulos et al. 、2022)。 注目すべきことに、研究者は、乳酸菌の介入が、選択的な腸内細菌叢の豊富さと多様性を調節することにより、心臓保護の役割を大幅に与える可能性があることも支持しています (Zhao et al., 2021)。 例として、L. salivarius Ls-33 補給は、肥満心臓患者のPrevotella-Bacteroides–Porphyromonas group/Firmicutes比を増加させることにより、腸内細菌叢の組成を変化させました (Zhao et al., 2021)。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 人体では、血中のコレステロール値が高いことが、動脈にアテローム硬化性プラークが発生する主な素因の 1 つと考えられています。 腸内の乳酸菌は、コレステロールの同化と、トリグリセリドと脂肪酸の代謝において重要な役割を果たしています。 提案された作用機序には以下が含まれます。 1) 微生物の胆汁酸塩加水分解酵素 (BSH) の作用による胆汁酸塩の分解、比較的溶解度の低いものにするため、腸上皮層による再吸収が減少します。L. gasseri LBM220 (Rastogi et al., 2021) および L. mucosae SRV5 および SRV10 (Rastogi et al., 2020) で観察されるように、糞便中への排泄を増加させます。 2) 微生物によるコレステロールの比較的可溶性の形態であるコプロスタノールへの変換 3) L. acidophilus ATCC 43121株で観察された微生物膜によるコレステロールの取り込み 4) 短鎖脂肪酸の生産。 短鎖脂肪酸は、肝臓の 3-ヒドロキシメチル-3-グルタリル-CoA レダクターゼ酵素を阻害することにより、肝臓のコレステロール合成を直接妨げる可能性があります (Amiri et al., 2021)。 腸内細菌叢分析により、マウスモデルにおける乳酸桿菌の補給が、Ruminococcus, Eubacterium および Roseburiaなどの共生生物を代謝する短鎖脂肪酸を選択的に促進し、それによって発酵短鎖脂肪酸のより高いレベルを促進することが明らかになりました(Amiri et al., 2021)。 この研究では、L. fermentum 296 を高給餌ラット モデルに 4 週間与えた場合、短鎖脂肪酸の産生が増加したため、HDL が大幅に増加し、有害な LDL が減少したことも示されました (Cavalcante et al., 2019)。 さらに、L. plantarum 06CC2 などの特定のLactobacillus株は、高コレステロール血症に直接関与しています。 これは、HFD Balb/c マウスの血清低密度リポタンパク質 (LDL)、トリグリセリド (TC)、遊離脂肪酸レベル、アポリポタンパク質 B を著しく減少させ、アポリポタンパク質 A-I レベルを増加させました。 同時に、この株は、コレステロール代謝に関与する酵素の発現を調節することにより、脂質沈着のために肝臓組織も標的にします (Yamasaki et al., 2020)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 胆汁酸および 短鎖脂肪酸とは別に、腸由来の微生物代謝産物、特にトリメチルアミン-N-オキシドおよびリポ多糖も心血管疾患の進行に関与していることを示す証拠が増えています。 トリメチルアミン (TMA) は、消化管の共生生物によって食物中のコリンと L-カルニチンから代謝される代謝産物であり、吸収されると肝臓に到達し、そこでトリメチルアミン-N-オキシドに酸化されます。 トリメチルアミン-N-オキシドレベルが高いと、コレステロール逆輸送の阻害、動脈血管壁の炎症誘発性変化、マクロファージでの高コレステロール蓄積の誘導、血小板過敏性および動脈血栓症が引き起こされます (Zhao et al., 2021)。 研究によると、乳酸桿菌種は、トリメチルアミン産生に関与する重要な酵素を産生する腸内微生物の増殖を抑制することにより、トリメチルアミン-N-オキシド関連の 心血管疾患リスクを軽減します。 これは、2018 年に Qiu らによって実施された研究によって証明されました。 ここで、L. plantarum ZDY04 介入は、動物モデルにおけるMucispirillum属およびErysipelotrichaceae科、Lachnospiraceae,科、およびBacteroidaceae科の相対的な微生物叢集団を変化させることにより、トリメチルアミン-N-オキシド循環レベルの大幅な低下と トリメチルアミン-N-オキシド誘発性アテローム性動脈硬化症を引き起こします。 L. plantarum Dad-13 を肥満成人に補給すると、腸内のBacteroidaceaeの個体数が増加し、Firmicutes 菌叢が減少します (Rahayu et al., 2021)。 また、L. plantarumの補給は、心血管疾患患者の血清 トリメチルアミン-N-オキシドレベルと、IL8、IL-12、およびレプチンの循環炎症因子を直接減少させました。 さらに、細菌の外膜成分としての リポ多糖は、マクロファージを活性化して、動脈硬化や心不全の発生を促進する炎症性サイトカイン (IL-1、TNF-α、IL-12、IL-6、IL-8) を分泌することにより、心血管疾患を誘発します。 これは、心筋細胞におけるミトコンドリア脂肪酸の酸化のダウンレギュレーションによるものです (Zhao et al., 2021)。 また、リポ多糖 は、TLR4 を介した白血球カテプシン G 活性化を介して血小板凝集を増加させ、血栓症を引き起こします (Zhao et al., 2021)。 L. brevis は、宿主における リポ多糖誘導性炎症性サイトカインの発現をブロックすることにより、心血管疾患発症を軽減しました (Chang et al., 2016)。 同様に、L. paracasei FZU103 は、高給餌 (HFD)を与えられたマウスの リポ多糖誘発性炎症軸とコレステロール代謝を調節することがわかりました。 メタゲノム研究でも 腸内細菌叢の変化が示されており、腸内のAlistipes, Ruminococcus, Helicobacter およびPseudoflavonifractorの存在量が増加しているが、Tannerella, Blautia, および Staphylococcosが減少している (Lv et al., 2021)。 L. plantarum LP91 を与えた リポ多糖誘導マウスでは、アテローム性動脈硬化の炎症因子、特に TNF-α と IL-6、血管細胞接着分子、E-セレクチンの発現がダウンレギュレートされていることがわかり、心血管疾患症状の改善が見られました (Aparna Sudhakaran et al.,2013)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.6. 代謝障害 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 最近の多くの出版物は、軽度の炎症を特徴とする肥満や真性糖尿病などの代謝障害の発症を和らげるLactobacillus種の有益な役割を報告しています (Archer et al., 2021)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.6.1. 肥満 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 肥満は、エネルギー恒常性に関与する複雑な内分泌器官である脂肪組織 (AT) の変化に関連する多面的な代謝疾患です。 通常、脂肪組織は、脂肪細胞、リンパ球、マクロファージ、線維芽細胞、プラスミノーゲン活性化因子阻害剤 (PAI-1)、アディポネクチン、レプチン、血管調節因子アンギオテンシン II、およびサイトカインを産生する内皮細胞を包含しています (Andersen et al., 2016)。 肥満の発症は、血管新生、酸化ストレスレベル、分泌されたアディポカイン、および浸潤したリンパ球の炎症状態の生理学的変化を伴う機能不全の脂肪組織につながります(Jo et al.,2009)。 肥満患者では、遊離脂肪酸 (FFA) レベルの上昇がシグナル伝達経路、特に TLR の活性化を引き起こします。 これらは、腸の固有層から細胞内で TNF-α、IL-6、IL1β、レプチン、レジスチン、ケモカインなどの分子の産生を増加させることにより、炎症誘発性応答に寄与します (Khan et al., 2021)。 腸内では、TNF-αの活性化はアポトーシスシグナル伝達経路、遊離脂肪酸レベルを刺激し、GLUT-4トランスポーターの発現をダウンレギュレートします。 高給餌 (HFD) マウス群では、高グルコース、遊離脂肪酸、およびアポトーシスに応答して脂肪組織に CD8+ T 細胞、TNF-α、IFN-ɣ、および CX3CR1int マクロファージが浸潤し、炎症を引き起こします。 注目すべきことに、肥満における炎症性変化および腸上皮の完全性の喪失の要因の 1 つは、細菌エンドトキシンリポ多糖の漏出を含みます (Khan et al., 2021)。 これは、リンパ球の不適切な分布、サイトカインレベルの変化、腸内細菌叢、および食事抗原に対する免疫応答につながります。 多くの研究は、脂質蓄積、酸化的損傷、炎症、および腸内細菌叢を緩和することにより、肥満の治療における乳酸菌の重要性に集中しています。 最近の出版物では、炎症性バイオマーカーが減少した高給餌マウス群の過剰体重と脂肪蓄積の低下における L. curvatus HY7601 とL. plantarum KY1032 の相乗効果が報告されました (Park et al., 2013)。 Choi et al., 2020 は、L. plantarum LMT1-48 で処理した場合の高給餌給餌マウスの体重減少を報告し、脂質の蓄積、脂肪組織への免疫細胞の浸潤、および脂肪細胞のサイズが減少しました。 特に、乳酸桿菌の複数の株、特にL. casei IMVB-7280、L. paracasei HII0、L. paracasei CNCM I-4034、L. rhamnosusCGMCC1.3、L. rhamnosus LA68 および L. casei IBS041 は、 肥満症状の軽減、おそらく、体重増加の減少、コレステロール値の低下、肥満および炎症の低下においてプラスの効果が証明されている(Wiciński et al., 2020)。 同様に、L. reuteri GMNL-263 は、血清炎症誘発性因子レベルを低下させ、白色脂肪組織 (WAT) エネルギー代謝をリモデリングすることにより、高給餌を与えられたラットの肥満に関連する症状を改善しました (Chen et al., 2018)。 さらに、Joung et al., 2021 は、L. plantarumを投与した高給餌給餌マウスの脂肪組織における脂肪蓄積の低下を報告しました。 全体として、Lactobacillusは肥満に関連する症状の治療において顕著な結果を達成しています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.6.2. 糖尿病 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 慢性代謝性疾患である真性糖尿病は、一貫して高い血清血糖指数を特徴としています。 2015 年の世界疾病負担に関する研究では、糖尿病が都市人口の主な死亡原因の 1 つであることが報告されています。 糖尿病患者の炎症状態と代謝障害との間には有意な関連性がありました。 仮定されたメカニズムには、セリン残基をリン酸化することによってインスリン受容体 (IRS-I) を不活性化する、炎症誘発性サイトカインである TNF-α のレベルの上昇が含まれます。 他のサイトカインである IFN-γ、TNF-α、および IL-1β は、膵臓の β 細胞に浸潤し、その後細胞のアポトーシスと損傷を誘発することによって相乗的に働きます。 したがって、筋肉、肝臓、および脂肪組織における炎症誘発性サイトカインのレベルの上昇は、インスリン抵抗性を引き起こすインスリンシグナル伝達を阻害するため、糖尿病病理の主要な要因です (Tsalamandris et al., 2019; Bezirtzoglou et al., 2021)。 興味深いことに、L. plantarum Y44 は、特に調節性抗炎症性サイトカイン IL-10 を活性化することにより、肝臓、腸、筋肉組織で炎症誘発性サイトカイン遺伝子のダウンレギュレーションを示し、免疫調節の役割を強調しています (Liu et al., 2020)。 同様に、 L. casei Shirota 株は、ストレプトゾトシン(STZ)誘発糖尿病ラットモデルにおいて、炎症誘発性サイトカインIL-4、IL-6、およびC反応性タンパク質(CRP)レベルの低下を示しました(Zarfeshani et al., 2011)。 Archer et al. (2021) は L. fermentum MCC2759 は、STZ 誘発糖尿病ラットの肝臓、MAT、筋肉、および腸組織のグルコースプロファイルと炎症誘発性サイトカイン、IL-10 の減少を示したことも報告しました。 この研究では、インスリン感受性 (GLP-1、GLUT-4、アディポネクチン)、腸バリアの完全性 (ZO-1)、および TLR-4 受容体の発現の増強も証明されました。 これに加えて、L. casei は、IL-2 および IFN-γ 産生の抑制による免疫後応答の改善により、血清グルコース含有量を減少させることが報告されています (Qu et al., 2018)。 Park et al (2015) による別の研究では、L. rhamnosus GG (LGG) の投与によるオスの db/db マウスに対する抗高血糖効果が、ER(小胞体)ストレスの増加とマクロファージの抑制に関連しており、インスリン感受性の増加につながることが報告されています。 また、いくつかの糖尿病関連遺伝子の発現の変化も役割を果たします。 プロバイオティクスL. rhamnosus NCDC17 は、その GLUT4(グルコース取り込み関連遺伝子)、pp-1(グリコーゲン合成関連遺伝子)、PPAR-γ(インスリン感受性関連遺伝子)などのグルコース代謝およびインスリン感受性関連遺伝子のmRNA発現をアップレギュレートし、G6PC (糖新生関連遺伝子) のダウンレギュレーションする能力により抗糖尿病能力があると報告されています (Singh et al., 2017)。 さらに、プロバイオティクスの摂取は腸内細菌叢の組成に影響を与え、それが次に腸上皮を緩和し、TLR4 シグナル伝達経路を減少させることで免疫応答を抑制し、最終的にインスリン感受性を高めます (Li et al., 2021)。 いくつかの他の仮定されたメカニズムには、腸の L 細胞からのグルカゴン様ペプチド-1 (GLP-1) 分泌の増強が含まれ、膵臓ベータ細胞から放出されるインスリンのレベルを上昇させ、糖毒性を低下させることによって食後の高血糖を抑制します (Kesika et al., 2019)。 L. kafiranofaciens M および L. kefiri K の投与に関する研究では、GLP-1 分泌が刺激され、同時にグルコース代謝が上昇することがわかりました (Kocsis et al., 2020)。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.7. 骨の健康(腸-骨軸) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 代謝性骨障害である骨粗鬆症は、貧弱な骨組織、劣化した骨量および多孔性を特徴とし、その後、骨折しやすい弱くて脆い骨につながります (Akkawi and Zmerly, 2018)。 特に、細菌全体またはその代謝物および/または構造成分(短鎖脂肪酸、硫化水素(H2S)、インスリン様成長因子-I(IGF-I)、リポ多糖、ペプチドグリカンなど)としての乳酸桿菌種は、強力な免疫細胞コントローラとして機能し、また、骨の吸収と形成における調節的役割を示します (Zaiss et al., 2019)。 これにより、「腸-骨軸」におけるリンカーとしての役割が確認されました。 要因すなわちIGF-Iは、主に食事摂取に応答して肝細胞で産生され、腸内細菌は腸-骨軸を直接媒介します(Zaiss et al., 2019)。 微生物代謝産物である硫化水素は、ガス伝達物質として機能し、腸内常在菌の 乳酸桿菌によって生成され、Wnt シグナル伝達を活性化することによって骨形成と出生後の骨格発達を刺激します (Grassi et al., 2016)。 骨芽細胞では、wnt シグナル伝達の活性化が骨芽細胞形成の促進を媒介し、骨芽細胞のアポトーシスを阻止します。 さらに、短鎖脂肪酸は、骨組織を拡散させ、免疫応答を調節する能力について大きな注目を集めています (Zaiss et al., 2019)。 短鎖脂肪酸、特に酪酸とプロピオン酸は、成熟した Treg 細胞の増殖を誘導します。 Treg の成熟は、樹状細胞 (DC) に発現する GPR109a および GPR43 受容体に依存します。 Treg 細胞は、免疫抑制性サイトカイン IL-10 および TGF-β を産生することにより、骨の骨内膜表面に存在する CD4+ T 細胞を抑制します。 骨芽細胞の分化を直接増加させ、破骨細胞形成を減少させることにより、骨粗鬆症と骨密度(BMD)の改善をもたらします(Smith et al., 2013)。 また、活性化された Treg 細胞は、Treg/Wnt10b/Wnt シグナル伝達経路を介して骨同化作用を誘導するため、カルシオトロピック副甲状腺ホルモン(PTH) 刺激骨形成に必要です (Yu et al., 2018)。 L. rhamnosus LGG を 4 週間摂取すると、循環および腸の酪酸レベルが直接増加し、腸から骨などの離れた臓器に拡散する能力が確認されました。 骨組織では、酪酸がTregを誘導し、骨の健康を改善します。 Tyagi et al., 2018 によるこの研究はさらに、L. rhamnosus LGG が腸内細菌組成を変化させ、腸内で短鎖脂肪酸の生成を誘発することが知られているClostridiaの菌数が多いことを証明しました。 したがって、L. rhamnosus LGG は、短鎖脂肪酸、Treg、および骨形成をつなぐ経路を実質的に仲介します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 現在までに、複数の研究が動物モデルを利用して、一次および二次骨粗鬆症の両方の骨損失を減少させる乳酸菌の有効性を決定してきました。 L. reuteri ATCC 6475 を健康なオスのマウスに 1 か月間経口摂取させると、未処理の対照と比較して、骨ミネラル含有量、椎骨および大腿骨の骨梁の密度、骨梁の厚さ、および骨梁の数が改善されました。 骨密度の増加は、骨芽細胞マーカーであるオステオカルシンによって証明されるように、骨形成率のレベルを高めます。 L. reuteri ATCC 6475 は、骨髄と腸の両方で破骨細胞形成促進サイトカインと炎症促進サイトカインの遺伝子発現を全身的に抑制することによって作用します (Nilsson et al., 2018)。 これらの抗炎症効果は、他の種の乳酸桿菌でも見られるように、腸バリアを通過するカルシウム輸送を直接増加させる傾向があります。 免疫細胞の活性化は、カルシウムレベルに大きく依存します。 低カルシウム血症の場合、 L. casei, L. reuteri および L. gasseriを豊富に含むヨーグルトでラットを治療すると、カルシオトロピック副甲状腺ホルモン依存的にカルシウム吸収が改善されました。 同様に、L. rhamnosus (HN001) もラットモデルでカルシウムとマグネシウムの保持を改善しました (Collins et al., 2017)。 Ohlsson et al., 2014 は、単一のL. paracasei 株 (DSM13434) または 3 つの株の混合物 (L. plantarum DSM 15312、DSM 1531、および L. paracasei DSM13434) のいずれかでマウスを 14 日間水中で処理した結果、コントロールと比較して、皮質骨ミネラル含有量が増加し、カルシウムおよび再吸収マーカーC末端テロペプチドの尿分画排泄のレベルが低下した。 異なるLactobacillus株は、異なるおよび/または重複する経路を介して作用します。 L. helveticus などは、カルシウムの取り込みを高めることによって骨密度を増加させるだけでなく、生理活性ペプチドであるバリル-プロリル-プロリン (VPP) およびイソロイシル-プロリル-プロリン (IPP) も分泌します (Parvaneh et al., 2018)。 さらに、不溶性無機塩の可溶性形態への変換、腸内ミネラル吸収部位の保護、カルシウム結合タンパク質の調節のトリガー、ミネラルとフィチン酸との相互作用の最小化は、骨の健康をサポートするLactobacillus のさまざまな菌株によって報告された主な作用です。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4. 今後の展望 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| シーケンシング処理、ゲノム アセンブリ、および注釈技術の指数関数的な進歩により、何千もの公に利用可能なLactobacillus 属のゲノムが生まれました。 これらのデータへのアクセスは、プロバイオティクス細菌の分子ビューに革命をもたらし、常在細菌叢と粘膜免疫システムとの間の相互作用に関連する複雑さを解明する研究を大幅に加速させました。 特に、ゲノムツール、特に機能ゲノミクス、プロテオミクス、トランスクリプトミクス、およびセクレトミクスの進歩は、胃腸の健康をはるかに超えた複雑で動的な宿主微生物クロストークの理解に役立っています。 例として、腸と脳 (腸脳軸) 間の双方向クロストークに関する研究は、腸の恒常性の神経化学的重要性を明らかにしています。 同様に、腸と肺のクロストーク (腸肺軸) を含む研究は、宿主の免疫応答の調節における微生物代謝産物の重要性を強調しています。 これに加えて、乳酸桿菌は. 樹状細胞やマクロファージなどの抗原提示細胞 (APC) 上のタンパク質性微生物関連分子パターン (MAMP) とパターン認識受容体 (PRR) との相互作用を通じて粘膜免疫を調節します。 最近、いくつかのLactobacillus のプロテオミクスおよびゲノム プロファイルが生物情報学的にスクリーニングされ、さまざまな細胞外タンパク質をカタログ化したセクレトーム データベースが作成されました。 たとえば、プロテオミクスに基づく方法を使用して、L. acidophilusの S 層関連タンパク質 (SLAP) を特定しました。 抽出後、質量分析によってS 層関連タンパク質を特定し、セクレトーム データベースを参照しました。 S 層関連タンパク質の突然変異分析は、in vitro 細菌 樹状細胞共培養アッセイを使用した免疫調節表現型を示しました (Huang et al., 2021)。 ただし、各プロバイオティクスによって誘導される転写ネットワークは、研究した各菌株に固有のものであり、宿主内で異なる代謝および免疫原プロファイルを示すことに注意することが重要です。 したがって、プロバイオティクスおよび 腸内細菌叢研究の分野で行われた将来の進歩を反映して、実験的デザインによるヒト臨床試験の必要性が高まっています。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5. 結論 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| まとめると、腸内細菌種、特に腸内の乳酸桿菌種は、胃腸バリア機能、免疫刺激、および腸内細菌叢の回復における潜在的かつ有能なプレーヤーとして特定されています。 これまでのところ、宿主の健康を促進するために、腸内に生息する乳酸桿菌属またはその代謝産物と遠隔臓器の免疫成分との間の複雑なクロストークを強調してきました。 腸内生態系の健康的なバランスは、潜在的に炎症誘発性細胞 [IFN-γ を分泌する Th 細胞、インターロイキン IL-17 を分泌する Th17 細胞、および IL-22] および抗炎症 Foxp3+ 受容体 T細胞の間の監視メカニズムの回路によって維持されています。 乳酸桿菌の特定の菌株は、Treg の成熟を制御するか、IL-10 および IL-12 の産生を促進することにより、獲得免疫システムの抗炎症フォークを刺激します。 現在の社会を悩ませている炎症性疾患の現在の流行を考えると、それを予防し、戦うための実行可能で利用可能で安全な治療法が必要です. 炎症性および代謝性障害の病因は多因子性で非常に複雑ですが、最近の文献では、プロバイオティクスを主要な治療介入として使用して、腸内細菌叢と免疫応答を調節することが示唆されています。 その結果、安定した一貫した代謝環境を維持するための腸内細菌叢の調節は、影響を受けた患者の予防および追加治療に役立つ可能性があります。 将来的には、腸内微生物叢の恒常性を直接維持し、機能的代謝物(トリメチルアミン-N-オキシド、短鎖脂肪酸、胆汁酸、リポ多糖など)を調節する乳酸菌による治療アプローチを実証するために、はるかに詳細な臨床研究が必要になるでしょう。 これにより、免疫炎症、高脂質コレステロール、酸化ストレスに関連するリスクがさらに低下します。 著者の観点からすると、胃腸内微生物の複雑さを改善することによって全体的な健康を改善しようとしている人は、乳酸菌が豊富な発酵食(ヨーグルト、キムチ、味噌、ザワークラウトなど)を摂取することを目標とすることがより有益であることに気付くかもしれません. 結論として、乳酸桿菌による治療は、炎症性疾患の治療と予防における潜在的なフロンティアを提供します。 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| この文献は、Front Pharmacol. 2022; 13: 1042189.に掲載されたGut microbiome and human health: Exploring how the probiotic genus Lactobacillus modulate immune responses.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |