Kirsten Berding et al., |
概要 |
| 過去 10 年間で、腸内細菌叢は脳のプロセスと行動を調節する重要な要素として浮上してきました。食事は、生涯にわたって腸内細菌叢の組成を形作ることに関与する主要な要因の 1 つです。しかし、食事が微生物叢への影響を介して脳に影響を与える可能性があるかどうか、またどのように影響するかについては、注目を集め始めたばかりです.微生物代謝物、免疫、神経、代謝経路など、腸から脳へのコミュニケーションのいくつかのメカニズムが特定されており、そのうちのいくつかは食事の調節を受けやすい可能性があります。微生物叢 - 腸 - 脳の軸に対する栄養介入の可能性を調査する動物研究は、この双方向のコミュニケーションにおける食事の役割についての理解を深めました。このレビューでは、食事、微生物叢、および宿主の行動/脳プロセスを三角測量する文献の現状を要約し、潜在的なメカニズムについて説明します。さらに、食事介入に対する反応性の決定要因と、脳の健康に対する食事の効果の根底にあるモジュレーターとしての微生物叢の証拠について概説します。特に、この取り組みにおける全食事アプローチの使用が十分に研究されていないことと、臨床集団からのより多くの証拠が必要であることを強調します。有望な結果が報告されていますが、特に臨床コホートからの追加データは、脳と精神の健康を改善するための微生物叢を標的とした全食事戦略の開発に関する証拠に基づく推奨事項を提供するために必要です。 |
| 目次(クリックして記事にアクセスできます) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.微生物叢の組成と機能に対する食事の影響 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.1.腸内細菌叢とは? | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.2.食事と腸内細菌叢 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.3.多量栄養素 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.3.1.食物繊維 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.3.2.食事中の脂質と脂肪酸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.3.3.タンパク質とアミノ酸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.微量栄養素 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.1.ビタミンとミネラル | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.2.ポリフェノール | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.3.甘味料 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.4.乳化剤 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.食品群 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.1.果物と野菜 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.2.ナッツ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.3.豆類 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.4.発酵食品 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.6.全体の食事 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.6.1.地中海式食事 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.6.2.植物ベースの食事 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.6.3.西洋の食事 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.微生物叢が脳と行動に与える影響 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.人生の極限における食事微生物叢 - 腸 - 脳軸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.1.出生前の期間 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.2.幼児期 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.3.幼年期および思春期 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.4.高齢者 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.腸内細菌叢と行動を操作するための全食アプローチの使用 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.1.前臨床研究からの証拠 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.2.ヒト試験からの証拠 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.1.短鎖脂肪酸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.2.タンパク質分解による代謝物 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.3.トリプトファン代謝物 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.4胆汁酸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.2.免疫シグナル伝達 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.3.迷走神経と神経機能 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.4.ホルモン経路 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.5.代謝経路 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.6.視床下部-下垂体-副腎軸(HPA) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7.食事介入に対する応答者と非応答者 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.微生物叢は食事が脳と行動に及ぼす影響を調節していますか? | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9.結論と今後の方向性 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1.序章 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 人体には、何百もの異なる種に属する何兆もの微生物 [細菌、ウイルス、古細菌、下等および高等真核生物、菌類 (1) を含む] が生息しており、その大部分は腸内に存在します。 ここ数十年で、脳の健康を含む人間の健康のさまざまな側面に対する腸内微生物叢の影響に関する知識が指数関数的に増加しました (2)。 さらに、食事は腸内微生物叢の形成に関与する重要な要因の 1 つであり、特定の微生物の存在量と代謝能力だけでなく、微生物の多様性にも顕著な影響を与えることが明らかになりました (3–5)。 さらに、最適なメンタルヘルスをサポートする上での食習慣の役割がますます強調されています (6–8)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 最近、サイコバイオティクスという概念が登場し、微生物叢に影響を与える精神的健康に細菌を介したプラスの効果がある外因性要因 (例: プロバイオティクス、プレバイオティクス、食事による) (9–12)について説明します。 加工食品、揚げ物、砂糖が豊富な食品が豊富で、構成繊維とポリフェノールを含む植物性食品が少ない西洋式の食事の摂取は、重要な有益な微生物の絶滅と日和見病原体の拡大と同様に (13, 14),微生物の多様性と機能の喪失につながる可能性があり、人間の健康に多大な影響を及ぼすことは明らかです。 また、健康的な食事を使用して腸と脳のコミュニケーションを積極的に調整することは、一般的な精神障害の予防と治療の両方の可能性を秘めていることも認識されています (15)。 プレバイオティクス繊維が豊富な果物や野菜など、単一の食品の補給の影響に焦点を当てた新たな研究があり、マイクロバイオームと宿主の相互作用を調節する有望な結果が示されています (16)。 そのようなアプローチは、特定の食品が人間の微生物叢と健康にどのように影響し、新しい機能性食品の発見につながるかについての理解を深める上で重要ですが、人間は毎食ごとに食品群を組み合わせて摂取しており、単一の食品を研究することは、全体的な健康だけでなく、微生物叢の多様性と組成に対しても、食事成分が持つ潜在的な相乗効果を見逃す可能性があります (17)。 したがって、全食アプローチの研究は、新しい食事介入の開発へのより現実的な道を表し、国の健康的な食事のガイドラインと政策に情報を提供する可能性があります。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| この物語のレビューでは、食事、微生物叢、および宿主の行動/脳のプロセスを三角測量する文献の現状を要約します。 さらに、食事 - 微生物叢 - 脳の相互関係の根底にある潜在的なメカニズムが議論されています。 食事介入への反応の重要な決定要因として個人の微生物プロファイルを強調する最近の進歩もレビューされています。 この分野での知識の増加は、最終的には精神衛生への微生物叢を標的とした栄養アプローチの開発につながると考えられています。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.微生物叢の組成と機能に対する食事の影響 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.1.腸内細菌叢とは? | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| シーケンス技術とバイオインフォマティクスの進歩により、食事が微生物叢の構成に与える影響についての理解が深まりました (18, 19)。細菌は、分類学的に、門、綱、目、科、属、種、および株に分類されます。現在までに、25 の異なる門、約 2000 の属、および 5000 の種が特定されています (20)。 25の門の中で、最も優勢なものには、 Firmicutes, Bacteroidetes, Actinobacteria, Cyanobacteria, Fusobacteria, Proteobacteria, および Verrucomicrobiaが含まれ(21)、Bacteroidetes 門とFirmicutes門は、健康なヒトの腸内微生物叢全体の70〜90%を占めています(22). Firmicutes 門内の属には、Clostridium、Lactobacillus、Bacillus、Enterococcus、および Ruminococcus が含まれますが、Bacteroidetes 門は主に Bacteroides および Prevotella 属で構成されています。Bifidobacteriumは、 Actinobacteria門の主要な代表的な属です (23)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 人間の腸内には 1000 種以上の細菌が確認されていますが、人は平均して 160 種しか保有していません (24, 25)。 「健康な微生物叢」の詳細については論争が続いていますが、耐性 (摂動に抵抗する能力) と回復力 (ベースライン状態に戻る) によって定義できることが示唆されています (26)。同様に、微生物の豊富さ (微生物の数) と多様性 (異なる微生物の量、すなわち α 多様性) は、しばしば健康な微生物叢の関連マーカーです (27)。さらに、特定の細菌属は有益な共生生物と見なすことができます。つまり、それらは人間の宿主と相互に有益な関係で生きています。. 同時に、他の細菌属は潜在的な病原体として分類されており、これらの細菌の比率の不均衡は宿主の感受性を高める可能性があります. これは特定の宿主のコンテキスト内で異なる場合がありますが、bifidobacteria とlactobacilliは一般に「良い」細菌と見なされ、プロバイオティクスサプリメントで一般的に使用されていますが、Escherichia coli、Clostridium属内の菌株、リポポリサッカライド形成分類群Enterobacteriaceaeは病状と症状に関連しています (28–30)。 同様に、Firmicutes:Bacteroidetes 比として表される 2 つの支配的な門の間の関係は、いくつかの病理学的状態と関連付けられています (31、32) が、肥満との関連性はまだ議論されています (33)。健全な微生物叢を定義することの難しさを反映する 1 つの要因は、個人間で観察される高い変動性です。したがって、特定の微生物の存在に基づいて健康なマイクロバイオームを定義するのではなく、「機能コア」と呼ばれる重要な微生物機能の存在が、健康な微生物の状態を定義する上でより重要である可能性があることも示唆されています (4, 26)。これは、代謝機能が異なる微生物によって実行される可能性があることを意味するため、微生物叢の組成が異なる個体でも同じ微生物機能が発揮される可能性があります。同様に、人間の微生物叢の既存の未知数により、健康な微生物叢の定義が難しくなっています。過去 10 年間でシーケンシング技術は大幅に進歩しましたが、現在の微生物研究では、いくつかの分類群および株レベルの多様性と機能性が未調査のままです (20)。この菌株レベルの多様性は、特定の細菌属と健康または疾患との関連を決定する上で重要である可能性があり、これは Prevotella 属 (特に P. copri) 内での議論の焦点となっています (34)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.2.食事と腸内細菌叢 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 成人期の中核となる腸内微生物叢は比較的安定していますが、腸内微生物群集を形成する環境要因が特定されています (23、35、36)。 短期 (37) および長期 (3) の食習慣は、微生物の組成と多様性の要因の 1 つとして認識されており、微生物叢に対する個々の栄養素と食事パターンの両方の影響が広く調査されています。 腸内微生物群集に影響を与える食事要因を表 1 にまとめます。食事が微生物叢の構成に与える影響についてはいくつかの一般化を行うことができますが、最近の研究では、食事と微生物の相互作用は高度に個別化されており、ベースラインの微生物叢の存在に依存していることも示唆されています ( 110)、これは、個々のベースラインの微生物叢に合わせて食事介入を調整する必要があるかもしれないことを示しています(19)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.3.多量栄養素 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内微生物は、未消化の多量栄養素の消化、吸収、代謝、および変換に関与し、ヒト宿主にとって有益で生物活性のある化合物を抽出します。 微生物分類群の特殊な機能により、各主要栄養素は微生物プロファイルに異なる方法で影響を与えます。 多量栄養素の比率、量、および種類の変動は、微生物叢の組成に対する影響の大きな要因であり(111)、特定の微生物が選択的な多量栄養素で繁栄し、それによってそれらの量が増加します。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.3.1.食物繊維 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸内微生物叢を形成するための最も広く研究されている主要栄養素は、炭水化物、特に食物繊維です。 欧州連合規則 1169/2011 は、食物繊維を次のように定義しています。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 「ヒトの小腸で消化も吸収もされず、次のカテゴリーに属する、3 つ以上の単量体単位を含む炭水化物ポリマー: 食用炭水化物ポリマー (I) 消費される食品中に自然に存在するもの、および (II) 食品原料から得られるもの 一般に受け入れられている科学的証拠によって実証された有益な生理学的効果を有する物理的、酵素的または化学的手段、または(III)一般的に受け入れられている科学的証拠によって実証された有益な生理学的効果を有する食用の合成炭水化物ポリマー」(112)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| よく研究されている別の種類の食物繊維はプレバイオティクスで、これは「健康上の利益をもたらす宿主微生物によって選択的に利用される基質」として定義されています (113)。 ほとんどのプレバイオティクスは食物繊維として分類できますが、すべての食物繊維がプレバイオティクスであるとは限らないことに注意することが重要です。 プレバイオティクスは一般的に発酵性がありますが、これはすべての食物繊維に当てはまるわけではありません。 プレバイオティクスの例には、ペクチン、イヌリン、フラクトオリゴ糖、およびガラクトオリゴ糖が含まれます。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 高繊維食の摂取が細菌の多様性の増加を促進し、有益な細菌(すなわちBifidobacterium sp. Lactobacillus sp., Akkermansia sp., Faecalibacterium sp., Roseburia sp., Bacteroides sp., および Prevotella)の増殖を促進し、潜在的な病原菌 (Enterobacteriaceaeなど) の減少につながることは一般に認められています (63–70, 114. –116)。 より具体的には、さまざまな繊維の化学的性質 (重合、溶解性、粘度など) が消化管内の代謝の場所を決定し、摂取に応じて特定の微生物の変化を引き起こします。例えば、補給研究では、β-グルカン (可溶性非デンプン多糖類) を含む全粒製品がヒト (71) とラット (72) の lactobacilli とbifidobacteriaの増殖をサポートするのに対し、無傷の穀物繊維 (例えば、全粒穀物、大麦繊維、小麦) はふすま、およびライ麦繊維) は、ヒトのActinobacteria, Bifidobacterium, Clostridium, Lachnospira, Akkermansia,および Roseburia の存在量を増加させます (63、65、66、73)。 人間が難消化性デンプンを摂取すると、Bifidobacterium, Faecalibacterium, および Eubacterium,が大幅に増加し、一部のRuminococcus株が減少しました (74, 75)。 繊維の溶解度も、微生物プロファイルへの影響を決定します。不溶性繊維と比較して、可溶性繊維は子豚モデルの微生物組成と多様性により顕著な影響を与えるようでした (76)。 それにもかかわらず、果物や野菜の主要な繊維源であるセルロースなどの不溶性、非発酵性繊維は、セルロース分解微生物(Ruminococcus や Fibrobacterなど)によって代謝される可能性があります。 可溶化された製品(オリゴ糖や多糖類など)をクロスフィードすることで、細菌の量だけでなく、それらの量にも影響を与えます(117)。 動物実験では、セルロースは微生物の豊富さを増加させることが示されました (77)。 そして微生物叢の構成を変更し、Peptostreptococcaceae, Clostridiaceae, Akkermansia, Parabacteroides, Lactobacillus, Clostridium, Eisenbergiella, Marvinbryantia, Romboutsia, Helicobacter, Enterococcus, あるいは Desulfovibrio (77–79) の量を増やし、Sutterellaceae, Lactobacillaceae, あるいは Coriobacteriaceaeを減らします (77 、79)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 微生物組成の変化に加えて、さまざまな食物繊維も微生物の酵素能力と代謝産物濃度に影響を与えます。 溶解度や発酵性などの化学的性質は、微生物発酵の程度と場所、および生成される代謝産物のタイプを決定します(76)。 可溶性の発酵性繊維は、複雑な炭水化物を分解し、健康を促進する短鎖脂肪酸、すなわち酢酸、プロピオン酸、酪酸を生成する微生物の酵素能力を高めることができます (114, 118)。 短鎖脂肪酸、特に酪酸は、胃腸(結腸細胞の主なエネルギー源であり、腸のバリア機能をサポートする)および代謝(グルコース恒常性、脂質酸化)の健康に関与しており、抗炎症および免疫調節特性を発揮し、中枢機能に影響を与える可能性があります( 詳細は後述) (119, 120)。 ヒトでの多数の介入研究は、炭水化物と全粒穀物の消費を減らすと、プロバイオティクスのbifidobacteriaを含む重要な酪酸産生菌や 短鎖脂肪酸自体の存在量が減少することを示しています (121–123)。 不溶性食物繊維は短鎖脂肪酸産生に顕著な影響を与えませんが、リノール酸、ニコチン酸、ニコチンアミド、グリセロリン脂質、グルタチオン、スフィンゴ脂質の経路、およびバリン、ロイシン、イソロイシンの代謝経路の変化が、不溶性繊維 (セルロースなど) の摂取に反応して観察された (78, 79)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.3.2.食事中の脂質と脂肪酸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ほとんどの脂肪酸は小腸で吸収されますが、食事の脂質と脂肪も微生物プロファイルに顕著な影響を示します。これらの変化が有益か有害かは、脂肪の種類によって異なります。さまざまな程度の飽和度が、微生物組成を異なる形で形成することが報告されています。 たとえば、飽和脂肪酸(SFA)の摂取量が多いと、ヒトの総細菌量(95) および微生物の多様性と豊富さの減少(95, 96) 、マウスにおける炎症誘発性細菌 (Bilophila wadsworthia など) の増加 (96–98) に関連することが示されています。 ヒトでは、より健康的な多価不飽和脂肪酸 (オメガ-3 PUFA など) が、 Lachnospiraceae および Ruminococcaceaeの分類群と同様に (82) より高い微生物多様性と相関しているLactobacillus, Lachnospira, Roseburia, and Bifidobacteriumなどの酪酸生産菌を含む有益な細菌の増殖を促進します (81)。 飽和度に加えて、鎖長も腸内微生物叢に対する脂肪酸の影響を決定します。動物研究の結果は、中鎖脂肪酸 (炭素数 7 ~ 12) がBifidobacterium, Bacteroides, および Prevotella の存在量を増加させClostridium histolyticum または Helicobacterの存在量を減少させることを示しています (119, 124-126)。一方、長鎖脂肪酸 (炭素数 13 ~ 18) は、動物モデルでBlautia, Clostridium, Coprococcus, Dialister, Lactococcus, Roseburia, または Bacteroides (119、127 ~ 129) の存在量を変化させます。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 中枢機能に対するω-3 (n–3) 脂肪酸の利点は、記憶力、気分、注意力、および認知能力の向上から、うつ病の発症リスクの低減およびストレス感受性の調節にまで及びます (131–138)。 これらの利点のほとんどは、脳の膜構造、機能およびシグナル伝達、神経伝達物質代謝回転の調節、神経新生、または抗炎症および抗アポトーシス効果への多価不飽和脂肪酸の関与に関連している可能性があります (139)。 多価不飽和脂肪酸がプレバイオティクスと見なされる可能性があるという考え (140) は、微生物の変化による別の間接的なメカニズムを示唆しています。 実際、ω-3 脂肪酸は、有益なbifidobacteriaを増加させ、enterobacteriaを減少させることにより、病理学的状態で真正生物の状態を回復することが提案されており、短鎖脂肪酸 の産生とエンドトキシン血症の抑制を通じて抗炎症環境をサポートします (81, 141)。 . このカスケードのイベントは、特にうつ病などの炎症関連障害において、脳と行動に上流の影響を与える可能性があります. 実際、動物モデルでは、ω-3脂肪酸の補給に関連する特定の微生物の変化(例えば、Lactobacillusと bifidobacteriaの存在量の増加、enterobacteriaに対するbifidobacteriaの比率の変化)は、行動の変化と密接に関連していました(142)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.3.3.タンパク質とアミノ酸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 食物タンパク質の供給源、濃度、およびアミノ酸バランスは、腸内微生物の組成、構造、および機能に影響を与える主な要因です。 人間の介入研究では、動物ベースのタンパク質は、植物ベースのタンパク質よりも微生物叢の組成に対してより顕著な効果を引き出します (37)。 マウスでは、動物由来のタンパク質は潜在的に有害な腸内微生物、例えば、Peptostreptococcaceae, Ruminococcaceae, Enterococcus, Streptococcus, Turicibacter, または Escherichiaを増加させます (92), 植物ベースのタンパク質は、 Bifidobacterium, Roseburia, および Lactobacillusの量を増やし、Bacteroides fragilis やClostridium perfringensなどの病原体の量を減らします (37, 42, 80)。 より具体的には、動物性タンパク質のさまざまな供給源は、特定の細菌の相対的存在量の明確な変化と関連しています。 たとえば、ラットとヒトの糞便接種物を用いた in vitro 研究を使用して、鶏肉タンパク質はActinobacteria, Bifidobacterium, および Bacteroides,を増加させることができるのに対し、牛肉タンパク質はProteobacteria および Oscillibacterのレベルの上昇に関連し、C. perfringens と C. histolyticum を減少させることが実証されました(93、94)。 さらに、さまざまな量のタンパク質摂取量は、微生物量にさまざまな影響を与えます. 子豚モデルでは、食事中のタンパク質濃度の低下は細菌の豊富さと Clostridium_sensu_stricto_1 の豊富さの減少をもたらしましたが、Escherichia–Shigellaの豊富さは増加し、適度なタンパク質制限はPeptostreptococcaceaeの上昇と関連していました (143)。 最後に、微生物の代謝産物は、消費されるタンパク質の量によっても影響を受けます。 高タンパク、低炭水化物の食事に切り替えると、酪酸を生成する細菌の量が減少し、結腸のタンパク質発酵と、分枝鎖脂肪酸 (BCFA) やフェニル酢酸および N-ニトロソ化合物の濃度などの健康に有害な代謝物が増加します(121)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.微量栄養素 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.1.ビタミンとミネラル | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ビタミンとミネラルは、神経伝達物質の合成と代謝、およびニューロンのエネルギー代謝における重要な補因子です。 腸内微生物叢が特定のビタミン、特にビタミン K およびビタミン B 群 [例えば、コバラミン (B12)、葉酸、およびリボフラビン (144, 145)] を合成できることはよく理解されており、その一部は直接吸収される可能性があります。 ビタミンとミネラルは主に上部消化管で吸収され、通常は少量しか結腸に到達しないため (146)、ヒトの結腸微生物叢に対するこれらの栄養素の影響を研究することは困難であり、一貫性のない結果が報告されています (147)。 それにもかかわらず、現在、遠位結腸に到達するビタミンが常在微生物の重要な栄養源として機能できるという証拠が蓄積されています(148)。 腸内細菌叢に対するビタミンDの影響に関する人間と動物の研究から得られた証拠を要約した最近の系統的レビューは、ビタミンDの状態または補給が微生物叢の組成を調節できることを示唆しています。しかし、特に人間の研究からの一貫性のないデータにより、傾向はまだ現れていません (147)。 文献の現状では、特定の分類群に対するビタミンの影響について結論を出すことはできませんが、それらの双方向の関係は、共生生物の豊富さと腸全体の恒常性を維持する上で重要な役割を果たすことが示唆されています (149–151)。例えば、ビタミンDおよびAと微生物叢との間の相乗効果は、免疫機能の調節および腸のバリア機能の維持において重要である可能性があります(150, 152)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ビタミンと同様に、ミネラルと微量元素は共生関係で腸内微生物叢と活発に相互作用します (153)。 多くの腸内細菌は成長と生存のためにミネラルを必要とするため(154)、一部のミネラルの欠乏と過剰は、微生物の不均衡と病原性微生物の増殖と適応度の増加に関連しています(155)。 例えば、ケニアの子供たちのコホートにおける鉄補給は、Bacillus cereus, Staphylococcus aureus, Clostridium difficile, C. perfringens,およびSalmonella,などの病原体の存在量を増加させ、潜在的に腸の炎症に寄与した(156)。 一方、他の研究では、過剰なミネラルがヒトの微生物叢の組成に有益な影響を与えるか、影響を与えないことが示されています(157)。 したがって、特に健康な集団において、さまざまなミネラル状態が微生物叢に与える影響を解読するには、さらに多くの研究が必要です。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.2.ポリフェノール | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ポリフェノールの植物化学的クラスには、フラボノイド (すなわち、フラバノン、イソフラボン、アントシアニジン) と非フラボノイド (すなわち、スチルベン、リグナン、およびタンニン) が広く含まれます。 ポリフェノールが豊富な食品には、果物や野菜、ココア、スパイス、全粒穀物、ナッツ、エキストラバージン オリーブ オイル、赤ワイン、コーヒー、緑茶などの飲み物が含まれます (158)。 ポリフェノールの約 90 ~ 95% は吸収されないため、腸内微生物によって分解される可能性があります (83)。 ポリフェノールの摂取には、主に抗炎症作用と抗酸化作用による神経保護効果 (159, 160)、高齢者集団の認知能力の改善 (161) および健康な若年成人 (162) など、多くの健康上の利点が関連しています。動物モデルでは、コルチコステロンと炎症誘発性サイトカインの放出が減弱し、抑うつ様行動が緩和されました(163)。 新しい観察研究では、ポリフェノールの食事摂取量の増加とうつ病の発生率の低下との関連性が報告されています (164, 165)。たとえば、Nurses' Health Study (n = 82,643 女性) の前向き分析では、柑橘類に含まれるポリフェノールの総クラスとサブクラスは、うつ病の発生率の低下と関連していると報告されています (164)。若年期ストレスに関する最近の動物研究では、特に微生物叢-腸-脳軸経路に関連する微生物において、微生物の多様性と組成を調節しながら、抑うつおよび不安様行動の改善とコルチコステロンレベルの低下も示されました(166)。 したがって、マイクロバイオームの変化は、ポリフェノールが精神的健康を改善する根本的なメカニズムである可能性があります. 最近の人間による介入研究では、フラボノイドが豊富なオレンジ ジュースは、フラボノイドが少ないオレンジ コーディアルと、 Lachnospiraceae, Bifidobacteriaceae, および Bacteroidaceaeに属する豊富な細菌属と比較して、抑うつ症状を大幅に軽減しました (167)。 興味深いことに、Lachnospiraceae_uc の存在量は、血清脳由来神経栄養因子 (BDNF) 濃度 (大うつ病性障害の患者ではしばしば減少する) と正の相関があり、フラボノイド治療後に増加したうつ病コホートにおける抗うつ薬の有効性に寄与しました(167)。 したがって、フラボノイド消費量の増加がLachnospiraceae_ucの存在量を上昇させ、それが血清脳由来神経栄養因子レベルを回復させ、抑うつ症状を軽減したことが示唆される可能性があります。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 微生物群集に対するポリフェノールの影響をテーマにした膨大な数の動物および in vitro 文献が利用可能です (4)。 ヒトコホートから入手できるデータは少なく、ほとんどの研究は、個々のフェノール化合物ではなく、ポリフェノールを含む食品全体の影響を調査しています (4)。 ポリフェノールは「プレバイオティクスのような」効果を示し、bifidobacteria や lactobacilli,などの有益な細菌株の増殖を増加させる一方で、C. perfringens や C. histolyticumなどの潜在的な病原体の数を用量依存的に減らします (83, 84)。 いくつかの一般的な細菌の変化はポリフェノールで観察できますが、特定の微生物が特定のフェノール化合物と関連している場合もあります。 Bacteroides, Clostridium, および Staphylococcus属の種は、カカオ (主にフラボノール) の摂取に伴って減少し、 Prevotella group, Blautia および Faecalibacterium prausnitziiは増加することが報告された (85) のに対し、コーヒー ポリフェノール [すなわち、フェノール酸 (クロロゲン酸)] は、Bacteroidesの存在量と直接関係があり、観察研究では、コーヒーの摂取量が多いと、Bacteroides–Prevotella–Porphyromonasのレベルが高くなります (86)。 赤ワイン (特にスチルベンの一種であるレスベラ トロールが豊富) は、α 多様性の増加と、Barnesiella, Phascolarctobacterium, およびPrevotellaceae__NK3B31 の豊富さと関連していました (87)。 ウコンに含まれる親油性ポリフェノールであるクルクミンの微生物代謝は、抗酸化、抗炎症、神経保護の特性を持つ代謝産物をもたらし、有益な細菌株を促進します (168)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.3.甘味料 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 人工甘味料(アスパルテームやサッカリンなど)および天然甘味料(ステビアなど)は、現在、食品に含まれる砂糖の量を減らすために、食品業界で一般的に使用されています。腸内細菌叢に対する食事の影響が知られているため、甘味料の摂取が腸内細菌叢の構成に及ぼす影響を調査する研究が増えています (詳細なレビューについては、169、170 を参照してください)。以前の人間の研究では、微生物の多様性と組成に対する有害な影響が示されていましたが (99, 100)、より最近の研究では、非栄養甘味料の摂取を「微生物叢の研究でますます冗長な用語 (171)」および酪酸濃度の減少に関連付けました。 (172)、一部の報告では、微生物叢に対する甘味料の明確な影響は確立されていない(170)、または一部の甘味料(サッカリン、スクラロース、ステビアなど)のみが微生物プロファイルに影響を与える(173)と結論付けています。したがって、微生物叢に対する特定の影響はまだ解明されておらず、さまざまな甘味料の化学的属性と結腸に到達する濃度に依存している可能性が最も高い(169)。例えば、アスパルテームとサッカリンは大部分が消化管の上部で分解されて吸収されるのに対し、スクラロースの 85% は結腸に到達し、ステビオール配糖体 (すなわちステビア) は無傷で結腸に到達し、細菌の代謝を必要とすると推定されています ( 169)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| スクラロース (すなわち、スプレンダ) の投与は、特に動物モデルで微生物の変化を誘発することが報告されています (例えば、総細菌量の減少; Firmicutes, Proteobacteria, Turicibacter, Roseburia, Akkermansia, Clostridiaceae, Christensenellaceae, および Clostridium symbiosiumの増加; Ruminococcus, Streptococcus, Dehalobacterium, Erysipelotrichaceae, および bifidobacteriaの減少)(101–103)。 興味深いことに、これらの微生物の変化のいくつかは、耐糖能異常 (100) や慢性炎症 (101) など、甘味料の消費に関連する健康への悪影響を引き起こすことが提案されています。 しかし、健康なボランティアを対象とした小規模な人間の研究では、スクラロースの短期摂取は腸内微生物叢の組成に大きな変化を誘発しませんでした (104)。 これらの不一致の一部は、微生物叢によるスクラロースの代謝の制限 (174) と、スクラロースの投与量の違い、およびスクラロースへの曝露期間に起因する可能性があります。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 抗菌性および抗真菌性を有し、主にBacteroidesの微生物酵素によって代謝されるにもかかわらず (175, 176)、限られた数の研究のみが、ステビアまたはその主要な化学化合物であるステビオール配糖体の腸内微生物叢への影響を調査しています。 . in vitro 研究では、ステビオール グルコシドが Lactobacillus reuteri などのプロバイオティック バクテリアの増殖を阻害する可能性があることを示唆していますが (105)、E. coliなどの病原体に対しても静菌効果を発揮する可能性があります (177)。 しかし、別の in vitro 実験では、ステビオール配糖体とのインキュベーション後に多様性や組成に変化は見られませんでした (106)。 人間を対象とした試験からの証拠は不足していますが、最近のラットの研究では、低用量のステビア (レバウディオサイド A) を幼少期から 9 週間にわたって摂取すると、Bifidobacteriaceae とLactobacillus intestinalisのメンバーが減少し、Bacteroides thetaiotaomicron と Akkermansia muciniphilaの存在量が増加することが示されました。 興味深いことに、ステビアの介入は、側坐核で輸送されるチロシンヒドロキシラーゼとドーパミンの減少によって証明されるように、中脳辺縁系の報酬システムを通じて食欲行動にも影響を与えるようでした(178)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.4.4.乳化剤 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 乳化剤 [カルボキシメチルセルロース (CMC)、ポリソルベート-80 (P80)、アラビノガラクタン、カラギーナン] は食品添加物で、西洋の食事で広く普及しており、食品の風味を変えたり、食感、安定性、貯蔵寿命を改善するために一般的に使用されています。 宿主の生理機能と腸内微生物叢の両方に対する乳化剤のほとんどの悪影響は、動物モデルを使用して実証されており(179)、乳化剤によって誘発される微生物の変化が、肥満、メタボリックシンドローム、腸の炎症、潜在的に病原体の移行を促進することによる結腸癌などの慢性炎症性疾患に寄与する可能性があることさえ示唆されています(107, 108)。 重要なことに、無菌 (GF) 動物 (107) および高度に制限された微生物叢を持つ動物 (180) による乳化剤の消費は、同じ有害な健康影響を誘発しないことが示されており、 宿主の健康に対する乳化剤の悪影響には、微生物の調整が必要な場合があります。 Porphyromonadaceae, Helicobacter, Campylobacter jejuni, Salmonella, および Clostridium cluster XI の増加、Bacteroidesの存在量の減少など、いくつかの特定の、潜在的に性依存的な微生物の変化が、マウスの乳化剤消費に関連している(181)。 メスのマウスでは、CMC はAnaeroplasmaを増加させ、P80 はProteobacteria, Clostridium, および Burkholderia,の相対的存在量を増加させたのに対し、オスのマウスでは、CMC はDoreaの存在量を増加させ、P80 処理はBacteroides, Burkholderia, Clostridium, および Veillonellaの存在量を増加させた(109)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.食品群 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.1.果物と野菜 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 発酵食品、果物や野菜、ナッツなど、個々の食品グループが腸内微生物叢に与える影響も研究対象となっています。最近、個々の果物や野菜が腸内微生物叢に及ぼす影響の詳細なレビューが提供されました (16)。 いくつかの人間と動物の研究は、果物と野菜の摂取が微生物の多様性と機能の増加、細菌門の豊富さの変化、Bifidobacterium や Lactobacillusなどの有益な細菌の成長と、E. coli や Enterococcus などの潜在的に有害な細菌の減少につながることを示しています(51–55)。 これらの利点の一部は、オリゴ糖、ペクチン、セルロース、イヌリン、リグナン、難消化性デンプンなどのいわゆる微生物叢アクセス可能な炭水化物 (MAC)、および特定の細菌によって生体内変換され、阻害する可能性のあるポリフェノールに関連している可能性があります。ポリフェノール (16) で詳細に説明されているように、病原性細菌の増殖を抑制し、有益な細菌を刺激します。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.2.ナッツ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 同様に、植物ベースの食事や地中海式の食事で一般的に消費されるナッツには、繊維、不飽和脂肪酸 (PUFA など) などの栄養素が豊富に含まれています。 生物活性化合物 [例えば、抗酸化物質 (トコフェロール)、ポリフェノール、フィトステロール] は、微生物叢の構成に潜在的なプレバイオティクス効果をもたらします (60, 182, 183)。 腸内細菌叢に対するナッツ摂取の影響に関するランダム化比較試験の最近の系統的レビューとメタアナリシスは、ナッツの摂取が属レベルで微生物叢を形成すると結論付けました(例えば、Clostridium, Dialister, Roseburia, およびLachnospira,増加、Parabacteroidesの減少)。 ;ただし、特定の効果は、消費されたナッツの種類と量、および介入の期間によって異なります (184)。例えば、ある無作為化対照クロスオーバー研究では、健康なボランティアが毎日42gのクルミを3週間摂取すると、一部の酪酸産生菌(例えば、Faecalibacterium やRoseburia)、Clostridium や Dialisterを含むFirmicutes属の相対存在量が増加しました(61)。一方、若年成人に 56.7 g のアーモンドを 8 週間摂取させると、α 多様性測定値が増加し、B. fragilis の存在量が減少することが明らかになりました (62)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.3.豆類 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 豆、レンズ豆、ひよこ豆などの豆類は、マメ科植物の食用種子です。 この食品グループは、植物ベースの食事のタンパク質源として役立つことが多く、葉酸、鉄、多価不飽和脂肪酸 (PUFA) /一価不飽和脂肪酸 (MUFA) 、特定の植物化学物質、食物繊維も豊富です。. 豆類のこの栄養成分は、腸内微生物叢の組成と代謝産物の生産の変化にも関連していました。最近のシステマティックレビューでは、豆類消費後に有意な変化が観察される可能性があるが、特にヒトでは結果に一貫性がないと結論付けられている(185)。 たとえば、Bifidobacterium sp. と Lactobacillus casei/L. bifermentum sp.のパーセンテージが高くなり、Clostridium cluster XI および I/II のパーセンテージが低いことは、ヒトのひよこ豆の摂取と関連していた (186)。 一方、うずら豆は代謝前症候群の集団では最小限の効果しかなく、Eubacterium limosumの存在量を低下させるだけでした (187)。 動物モデルでは、抽出された豆類の粉でより顕著な変化が観察されました。 マウスでは、基礎食と比較して、白インゲン豆と黒インゲン豆の粉は Prevotella, S24–7, および Ruminococcus flavefaciensの存在量と 短鎖脂肪酸産生を増加させ、Ruminococcus gnavus、Oscillospira、Coprococcus、Lactococcus、Streptococcus、Coprobacillus、Parabacteroides、Adlercreutzia などの存在量を減少させました。。 いくつかの豆特有の変化も観察され、黒インゲン豆の粉は α 多様性を増加させ、白インゲン豆の粉は潜在的な病原体 C. perfringens の存在量を減少させた(188)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.5.4.発酵食品 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 発酵食品(ザワークラウト、キムチ、ケフィア、乾燥発酵ソーセージ、ヨーグルト、チーズ、コンブチャ、味噌など)は、制御された微生物の増殖を通じて生産された食品または飲料として定義され、両方のプロバイオティック微生物叢(最も一般的にはLactobacillus, Streptococcus, Lactococcus, および Leuconostoc)および酵母ならびに微生物代謝産物を含み(189、190)、何世紀にもわたって人間によって消費されてきました。しかし、それらの人気は最近急上昇しており、宿主の微生物叢やメンタルヘルスを含む健康への影響についての新しい調査につながっています (191, 192)。 当然のことながら、「生きた」発酵食品を摂取すると、食事中の微生物数が 10,000 倍に増加し、腸内微生物プロファイルを調節する可能性があります (193, 194)。例えば、肥満女性のキムチ摂取後にBifidobacterium存在量の有意な増加が観察され(56)、発酵豆乳はcoliform(大腸菌)群とC. perfringensの減少、 Bifidobacterium と Lactobacillusの増加をもたらした(57)。 最近のマウス研究では、ケフィアの投与により、L. reuteri, Eubacterium plexicaudatum, およびBifidobacterium pseudolongum,の存在量が増加し、 Lachnospiraceae bacterium 3_1_46FAA, Propionibacterium acnes, およびBacillus amyloliquefaciensが減少しました。 そして、腸内微生物叢の機能的可能性を神経活性代謝産物の産生に向けてシフトしました(58)。 別の最近の人体研究では、発酵乳飲料を 4 週間摂取すると、いくつかの特定の属 (例えば、Holdemania, Gordonibacter, Lactobacillus, 未分類の Mollicutes (RF-9), および Clostridiales の 2 つの未分類の属) の存在量が増加し、常在微生物のいくつかの機能を強化した(195)。 これらの有望な結果にもかかわらず、最近の文献レビューは、特定の発酵食品に関連する特定の微生物パターンを推測するのに十分なデータがまだ利用できないと結論付けました(196)。 この不一致は、予測が困難な方法での発酵製品間の微生物組成の変動に起因する可能性があります。 それにもかかわらず、発酵食品が常在微生物と健康転帰に与える影響をさらに解明するには、より大規模な臨床試験が必要です(196)。 この取り組みにおいて、American Gut Project の 115 人の個人からのサンプルを分析した別の最近の研究では、発酵植物を週に 1 ~ 2 回、または 1 日 1 回摂取した人々は、用量依存的で、β 多様性によって測定された腸内コミュニティは、非消費者と比較して有意に異なる腸内コミュニティを持っていたことが示されました。 さらに、発酵食品の消費と微生物の機能プロファイルとして豊富な細菌分類群(例えば、Bacteroides, Pseudomonas, Dorea, Prevotella, Oscillospira, F. prausnitzii, Lactobacillus spp.)の関連性が報告されています (59)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.6.全体の食事 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 単一の栄養素が微生物叢の組成に与える影響を理解することで、食事と微生物叢の相互作用に関する理解が大幅に進歩しましたが、食事は全体として考慮する必要があることが示唆されています。 これは、一般的な食物消費パターンをより反映しており、微生物叢の組成に対する栄養素の相互作用による潜在的な相乗効果または相加効果を考慮しています (197)。 したがって、特定の食事パターンに関連する微生物叢のプロファイルを作成する研究が開始されており、以下で概説されています。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.6.1.地中海式食事 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 果物、野菜、豆類、ナッツ、オリーブオイル、魚の摂取量が多く、赤身肉、乳製品、飽和脂肪の摂取量が少ないことを特徴とする地中海式ダイエット (198) は、以下を含むさまざまな健康上の利点があることでよく知られています。メンタルヘルスと認知 (199–202)。 より最近の人間による介入研究も、微生物叢プロファイルに対する地中海式食事の有益な影響を支持しています。 微生物の多様性と健康を促進する細菌分類群 (Clostridium cluster XIVa, F. prausnitzii, Roseburia, Eubacterium, B. thetaiotaomicron, Parabacteroides distasonis, Bifidobacterium adolescentis, およびBifidobacterium longum) の豊富さは地中海食の消費と関連しています(38–40)。 さらに、地中海式食事の順守は、短鎖脂肪酸レベルの上昇や分枝鎖脂肪酸、胆汁酸、トリメチルアミン N-オキシド (TMAO) の減少など、有益な微生物叢関連のメタボローム プロファイルと関連していました (38, 40, 41)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.6.2.植物ベースの食事 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ベジタリアンやビーガンの食事を含む植物ベースの食事は、果物、野菜、豆類、ナッツ、種子が豊富な食事パターンです。魚介類が含まれる場合がありますが、肉、卵、乳製品などの動物性製品は含まれていません。 特定の微生物組成は植物ベースの食事への順守の程度に依存しますが、一般的に好ましい微生物パターンが観察されています。重要な証拠は、ブルキナファソに住む子供たち (主に菜食主義者の食事) の微生物叢の組成を、典型的な西洋の食事を消費するイタリアに住む子供たちと比較した以前の研究によって具体的に提供されました (43)。 ブルキナファソに住む子供たちでは、微生物の豊富さと生物多様性が観察されました。より具体的には、これらの子供たちは、難消化性多糖代謝に特化した微生物プロファイルを持ち、Bacteroidetes と Actinobacteriが豊富で、イタリアの子供と比較して、Prevotella, Xylanibacter, とTreponemaが微生物叢に独占的に含まれていました。 一方、Firmicutes と Proteobacteria は、Enterobacteriaceae (すなわち Shigella と Escherichia) の過剰表現で、イタリアの子供でより豊富でした。 成人では、アフリカの農村部に住む人々の微生物叢と米国に住むアフリカ系アメリカ人の微生物叢を比較した研究でも、アフリカの農村部での主に菜食主義者の食事は、Prevotella、Succinivibrio、および Oscillospira の優位性と関連し、総量と酪酸が増加することが明らかになりました。アフリカ系アメリカ人の微生物叢は、Escherichia とAcinetobacterなどの潜在的な病原菌に富んでいた(44)。 これらの違いは他の環境要因にも起因する可能性がありますが、菜食主義者と非菜食主義者を比較した研究では、同様の微生物プロファイルが明らかになりました. したがって、これらのタイプの食事の腸内微生物叢の特徴的なパターンには、細菌の豊富さ、Bifidobacterium, Lactobacillus, Ruminococcus, Eubacterium rectale, Roseburia, Prevotella, F. prausnitzii, および Anaerostipes の数の増加が含まれます。 しかし、Clostridium sensu stricto、C. perfringens、C histolyticum、および Odoribacter の量が少ない (42、45–49)。 それにもかかわらず、動物ベースの食事と比較した植物ベースの食事が微生物叢に及ぼす影響に関するデータは 微生物組成がビーガンと雑食動物の間でわずかに異なるだけであることを示す人間の研究によって証明されているようにまだ決定的ではありません(110)。 ただし、細菌のメタボロームにはかなりの変動が観察され、基質としての食事が微生物の組成自体よりも微生物の代謝産物の産生を決定する上で大きな役割を果たしている可能性があることを示唆しています。 実際、人間の雑食性、菜食主義者、または完全菜食主義者の食事は、タンパク質と代謝産物の微分的な微生物合成に関連しています。 菜食主義者と完全菜食主義者の食事は、腫瘍の抑制に関与する高レベルの酵素と関連していましたが、雑食動物は、フェノールやインドール誘導体、トリメチルアミン N-オキシドなどの有害な微生物代謝産物のレベルが最も高かった (50)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2.6.3.西洋の食事 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 飽和脂肪、動物性タンパク質、および精製された炭水化物が多いが、食物繊維の量が不十分な西洋の雑食タイプの食事は、微生物叢の組成をより病気に関連したタイプにシフトさせることが一般に認められています (203)。ヒトや動物が長期的に西洋食を摂取すると、有益な微生物が絶滅し、細菌の多様性が減少し、微生物叢が優勢なBacteroides主導型のエンテロタイプになる可能性があります (13, 14, 116, 88)。 同様に、Firmicutes と Bacteroidetes の比率の増加 (41) と、保護的な 短鎖脂肪酸産生細菌 (F. prausnitzii など) の減少 (89, 204) がしばしば観察されます。最近では、ヒトを対象とした小規模なパイロット クロスオーバー研究で、Collinsella, Parabacteroides, や B. wadsworthia,などの胆汁耐性微生物が増加し、西洋式の食事を採用してから 4 日以内に腸内微生物叢と関連する代謝物に有害な影響が及ぶことが示されました。 トリメチルアミン N-オキシド が増加し、代謝物であるインドール-3-乳酸とインドール-3-プロピオン酸が減少する (205)。 西洋の食事と同様に、動物実験における高脂肪食は、腸内微生物群集の構造を再現可能に変化させ、全体的な微生物叢の多様性と有益な細菌 (A. muciniphila, Bifidobacterium, Lactobacillus, およびLactococcusなど) を減少させます。 Firmicutes:Bacteroidetes の比率と、 Enterobacteriales, Clostridium cluster XVIa, Mollicutes, およびB. wadsworthia の量を増やします (90、91、206)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 超加工食品(砂糖入り飲料、スナック、ファーストフード)は、西洋の食事の特徴であり、すぐに摂取できる処方であり、精製された物質から作られ、カロリーが高く、飽和脂肪が豊富で、単糖、塩、その他の添加物が加えられています。添加物;それらは、うつ病を含む健康状態の悪化と一貫して関連しています(207)。これらの食品成分は微生物叢に有害であり、いくつかの研究では、超加工食品の消費が腸内微生物組成に与える影響が報告されています (208)。 例えば、Dialist、Coprococcus、Megasphaera、Oscillospira、および Blautia obeum の豊富さは、人間の加工食品の摂取に関連して最も豊富であるように思われました (209)。超加工食品の消費と微生物叢の構成との関連が報告されているため、これらの超加工食品を消費する子供の増加と、微生物叢を含む子供の発達への影響による懸念も高まっています。 たとえば、 Lachnospiraceae 属( Clostridium clostridioforme, C. bolteae, C. celerecrescens, またはC. sphenoidesに関連する), Ruminococcus, and Bacteroidesは、加工食品グループ (加工肉やおいしいスナックなど) と負の関連がありました。 一方、Lachnospiraceae (Fusicatenibacter saccharivorans), Blautia, および Clostridiumは、子供の加工食品の消費と正の関連がありました (210)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3.微生物叢が脳と行動に与える影響 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 過去数十年間、微生物叢は、微生物叢 - 腸 - 脳軸と呼ばれる双方向のコミュニケーションを介して、脳のプロセスと行動を調節する重要な役割を果たしてきました (2)。特に、異常な行動と神経化学的プロファイルを示した 無菌動物を使用した研究は、腸内微生物叢と脳の発達とプロセスの間の関連性を確立する上で重要でした (211–217)。さらに、微生物叢は、動物モデルのストレス反応を調節する視床下部-下垂体-副腎 (HPA) 軸の発達と同様に、正常な神経発達において多くの重要な役割を果たしています (218, 219)。人間の腸内微生物叢と脳機能との間の直接的なつながりを確立することは、より困難であり、相関関係があります。最近の人間の脳画像研究では、健忘性軽度認知障害患者の腸内微生物叢組成と脳活動パターンとの間に相関関係が見られました(220)。さらに、特定の細菌分類群の存在量と、自閉症スペクトラム障害 (ASD) などの疾患の症状との間に相関関係が確立されています (221)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 微生物叢と脳の関係に関する追加の証拠は、有益な微生物であるプロバイオティクスの直接投与を宿主の行動および認知の変化に関連付ける動物と人間の両方の研究から得られています。動物実験では、プロバイオティクス (Lactobacillus plantarum、L. rhamnosus、B. longum など) の投与が抗不安作用と抗うつ作用を持ち (222, 223)、認知機能の側面に影響を与える可能性があることが実証されています (224)。同様に、最近のメタアナリシスと系統的レビューは、プロバイオティクスが人間集団の不安、抑うつ、および主観的ストレスを改善する可能性について、予備的ではあるが有望な証拠を報告しました (225–228)。一方、最近の別のメタアナリシスとシステマティック レビューでは、利用可能なデータの質と量が限定されているものの、人間を対象とした試験から得られた現在のエビデンスは、プロバイオティクス、プレバイオティクス、発酵食品が認知機能に影響を与えることを支持していないと結論付けています。結論 (229);たとえば、同じ株のプロバイオティクスまたは同じタイプのプレバイオティクスを調査する研究が不足しており、プロバイオティクスおよびプレバイオティクス固有の効果がしばしば観察されます。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.人生の極限における食事微生物叢 - 腸 - 脳軸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 腸管の微生物のコロニー形成は、生涯を通じて連続したプロセスです。特に人生の最初の数年間、微生物群集は動的であり、成体微生物叢の一部になるいくつかの細菌は、人生の最初の数ヶ月の間にすでに腸にコロニーを形成しています (230–232)。思春期における微生物叢の継続的な発達がいくつかの研究で示されているが、極端な外部ストレッサー(例えば、食事の変化や抗生物質治療)がなくても比較的安定して回復力のある、個別化されたものではあるが、中核となる微生物プロファイルが成人期に発達する(233 –235)。加齢に伴い、微生物叢は再びより流動的になる可能性があり、これは虚弱と老化の加速に関連しています (23, 233, 236)。遺伝的要因、出産方法 [経膣分娩または帝王切開 (帝王切開)]、妊娠期間、運動、薬物使用、生活環境、および食事など、いくつかの要因が人生のさまざまな段階で微生物組成に影響を与えることが知られています (237– 240)。人生のさまざまな段階での微生物叢-腸-脳軸の食事操作の影響を研究することへの関心が高まっています(241)。ここでは、出生前から高齢者までの食事 - 微生物叢 - 腸 - 脳軸の既存の証拠の簡単な概要を提供します。グラフ表示を図 1 に示します。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
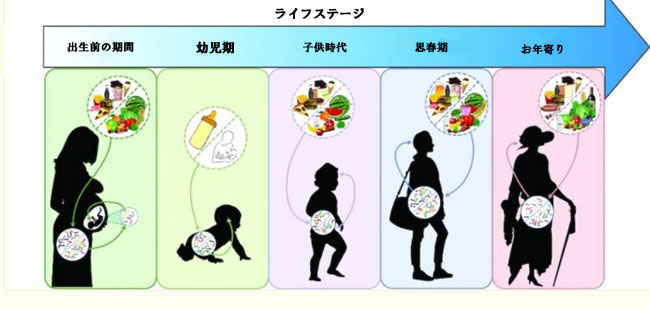 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 図 1 人生の極限における食事と微生物叢 - 腸 - 脳軸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 食事は、生涯にわたって微生物叢 - 腸 - 脳軸に影響を与える可能性があります。出生前の期間中、母親の食事は、潜在的に微生物叢を介したメカニズムを通じて、子孫の認知発達に影響を与えます。乳児期には、母乳または粉ミルクの授乳が微生物叢の構成に大きな影響を与えます。新たな研究は、これが脳と行動に影響を与える可能性があることを示唆しています。 「離乳反応」のタイミングは、微生物叢と脳の相互作用の発達を促進する上で重要である可能性があります。小児期および思春期における微生物叢 - 腸 - 脳軸の継続的な発達は、軸の適切な発達のために健康的な食事摂取が重要である可能性がある追加の敏感な時期を示す可能性があります。高齢者では、微生物叢が再び変化しますが、これは食事の摂取によって部分的に引き起こされる可能性があります。微生物叢のこれらの変化は、フレイル、「炎症老化」、および認知機能に関連している可能性があります。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.1.出生前の期間 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 出生前の期間中、食事を含む母体の要因は、子孫の微生物プロファイルに影響を与えるだけでなく、乳児の認知と行動に永続的な影響を与える可能性があります。適切な母親の栄養は、成長する胎児の発育の鍵であり、栄養不足は神経学的異常の確立された原因です(葉酸摂取量の低下や神経管欠損など。.同様に、ヒト以外の霊長類では、妊娠中の不健康な食事は、子孫の認知結果の悪化、ストレス反応の強化、および行動の混乱に関連しており(242)、人間の観察研究も、妊娠中の母親の食事が子供の感情的および認知的結果に関連しています( 243)。近年、動物モデルからの証拠は、腸内微生物叢が母親の食事と神経発達を結びつける根本的な要因である可能性を示唆しています (244–246)。高脂肪食または西洋食の影響に関する調査により、食餌によって引き起こされる微生物プロファイルの変化が、子孫の行動障害および認知障害を媒介することが明らかになりました (244–246)。 一方、ごく最近の実験データは、微生物代謝産物の作用を介して脳の発達の母体の微生物調節が起こる可能性があることを示しています(247)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 出生時に、新生児は消化管にコロニーを形成する最初の多数の微生物にさらされ、出生モードに基づく微生物叢組成の違いが報告されています。したがって、自然に生まれた乳児は母親の膣内細菌にさらされますが、帝王切開で生まれた赤ちゃんは、最初に母親の皮膚、病院環境、または医療従事者の細菌に感染します(237)。これらの違いは、生後数週間または数か月で消失する可能性がありますが(248)、生後1年以内に獲得された一部の微生物は、成体微生物叢の重要な居住者になることが示唆されています(249)。同様に、いくつかの他の研究は、微生物プロファイルに対する帝王切開の影響が 4 歳まで続く可能性があることを示唆しています (250)。帝王切開分娩に関連するこれらの微生物の変化(Bifidobacteriumの枯渇など)は、行動や認知に永続的な影響を与える可能性があります。しかし、これらの微生物および行動の変化の一部は、プレバイオティクス混合物 (ガラクトオリゴ糖、フラクトオリゴ糖) を補給することで元に戻すことができ、微生物叢を標的とした食事介入を使用して、C セクションに関連する負の影響の一部を軽減できることを示唆しています (251)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.2.幼児期 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 動物では、微生物叢が行動、神経化学、および脳の形態に長期にわたる影響を与える可能性がある、脳の発達中の重要なウィンドウまたは敏感な期間が特定されています(219)。 動物実験では、微生物へのこの初期の曝露が神経発達に不可欠であり、微生物叢の欠落または変化に関連するいくつかの行動への影響は、人生の後半で元に戻すことができないか、特定の期間中にのみ元に戻すことができることが示されています (218, 245, 252)。 . このように、幼少期の食事 - 微生物叢 - 腸 - 脳の軸にますます重点が置かれています。 実際、最初の 1000 d は、脳の健康を含むその後の健康をプログラムするために重要であることがわかっています (253)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 出生後、乳児の腸のコロニー形成は主に初期の摂食モードによって決定され、母乳で育てられた乳児と粉ミルクで育った乳児では微生物組成が区別されます(254, 255)。一般に、母乳で育てられた乳児は、Bifidobacterium 種が優勢な、多様性が低く、種が豊富な微生物叢を抱えていますが、人工乳で育てられた乳児の微生物叢は、機能的に成人の微生物叢に似ており、多くの場合、Klebsiella, Enterococcus, Peptostreptococcaceae, Akkermansia, Veillonella, および C. difficile などの微生物分類群が豊富です(256, 257)。 このように、母乳のさまざまな内因性因子が微生物叢の発達に影響を与えます。 母乳には、共生細菌、相利共生細菌、プロバイオティクス細菌などの独特の微生物群集があり、そのうちのいくつかは新生児の腸の最初のコロニー形成者として見られることがあります (258, 259)。 さらに、プレバイオティクスの人乳オリゴ糖 (HMO) は常在微生物によって発酵され、有益な細菌の増殖を促進します (260)。免疫グロブリン、抗体、抗菌ペプチド、ラクトフェリンなど、母乳に含まれる他の生物活性化合物は、微生物叢に依存しない利点を乳児にもたらす可能性がありますが、健康上の利点 [免疫疾患や胃腸疾患、肥満、2型糖尿病の発生率の低下など] を仲介します。 2型糖尿病、ならびに認知機能の改善 (261, 262)] は、健康な微生物叢の構築を通じて示唆されている (263–265)。幼い頃の乳児の腸内微生物叢の組成は、乳児期後半のその後の認知発達と行動の結果に関連していますが(266, 267)、これらの関係が母乳育児と早期の食事摂取によって媒介されるかどうかはまだ調査中です. マウスモデルにおける出生時からのオリゴ糖の補給は、微生物叢の組成と多様性を調節し、糖分解を増加させ、タンパク質分解発酵を減少させる一方で、社会的および不安様行動の改善ももたらしました (268)。微生物の豊富さ、組成、および酵素活性を変化させることにより、正常な神経発達をサポートします。ヒトのデータはこれまでのところ限られているが、生後 12 カ月の乳児における食物繊維に関連する Prevotella の相対的存在量は、最近、2 歳の子供の行動調節障害と関連していた(269)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| さらに、離乳または固形食の導入のタイミングは、腸と脳のコミュニケーションの重要な経路である微生物叢と宿主の相互作用、特に免疫系の適切な発達のための重要な要素として研究されています。離乳のタイミングを変えると、免疫応答の病理学的刷り込みと、後の免疫病理に対する感受性の増加につながる可能性があります(270)。子豚では、早期の離乳ストレスが腸のバリア機能を損ない(271)、炎症を増加させ(272)、酸化ストレスを増加させます(273)。人間の研究では、固形食品を早期に導入すると、肥満 (274, 275) または免疫疾患 (276) のリスクが高くなることが示されました。宿主の発生に対する微生物叢の既知の深刻な影響により、腸内微生物叢の成熟の加速は、宿主プロセスに対する早期離乳の有害な影響に寄与する経路として提案されてきた(275)。実際、動物研究では、いくつかの食事成分への初期の曝露が微生物叢の組成を変化させ、それが全身性炎症を助長し、血液脳関門 (BBB) 透過性の変化などの脳に影響を与える可能性があることを実証しました (277)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.3.幼年期および思春期 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 小児期および思春期に健康的な食習慣を奨励することは、成人期に健康的な食事を発達させるために不可欠であり(278, 279)、それによって全体的な幸福の基礎を築き、健康で成熟した微生物叢を確立します。 不十分な栄養による微生物叢の未発達は、包括的な結果をもたらす可能性があります。栄養不足の子供の糞便サンプルを 無菌マウスに移植すると、ヒト宿主で観察されたものと同様の代謝および免疫調節異常がマウスモデルで誘発され、栄養失調の症状の一部は特定の有益な微生物の欠如に起因する可能性があることが示唆された(280)。生理的影響に加えて、幼児期の低栄養の結果としての未熟な微生物叢は、神経学的異常と因果関係があると提案されています (281)。中等度の急性栄養失調の子供たちに、微生物叢に向けられた補完食品プロトタイプを補給すると、微生物叢と神経発達のマーカーが健康な子供の方向にシフトし、未熟な微生物叢が不健康な成長と発達に因果関係があることを示しています. しかし、微生物叢を標的とした食物療法によって救うことができます (282)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 以前は、固形食品の導入により微生物叢が「成熟」し、成人の微生物叢に似ていると考えられていましたが、いくつかの研究では、青年期にさらに発達することが示唆されています (283, 284)。思春期は神経解剖学的変化と成熟の重要な時期でもあり、認知機能、社会的認知、実行機能などの行動発達につながります (285, 286)。これらの成熟の延長は、成熟期の思春期の脳の微生物プライミングのための追加の敏感な期間をもたらし、思春期の精神的健康を改善する機会を提供する可能性があります (219, 287)。したがって、食事の摂取は、健全な微生物叢と脳のコミュニケーションの重要な推進力になる可能性があります。 果物や野菜の推奨摂取量を満たしていない不健康な食事やダイエット、脂肪や糖分の多い食べ物や飲み物の過剰摂取は、思春期の特徴である可能性があります(288–290). 人間のコホートから得られた調査結果は、思春期の脳と精神の健康にとって十分な栄養と食事の質の重要性を示しています (291, 292)。 この発達期間中の食事、腸内微生物叢、および脳の間の相互作用はほとんど研究されていませんが、ある研究では、ω-3 脂肪酸とビタミン A の補給が、微生物の障害と新しい物体認識の障害を逆転させることが示されました。 動物モデルにおける社会的不安定ストレスによって引き起こされる海馬および前頭前皮質の血清脳由来神経栄養因子レベルの変化も同様である(293)。 思春期のマウスモデルでは、21日間カフェテリア食にさらされると、種の均一性(シャノン多様性指数で測定)が減少し、Roseburia、Turicibacter、およびEnterorhabdusの量が減少し、行動的症状は観察されませんでしたが、遺伝子発現が変化しました前頭前皮質と扁桃体の神経免疫と神経伝達に関与している (294)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4.4.高齢者 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 人生のもう一方の極限である高齢者集団では、微生物叢が再び変化し、フレイル、炎症状態、認知機能などの宿主の健康面に影響を与えます (233, 234)。一般に、微生物の多様性が減少し、有益な細菌 (bifidobacteria, lactobacilli, Clostridium cluster XIVa, F. prausnitzii) の数が減少しますが、通性嫌気性菌および日和見菌、さらには炎症性病原菌 (Escherichia spp.、Enterobacteriaceae spp.、Bacteroides spp.、C . difficile など) は、若い人 (235, 295, 296) と比較すると増加します。 アルツハイマー病や血管性認知症などの認知機能低下の疾患は、健康な対照と比較した場合、異常な微生物組成と関連しています(297)。地理、生活状況 (長期介護施設またはコミュニティ)、薬の使用、および食事などのその他の環境要因は、高齢者の微生物と健康の相互作用に大きな役割を果たす可能性があります。したがって、健康的な食事は認知機能の健康を維持する上で基本的なものであるという確立された概念 (298) は、食事、微生物叢、および炎症の間の関連性によって部分的に媒介される可能性があります。これらの相互作用を理解するための調査が行われています。動物研究では、イヌリン補給による微生物叢組成の変化は、中年に関連する神経炎症障害を逆転させることができた(299). 高齢者の栄養に関する人間の欧州プロジェクト (NU-AGE) コホートでは、高齢者の地中海食への順守は、認知機能の改善を含む健康的な老化のマーカーと正の相関がある微生物分類群と関連していました (300)。したがって、高齢者の微生物叢と炎症を積極的に操作するために食事がどのように使用できるかを理解することは、認知能力を維持するための道となる可能性があります。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 既存の文献から、生涯にわたる食事と微生物叢と脳のコミュニケーションとの間の明確な相互作用が明らかになりました。 ますます多くの研究がこの相互作用を調査しているため、新しい発見は、神経発達と思春期の脳の成熟に対する微生物破壊の有害な影響を最小限に抑えるための早期介入戦略の開発に情報を提供し、高齢者集団が認知とメンタルヘルスを維持するための栄養療法を導くのに役立ちます。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.腸内細菌叢と行動を操作するための全食アプローチの使用 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 過去 10 年間の研究により、メンタルヘルスにとって適切な栄養が重要であることが明らかになりました。現在、健康的な食事パターンを一般的な精神疾患、特にうつ病のリスクの低下と関連付ける広範な観察データが、多くの異なる国や文化にわたって存在し(202)、新しい試験データは、食生活の改善が抑うつ症状を改善できることを示しています(301)。 地中海式食事は、メンタルヘルスを含む健康への影響に関して最も研究されている食事パターンですが (201)、世界の多くの地域の伝統的な食事パターン (ノルウェー料理や日本の食事など) も保護的な関連性を示しています (302, 303)。また、うつ病やアルツハイマー病を発症するリスクの低下、および認知機能低下の一般的な遅延と相関しています (201, 304)。 一方で、偏った食生活 (西洋式の食事など)、低品質、加工食品、または高脂肪/糖分の多い食品の摂取、および栄養失調 (過栄養および低栄養) は、メンタルヘルスの悪化(305 , 306)、認知機能障害 (298)、および不安 (307)、うつ病 (292, 308, 309)、または他の精神疾患を発症するリスクの増加に関連している可能性があります これらの関連性は、精神障害の主な発症年齢である思春期初期を含む年齢層全体で観察されます (292, 310)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| この食事と脳のつながりの根底にあるメカニズムは、ますます解明されつつあります。プロバイオティクスとプレバイオティクスが微生物叢と気分や認知に及ぼす影響はより広く研究されていますが (12)、自然食品や食事療法に関する調査はほとんどありません。最近のレビューでは、主にポリフェノールと微生物叢アクセス可能な炭水化物の含有量に起因する丸ごとの果物と野菜の、微生物叢と肥満や結腸の炎症などの関連疾患に対する利点が強調されています(16)。前臨床研究からのいくつかの証拠は、食事、微生物叢、および脳/行動の間の三角関係を示していますが、人間集団での同様の研究は不足しており、不安、うつ病、または認知に対する食事の影響を調査しているほとんどの臨床研究は、微生物叢の組成の変化を調査していません (199 、311、312)。臨床研究と前臨床研究の両方から、腸内微生物叢、神経化学、および行動に対する全食事療法の影響を調査した研究をまとめた非網羅的なリストを表 2 (表が大きすぎて和訳できませんでした)に示します。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
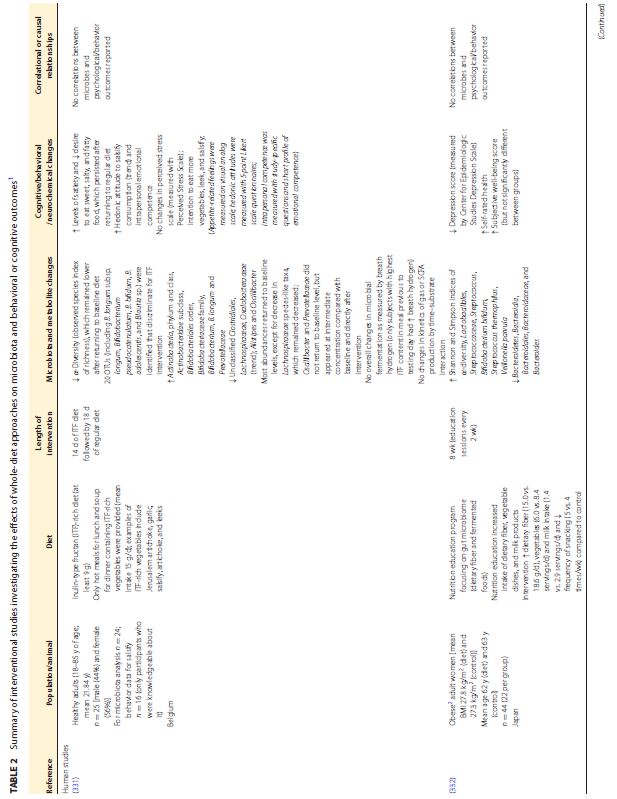 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
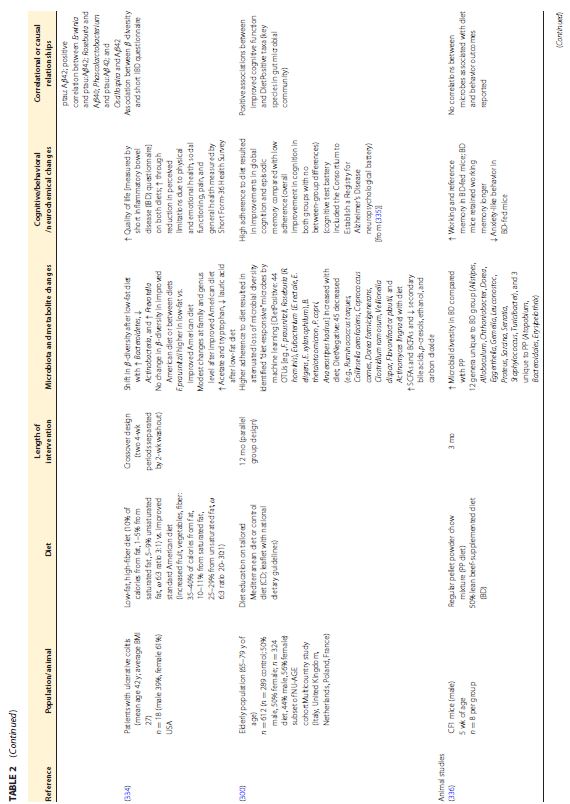 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
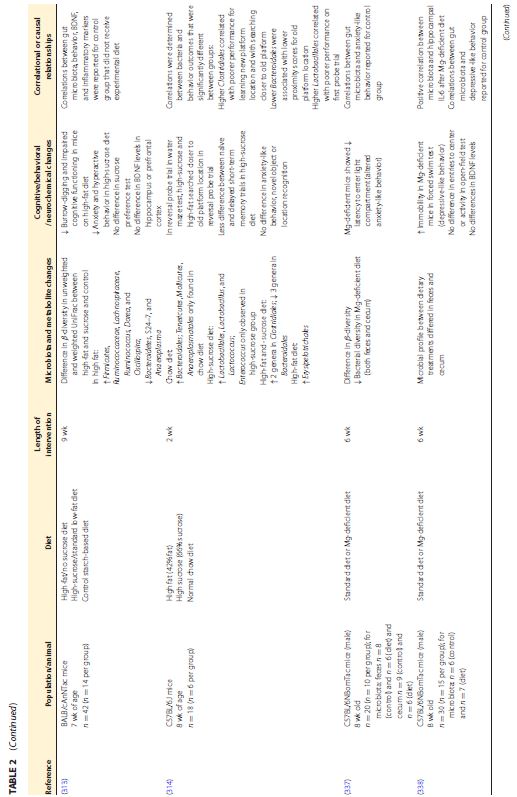 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
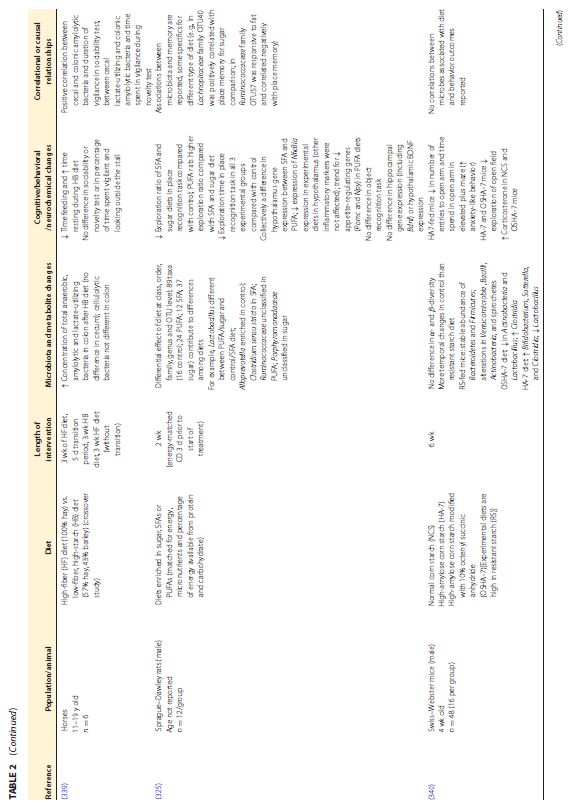 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
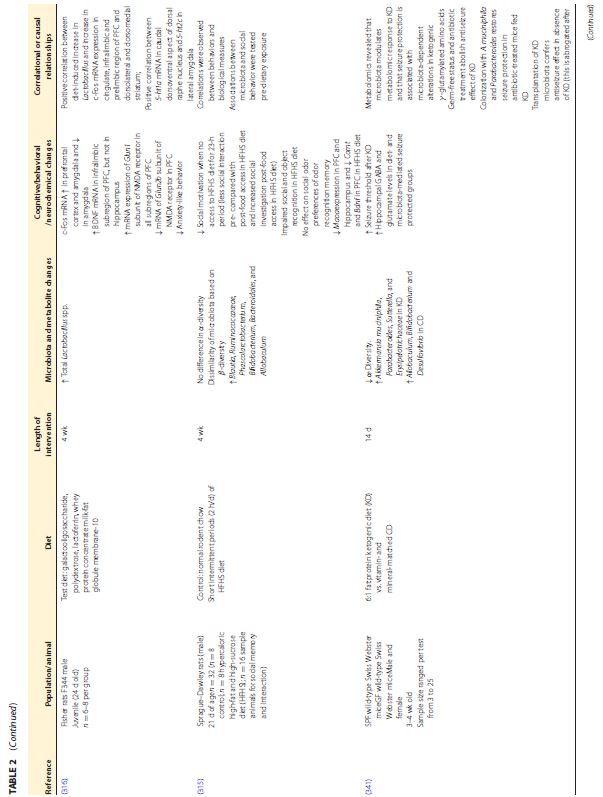 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
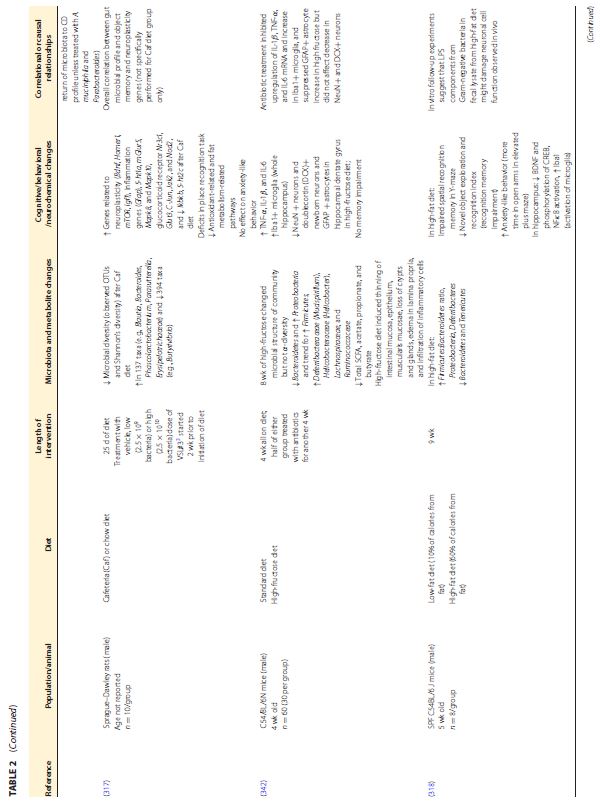 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
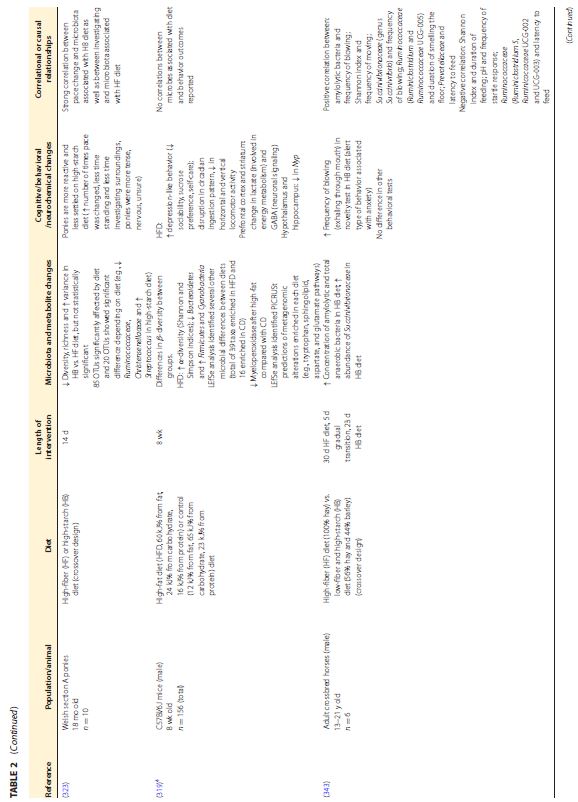 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.1.前臨床研究からの証拠 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 今日まで、多くの研究は、微生物叢と脳のプロセスに対する不健康な食事の影響を理解することに焦点を当ててきました。 高脂肪、高スクロース、または高カロリーの食事を与えると、記憶機能の低下、認知の柔軟性の低下、過活動行動、または社会的行動の変化によって証明されるように、認知と行動に悪影響を与える微生物叢の組成に好ましくない変化が生じます (たとえば、Firmicutesの量の増加とBacteroidetesの量の減少、およびClostridiales と Bacteroidalesの割合の増加)(313–315)。 さらに、神経化学の変化 (海馬における血清脳由来神経栄養因子の減少など)、ニューロン活動 (前頭前野および扁桃体における c-Fos 活性の増加など)、およびシグナル伝達 [γ-アミノ酪酸 (GABA) の変化など] の変化が増加します。脳の炎症 [例えば、ミクログリアの増加、炎症性遺伝子の発現、およびグリア線維性酸性タンパク質 (Gfap)] および神経可塑性に関連する遺伝子の発現 [例えば、血清脳由来神経栄養因子、ホーマータンパク質ホモログ 1 (Homer1)、ラパマイシンの哺乳類標的 (mTOR)、およびインスリン様成長因子 1 (Ifg1)] は、これらの食事の投与と関連していた (316,–319)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 一方、微生物叢と脳の相互作用を媒介する健康的な食事の潜在的な利点に関する調査は、ごく最近になり始めたばかりです。 例えば、最近の研究では、糖尿病マウスモデルを断続的に絶食させた後、行動、神経化学、脳構造の変化に関連する微生物組成と代謝産物の変化が観察された(320)。 別の最近の動物研究では、プレバイオティクスβ-グルカンの補給は、高脂肪、繊維欠乏食によって誘発されるミクログリアの活性化と神経炎症だけでなく、微生物叢の変化と認知障害を無効にしました(321)。 重要なことに、微生物叢の構成は認知変化の前に救済され、β-グルカンのプラスの効果は抗生物質治療で排除され(321)、食事による微生物の変化と認知機能との間の潜在的な因果関係が示唆された。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ほとんどの研究は微生物と行動の変化を別々に報告していますが、一部の研究では、食事による微生物組成の変化と行動の結果を関連付けようとしています。例えば、カフェテリアの食事へのさまざまな曝露時間によって誘発されるCoprobacterの違いは、空間認識記憶のパフォーマンスの予測因子として特定されました(322)。 同様に、高繊維食によって促進された微生物に関連するより安定した行動と同様に、高デンプン食によって誘発された行動反応性の増加と微生物プロファイルとの間の強い関連が馬で観察された(323)。 幼若ラットでは、Lactobacillus の食事に関連した増加と、ニューロン活性化およびセロトニン (5-HT) 受容体の mRNA 発現との間の正の相関関係が報告されており (316)、脳化学に対する食事の影響が特定の微生物によって媒介されていることが示唆されています。 最後に、最近の研究では、高脂肪食によって誘発された微生物叢の組成と、微生物叢に関連する血漿代謝物が、ラットの抑うつ様行動と関連し、その予測因子であることが示されました (324)。 興味深いことに、プロバイオティクス (VSL#3、bifidobacteria 3 株、lactobacilli 4 株、Streptococcus 1 株を含む) による事前治療は、海馬依存の場所認識タスクにおける食事誘発性の認知障害を防ぎ、カフェテリア食への曝露によって減少した分類群の特定の細菌を救うことができました(317)。 同様に、短期間の食事摂取 (2 週間) でさえ、微生物叢の構成を炎症関連経路と記憶障害に関連する形で変化させ (325)、微生物叢と脳機能への急速な影響を示しています。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5.2.ヒト試験からの証拠 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 食事操作後の抑うつおよび不安症状の改善を示す臨床的介入からの既存の証拠があるが (199, 311, 326)、これらの研究は、今日まで、腸内微生物叢のデータを収集していない。 ほとんどの人間の介入研究は、微生物叢を操作し、脳機能と精神的健康への影響を調査するために、プロバイオティクスとプレバイオティクスの補給に焦点を当ててきました (327, 328)。 たとえば、B. longum1417 は安静時の神経活動と社会的ストレスに対する神経反応を調節しましたが、β-ガラクトオリゴ糖の補給は、ストレスの生化学的尺度である唾液コルチゾール覚醒反応を減少させ、感情情報処理を改善しました (329)。オリゴフルクトース強化イヌリンは、健康なボランティアの記憶力と気分を改善しました (330)。 プロバイオティクスとプレバイオティクスが豊富な発酵食品は、腸と脳の軸に同様の有益な効果をもたらす可能性があります。 47 人の健康な若い医学生を対象としたパイロット研究では、 L. casei シロタ株を含む発酵乳飲料の摂取は、腹痛や風邪の症状などのストレスによる身体症状を軽減し、プラセボ群で観察された唾液中コルチゾールの増加を防ぎ、トリプトファン代謝におけるストレス誘発性異常を正常化し、セロトニン生合成を改善する可能性があります(344)。 健康な女性ボランティアを対象とした別の小規模研究では、5 つのプロバイオティクス菌株 (Bifidobacterium animalis subsp. lactis, Streptococcus thermophilus, Lactobacillus bulgaricus,2株および Lactococcus lactis subsp. lactis) を含む発酵乳製品を 4 週間摂取すると、広範な脳ネットワーク(すなわち、一次内受容領域および体性感覚領域、中脳中脳領域のクラスターであり、中脳水道周囲灰白質を中心とする)の活性が変化しました(345)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| これまでに発表された全食事アプローチを使用した介入研究の中で、ボランティアとして健康な成人を含めたのは1つだけで、イヌリン型フルクタンが豊富な食事の後に栄養行動と対人感情能力の改善が報告されました(331)。 肥満集団では、腸内微生物叢(繊維を含む食品や発酵食品の増加)に焦点を当てた栄養教育プログラムにより、うつ病スコアが減少し、自己評価された健康が増加し、B. bifidum や S. thermophilus などの有益なバクテリアが豊富に含まれているα多様性とが増加しました(332)。 炎症状態 (346)、トリプトファン - キヌレニン代謝 (347)、視床下部-下垂体-副腎軸活性 (348) など、うつ病に関連する他の生物学的結果は報告されていません。 したがって、この研究で観察されたうつ病スコアの低下が、全体的な主観的な肯定的な変化と身体の満足によるものなのか、それとも根底にある生物学的または微生物的メカニズムによるものなのかを解読することは困難です。 アルツハイマー病を発症するリスクのある高齢者の集団において、修正地中海食を摂取した後の微生物の変化は、脳脊髄液中のアルツハイマー病のバイオマーカーの改善と相関していた(333)。 食事介入が潜在的な保護特性を持つ細菌の変化につながる可能性があることを示唆しています。 NU-AGE コホートでは、地中海食によって誘発される微生物分類群の増加は、カスタマイズされた地中海食を 12 か月間続けた高齢者の認知力の改善と虚弱および炎症のリスクの減少と関連していた (300)。 人間を対象とした研究から説得力のある証拠が出てきていますが、利用できる研究の数が限られているため、精神的健康を改善したり、病気の症状を治療したりするための特定の食事の使用について、証拠に基づいた推奨を提供することは困難です (7)。 したがって、将来の高品質で大規模なコホート研究は、この有望な分野の理解を深めるために不可欠です。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.腸と脳のコミュニケーションの食事操作の根底にある提案されたメカニズム | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
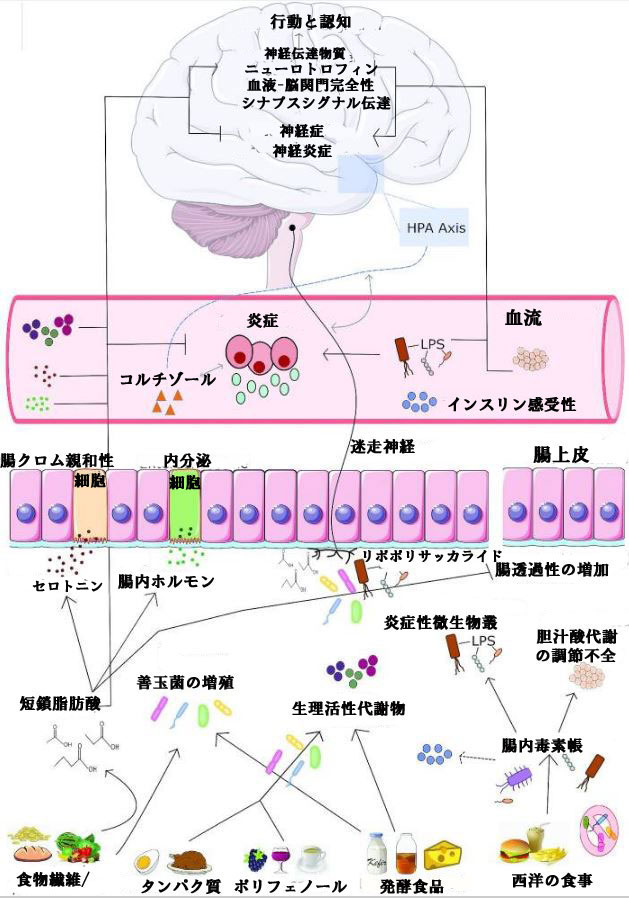
| 微生物叢と脳のコミュニケーションの複数のメカニズムが提案されています (2)。 これらのメカニズムのいくつかは、食事による調節を受けやすく、いくつかの調査では食事が脳に及ぼす影響の根底にあることが示唆されています。 これらのメカニズムの概要を図 2 に示します。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 図 2 食生活の調整を起こしやすい腸と脳のコミュニケーションのメカニズム | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 微生物由来の代謝産物、ホルモン、免疫、代謝、神経経路など、食事が腸と脳のコミュニケーションを調節する複数のメカニズムが存在します。健康的な食事(食物繊維、ポリフェノール、発酵食品など)は、有益な微生物の増殖を促進します。これらの微生物は、生理活性代謝産物、神経伝達物質 [セロトニン (5-HT) など]、腸ホルモンの産生を刺激し、直接的または間接的なシグナル伝達経路を通じて脳や行動に影響を与える可能性があります。コミュニケーションのもう1つの重要な手段は、食物分解または微生物からの微生物代謝産物による迷走神経の刺激です。不健康な食生活 (西洋の食事など) は、有害な細菌の増殖につながる可能性があります。この腸の「dysbiosis (腸内毒素症)」は、胆汁酸代謝、腸透過性、炎症、および代謝経路の変化を介して、機能不全の脳プロセスおよび神経炎症を引き起こす可能性があります。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.微生物代謝物 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.1.短鎖脂肪酸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 未消化の食物を代謝する微生物の能力により、生成された代謝産物は食事 - 微生物叢 - 脳の三角形の重要なメディエーターです。 代謝物が宿主の脳の機能と行動に影響を与える可能性があるさまざまなメカニズムが説明されています。 最近、断続的な絶食後に特異的に増加することが判明した微生物代謝産物の投与により、認知機能が改善され、動物の認知改善における微生物代謝産物の因果的役割が部分的に支持されました(320)。 おそらく最もよく研究されている代謝産物は、繊維の微生物発酵の生成物である 短鎖脂肪酸です。 重要なことに、短鎖脂肪酸受容体、主に遊離脂肪酸受容体 (349) およびモノカルボン酸トランスポーター (350、351) が中枢神経系 (CNS) および末梢神経系で発見されており、直接シグナル伝達の可能性が示されています。 さらに、短鎖脂肪酸は神経栄養因子 [神経成長因子、血清脳由来神経栄養因子、およびグリア細胞株由来神経栄養因子 (352)] または神経伝達物質の合成 [グルタミン酸、グルタミン、およびγ-アミノ酪酸 (353)] を刺激し、成長、生存、分化、中枢神経系のニューロンとシナプスの興奮性 (352) であり、学習、記憶、ストレス、気分に重要な役割を果たしています。 動物モデルでは、短鎖脂肪酸の投与は、心理社会的ストレスに関連する行動障害 (抑うつ様行動)、ストレス反応性、および腸透過性を低下させ (354)、短鎖脂肪酸が脳の恒常性と行動に直接影響を与える可能性があることを示しています。 さらに、短鎖脂肪酸、特に酪酸は、オクルディンの発現を増加させることによって血液脳関門の完全性を高めることも示されており (355, 356)、それによって潜在的な神経毒性因子から脳を保護します。 最後に、酪酸と、程度は低いが他の 短鎖脂肪酸も、ヒストン脱アセチル化酵素 (HDAC) の強力な阻害剤として作用することができる (357)。 ヒストン脱アセチル化酵素は一連の神経精神障害 [例えば、うつ病や統合失調症 (358)] に関与しており、ヒストン脱アセチル化酵素阻害剤は不安や恐怖に関連する障害の潜在的な認知エンハンサーである可能性があります (359)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 短鎖脂肪酸が腸と脳のコミュニケーションと脳機能に影響を与える他の経路には、免疫、内分泌、神経、迷走神経、およびその他の液性経路が含まれます (360)。 炎症に対する短鎖脂肪酸の効果は、腸バリアの改善によって媒介される可能性があり (361)、それによって、免疫分子と細菌リポポリサッカライド( LPS) が末梢に移行するのを潜在的に防ぎ、全身性炎症と最終的には神経炎症を軽減します (362)。 同様に、短鎖脂肪酸は、サイトカイン、樹状細胞、マクロファージ、T 細胞などの免疫細胞の分化と活性化を仲介することができます (363)。 短鎖脂肪酸はまた、胃腸管の迷走神経求心性神経を活性化し (364)、電気信号を変換して脳および脳機能の神経伝達物質レベルを調節します。 最後に、短鎖脂肪酸は、G タンパク質共役受容体 (GPCR) の活性化によって、グルカゴン様ペプチド 1 (GLP-1) やペプチド YY (PYY) などの他のさまざまな腸内ホルモンの分泌を刺激することで、脳回路に間接的に影響を与えることもできます。 これらのホルモンには神経活性の可能性がありますが、短鎖脂肪酸によって誘発される ペプチド YYおよび グルカゴン様ペプチド 1レベルの変化が脳機能にどのように関連するかはまだ決定されていません (360)。 興味深いことに、最近、短鎖脂肪酸は、気分と食物摂取の境界面にある重要なG タンパク質共役受容体であるグレリン受容体を介したシグナル伝達を弱めることができることが示された (365)。 これは、末梢と中枢神経系の両方にまたがる幅広い機能を備えた、腸脳軸で発現する重要なG タンパク質共役受容体を標的とする可能性について、短鎖脂肪酸および他の微生物叢代謝産物の新たな治療の可能性をさらに強調しています。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 脳機能に対する短鎖脂肪酸の潜在的な有害な影響が報告されており、特定の神経疾患の症状に寄与する可能性があることに注意してください。 たとえば、動物モデルでは、プロピオン酸塩は (脳室内または高濃度で脳に直接投与されますが) 自閉症のような行動を誘発する可能性があり (366, 367)、自閉症スペクトラム障害のヒトでは糞便中の短鎖脂肪酸レベルの上昇が報告されています (221, 368)。 . 同様に、無菌マウスでは、短鎖脂肪酸混合物を投与すると、パーキンソン病の特徴的な症状であるミクログリアの活性化と運動障害 (369) が生じ、パーキンソン病の症状における短鎖脂肪酸の役割を潜在的に示しています。 しかし、結果は決定的なものではなく (370–372)、特定の疾患状態における脳の健康における短鎖脂肪酸のメカニズムを理解するための追加の研究が必要です。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.2.タンパク質分解による代謝物 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 炭水化物消化からの代謝産物に加えて、他の栄養素の微生物消化からの代謝産物は、食事 - 腸 - 脳のつながりに関与している可能性があります。腸内微生物叢内のいくつかの分類群(Bacteroides, Clostridium, Fusobacterium, Streptococcusなど)は、食物タンパク質を分解する酵素能力(すなわち、プロテアーゼ)を持っており、タンパク質が豊富で繊維の少ない食事で微生物のエネルギー収穫が炭水化物からタンパク質源に移行する特に重要な役割を果たす可能性があります(373, 374)。 微生物のタンパク質分解活性は、宿主へのアミノ酸の利用可能性に影響を与えるだけでなく (375)、生理活性代謝産物 [例えば、神経伝達物質、分枝鎖脂肪酸、アミン、フェノール、インドール (376)] の生成ももたらします。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| いくつかの微生物は、γ-アミノ酪酸、ノルエピネフリン、ドーパミン、セロトニン などの食物タンパク質から神経伝達物質を生成する能力を持っています (377)。 これらの微生物由来の神経伝達物質が宿主の脳機能にどのように影響するかの正確なメカニズムはまだ明らかにされていませんが、迷走神経、免疫系、および神経伝達物質の合成のための前駆体の末梢利用の調節が候補経路として提案されています (2)。 したがって、これらの神経伝達物質の到達範囲は、局所的な効果を超えて、中枢神経系機能に重要な影響を与える可能性があります (378)。 たとえば、芳香族アミノ酸であるトリプトファン (セロトニン の前駆体)、チロシン、およびフェニルアラニン (どちらもドーパミンの前駆体) の微生物による代謝と、それらの異化産物は、マウスの短期記憶および作業記憶の障害と関連していることが最近報告されました。肥満のヒト被験者から微生物移植を受けた(379)。 分枝鎖脂肪酸は、高タンパクで低複合炭水化物 (すなわち、食物繊維) の消費シナリオにおいて、特に重要な微生物メッセンジャーである可能性があります。これは、これらの食事の消費が分枝鎖脂肪酸の微生物生産を増加させる可能性があることが研究で示されているからです (121, 380, 381)。 微生物由来の分枝鎖脂肪酸、イソ吉草酸、およびイソ酪酸は、上皮生理学および粘膜免疫系に関連付けられています (380、382、383)。 高レベルのイソバレレートは、人間の研究で抑うつ気分およびコルチゾールレベルと相関しています (384)。 過敏性腸症候群の患者では、イソ吉草酸と吉草酸の濃度は、腹痛と過敏症と正の関連がありました(385)。 最後に、食事中のコリンからのトリメチルアミン (TMA) の微生物生産にも大きな研究の焦点が置かれています。 吸収されると、トリメチルアミンはトリメチルアミン N-オキシドに変換され、これはさまざまな慢性疾患、特に循環器系に関連する疾患に関与している [例えば、脳卒中 (386)]。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.3.トリプトファン代謝物 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| トリプトファン代謝、特に セロトニン、キヌレニン、およびインドール誘導体の生成に至る主な経路は、腸内微生物叢によって厳密に調節されているようです (378, 387)。 これらの代謝産物は、微生物叢と脳のコミュニケーションにおいて重要な生物活性メッセンジャーとして機能し、食事摂取、特にタンパク質によって調節できます。 体内の セロトニン の大部分 (95%) は、食事のトリプトファンから腸内クロム親和性細胞によって生成されます。 腸で生成された セロトニンが血液脳関門を通過できないため、脳への直接的な影響はありそうにありませんが、迷走神経の求心性神経の刺激と免疫応答の調節は潜在的なシグナル伝達経路である可能性があります (388, 389)。 さらに、セロトニンの産生は末梢のトリプトファンの利用可能性に影響を与える可能性があり、したがって脳の セロトニン レベルに間接的に影響を与える可能性があり、5-HT の末梢の変化が中枢の セロトニンシグナル伝達系に影響を与える可能性があることを示唆しています (390, 391)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| トリプトファン代謝の他の代謝産物であるキヌレニンも、一連の神経生物学的機能に関与している(392)。キヌレニンは血液脳関門を通過し、キヌレン酸やキノリン酸などの神経活性グルタミン酸作動性産物にさらに代謝される可能性があります。 N-メチル-d-アスパラギン酸 (NMDA) 受容体に対するアンタゴニストとして、キヌレン酸は通常神経保護的ですが、キノリン酸はN-メチル-d-アスパラギン酸受容体に対するアゴニストとして作用し、神経毒性効果を発揮します (393)。しかし、高濃度では、両方の代謝産物が神経伝達を妨害し、認知障害を誘発する可能性があります (394)。キヌレニン経路の調節不全は、うつ病 (395)、統合失調症 (396、397)、および自閉症スペクトラム障害(398) を含むいくつかの脳障害に関連しています。実際、いくつかの予備的な臨床介入研究では、プロバイオティクスおよびプレバイオティクスの介入がトリプトファン - キヌレニン代謝産物を調節できることが報告されています。例えば、臨床的うつ病患者を対象とした 2 つの無作為対照試験では、プラセボと比較して、プロバイオティック介入を受けた後のキヌレニン:トリプトファン比とキヌレニン濃度の変化が報告されました (399, 400)。同様の結果が、2型糖尿病の女性のサンプルにおける耐性デキストリンのランダム化比較試験で報告されました(401)。ポリフェノールなどの他のプレバイオティック食事成分も、キヌレニン代謝を調節する可能性があります (402, 403)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 多くの重要な有益な機能 (例えば、抗酸化効果) がインドールに関連付けられていますが、これらの代謝産物の過剰生産は、動物に不安や抑うつのような行動をもたらしました (404)。 さらに、インドールは特定の受容体 (例えば、アリール炭化水素受容体) に結合することによって炎症関連遺伝子の発現を増加させ、セロトニン受容体に結合して行動や腸の運動性に影響を与えることができます (405)。 ヒトのトリプトファン代謝産物に対する食事の直接的な影響は、最近のパイロット研究で実証されました。この研究では、地中海食とファーストフードの食事の摂取がトリプトファン代謝産物に異なった影響を与えました(205)。 神経細胞に有益な効果を与えることが示されているインドール-3-乳酸とインドール-3-プロピオン酸の増加は、地中海食後に観察されましたが、これらの代謝産物はファーストフード食後に減少しました。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.4胆汁酸 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 脂質消化を助けるために肝臓と細菌の酵素によって生成される胆汁酸は、脳機能に対する脂肪と微生物叢の相互作用効果に関与している可能性があります(406)。 微生物由来の二次胆汁酸は、宿主の生理機能だけでなく、腸内微生物叢の構成と機能に影響を与える能力があるため、王国間シグナル伝達分子であるという仮説が立てられています (407, 408)。 最近、異常な胆汁酸代謝が神経疾患で報告され、アルツハイマー病の発症/進行および認知機能低下の要因として、腸内微生物叢による胆汁酸の変換の役割が示唆されました (409, 410)。 胆汁酸がシグナルを伝達できるさまざまな中枢神経系経路が提案されています。 いくつかの報告は、胆汁酸が血液脳関門を通過できることを示唆していますが、他の胆汁酸コンジュゲートは、ファルネソイド X 受容体および TGR5 に存在するタケダ G タンパク質共役受容体 5 (TGR5) に結合することによって、胃腸管でこれらの受容体に結合し、シグナル伝達カスケードを開始することにより、中枢神経系または間接的に神経活性リガンドとして脳に直接信号を送ることが示されています。(411)。 ラットでは、胆汁酸は血液脳関門の透過性を増加させることも実証されており (412)、他の神経活性代謝産物または神経毒の脳への流れを増加させ、脳の機能と行動に影響を与える可能性があります。 胆汁酸が微生物叢が特定の脳プロセスに影響を与える重要な経路である可能性があることを示唆する予備的な証拠しかありませんが、 最近の実験データは、西洋食によって誘発された腸内細菌叢の変化が神経炎症を引き起こし、胆汁酸合成の調節不全と TGR5 シグナル伝達の混乱を介してシナプス可塑性を低下させたことを示唆している (406)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.1.5.その他の代謝物 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ポリフェノールやフェノール化合物、フィチン酸塩の代謝から生成される生理活性分子など、腸と脳のコミュニケーションを仲介する他の微生物代謝産物が発見されています (413)。たとえば、植物の茎やさまざまなハーブに含まれるフェノール化合物であるフェルラ酸は、Lactobacillus属のエステラーゼ活性によって分解されます。 4-ビニルグアヤコールとヒドロフェルラ酸 (414) に変換され、さらにカフェイン酸とバニリン酸に変換されます。これらの化合物は、アルツハイマー病などの神経変性疾患に対して強力な治療効果があり (415)、マウスの酸化ストレスと認知障害を軽減し (416)、血液脳関門破壊と不安様行動を軽減することができます (417)。さらに、いくつかの研究では、生物学的に利用可能な複数の微生物叢由来のフェノール酸が、炎症およびシナプス可塑性に関連するメカニズムを調節できることが示されました (418, 419)。同様に、地中海食によく見られる食餌性ポリフェノール (すなわち、イソフラボンとリグナン) の有益な効果のいくつかは、微生物ポリフェノール分解の代謝産物に起因する可能性があります。これは、より高い血液脳関門透過性を示し、神経炎症ストレスに対して、これは、ポリフェノールの微生物代謝が脳の完全性と精神的健康を保護するメカニズムであることを示唆しています (420)。最後に、ナッツ、豆、穀物に豊富に含まれるフィチン酸の微生物分解から生成される代謝産物であるイノシトールリン酸は、ヒストン脱アセチル化酵素 3 の強力な調節因子であることが示されました (421)。神経精神障害における ヒストン脱アセチル化酵素の潜在的な役割を考えると、これは、食事と脳の接続の界面における別の潜在的な微生物代謝産物である可能性があります。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.2.免疫シグナル伝達 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| バランスのとれたマイクロバイオームは、健康な免疫系の発達と維持に必要であり、この平衡の崩壊は、長期にわたる健康への影響をもたらす可能性があります (422)。 炎症は、うつ病を含むさまざまな精神疾患の根底にある原因として特定されているため (423, 424)、免疫系は腸内微生物叢とメンタルヘルスの間の重要なリンクとして浮上しています。 栄養は、この相互関係の極めて重要な調節因子となる可能性があります (425, 426)。 動物実験では、高脂肪食の摂取が、潜在的に認知障害を引き起こす可能性のある「炎症誘発性」微生物プロファイルを促進することにより、結腸、末梢、および神経の炎症を増加させることが示されています (427, 428)。 同様に、加工食品を多く含む食事によって引き起こされる腸内微生物のプロファイル (細菌の多様性の減少、組成の変化) は、低度の全身性炎症および酸化的変化のトリガー要因として認識され、神経変性疾患および炎症関連疾患の発症を助長します (429, 430)。 食事由来の代謝産物、免疫調節微生物の適応度の調節、微生物の組成と活動の変化など、免疫調節性食事因子の宿主または微生物代謝の変化と同様に食事 - 微生物叢 - 免疫相互作用を説明するさまざまなタイプのメカニズムが提案されています (431)。 一例として、次のシナリオが提案される可能性があります: 不健康な食習慣は、微生物叢と腸の透過性を介した炎症誘発性シグナル伝達の増加をもたらし、いわゆるリーキーガットを促進します (362)。 その結果、免疫細胞と細菌成分 (リポポリサッカライドなど) は、炎症を起こした腸管から逃れ、循環に移行する可能性があります。 結果として生じる全身性低悪性度炎症は、最終的に、脳内皮細胞上の Toll 様受容体への細菌リポポリサッカライドの結合、炎症誘発性転写因子 NFκB (活性化 B 細胞の核因子κ-軽鎖エンハンサー) シグナリングまたは MAP (マイトジェン活性化タンパク質) キナーゼ経路 (432)。 いくつかの報告では、全身性炎症が血液脳関門の完全性を損なう可能性があり、脳からの外来分子の脳への通過を可能にし、サイトカインの放出を引き起こし、ミクログリアと星状細胞の炎症誘発性を活性化し、神経炎症とニューロンおよび神経と脳のプロセスの破壊を開始する可能性があることを示唆しています。 (426、433、434)。 また、微生物叢が髄膜 (脳と脊髄を覆う膜) の免疫細胞 (特に IgA) の存在に影響を与え、病原体の脳への浸潤を防ぐことができることも動物モデルで最近実証されました (435)。 したがって、食事によって変化した微生物叢は、防御免疫細胞の量を減らし、神経炎症を引き起こす細菌の侵入を可能にする可能性があることが示唆される。 貧弱な食習慣 (つまり、高カロリー/脂肪の多い食事) に関連する微生物叢が誘発する炎症は、迷走神経のリモデリング (436, 437) または神経伝達物質の合成と分泌の変化 (438) を通じて、行動と認知に有害な影響をもたらす可能性があります。 また、血液脳関門タンパク質を分子的に模倣する食物の断片がリーキーガットを通って移行し、免疫応答を誘発して、血液脳関門の完全性を攻撃して損なう抗体を産生する可能性があることも示唆されています (439)。 最後に、炎症もキヌレニン経路の調節不全を引き起こす可能性があります。末梢炎症の増加に伴い、サイトカインはキヌレニン経路を刺激し、脳へのキヌレニンの供給を増加させ、キヌレン酸やキノリン酸などの下流代謝産物の産生を促進し、コリン作動性、グルタミン酸作動性、およびドーパミン作動性神経伝達を妨害します (392, 394)。 うつ病では、キヌレニン経路が炎症と抑うつ症状の間のつながりとして提案されています (395, 440)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 炎症が宿主の健康に与える影響に関する知識が蓄積されているため、抗炎症食が臨床診療における治療アプローチとして提案されています (441)。抗炎症食の中心となるのは、ポリフェノールを多く含む野菜や果物、植物由来のタンパク質源、魚、全粒穀物、オリーブ オイルを摂取することです。また、ハーブ、スパイス、サプリメントも取り入れます。この食事は、抑うつ症状を軽減するための潜在的に効果的な治療法として示唆されており、観察研究では、抗炎症性の食事パターンをより順守することでうつ病のリスクが低下することが示されています (442)。 増加する証拠はまた、微生物叢の食事調節が(高脂肪食の場合)または炎症状態を改善する(高繊維摂取に関連して)可能性があることを示唆しています。パーキンソン病やアルツハイマー病などの疾患 (443)。 実際、一部の研究では、メンタルヘルスに対する健康的な食事の利点の根底にあるメカニズムとして、炎症の減少が強調され始めています。 最近、ケフィアが微生物叢を明確な免疫学的および行動的効果に向けることが実証され、微生物叢-腸-免疫-脳軸を介したシグナル伝達カスケードが示唆された(58)。 発酵食品のこれらの抗炎症特性は以前に提案されており、一部は有益な微生物または生物活性化合物の増加に起因する可能性があります (444)。 同様に、高繊維食品を摂取することによる認知的および精神的健康上の利点は、微生物叢と免疫の相互作用に対する栄養の調節的役割に起因する可能性があります。. 例えば、短鎖脂肪酸は免疫調節性があり、G タンパク質共役受容体を刺激し、自然免疫応答を促進し、制御性 T 細胞を誘導します (445)。 短鎖脂肪酸の補給は、腸のバリア障害を軽減することにより、高果糖食に関連する神経炎症および神経細胞の損失から保護されます (342)。 別の研究では、高脂肪、繊維欠乏食を与えられた動物において、微生物叢アクセス可能な炭水化物が微生物の変化を防ぎ、腸の密着結合を強化し、結腸、全身、および神経炎症を軽減し、シナプスシグナル伝達分子と認知障害を改善したことが示されました (446)。 これらの肯定的な効果は、抗生物質治療後には観察されず、微生物叢が食事、炎症、および認知機能障害の間の相互作用における重要な調節因子であることを示唆しています。. 同様に、難消化性デンプンの補給は微生物の変化を元に戻し、全身性炎症を改善し、高脂肪食ラットの迷走神経求心性繊維のリモデリングを防ぎました(447)。 また、プレバイオティクス (10% オリゴフルクトース強化イヌリン) は、中年マウスのストレス誘発性微生物変化、免疫プライミングおよびミクログリア活性化を逆転させました (299)。 食事成分の他の微生物代謝物、すなわちトリプトファン誘導体は、ミクログリアと星状細胞の活動を調節することによって神経炎症を軽減する可能性があります (448, 449)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.3.迷走神経と神経機能 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 画期的な動物研究では、迷走神経切除後に微生物の特定の行動への影響が消滅することが実証され、迷走神経が微生物叢由来の信号を脳に伝達するもう 1 つの重要な役割を果たし、迷走神経が最も直接的な伝達経路となっていることが明らかになりました (450–452)。 簡単に言えば、腸上皮の下に位置する迷走神経求心性神経は、腸内微生物または微生物代謝産物によって刺激される可能性があります。 迷走神経シグナル伝達を使用することが示されている微生物には、病原体 C. jejuni (453) または共生生物 L. rhamnosus および B. longum (450, 451) が含まれます。 迷走神経刺激活性を持つ微生物代謝産物の中には、腸の迷走神経末端を活性化できる S短鎖脂肪酸、具体的には酪酸 (454)、および迷走神経求心性ニューロンに存在する受容体に結合できる γ-アミノ酪酸などの神経伝達物質 (455) があります。 同様に、細菌リポポリサッカライドによる迷走神経刺激は神経炎症を引き起こし、脳機能を変化させ、動物モデルで抑うつ様または不安行動を誘発することが示されました (456)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 食物摂取の調節における迷走神経の役割は、かなり長い間評価されてきました (457)。 最近では、前臨床研究で、食事によって誘発される腸内微生物叢の変化が、迷走神経の腸と脳のコミュニケーションを混乱させる可能性があるといういくつかの証拠が提供されました (436, 437, 458)。 例えば、高脂肪/高糖食は微生物の変化を誘発し、腸の炎症と腸の透過性の増加を引き起こし、ミクログリアの活性化と迷走神経のリモデリングを増加させました (436, 437)。 興味深いことに、迷走神経のリモデリングは抗生物質治療後に抑制され(436)、これは、微生物叢が迷走神経シグナル伝達に対する高脂肪食の有害な影響を媒介することを示唆しています。 同様に、高脂肪食によって誘発された視床下部の炎症は、迷走神経切除術によって軽減されました (459)。これは、迷走神経が食事誘発性と微生物叢関連の神経炎症の間の重要な結合であることを示しています。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 今日まで、食事 - 微生物叢 - 脳の三角形の経路としての他の神経機能に関する調査は不足しているため、さらなる調査が必要です。 しかし、プロバイオティクス菌株 (B. longum および L. rhamnosus) がニューロンの興奮性と発火能を変更したことを示す研究 (460, 461) は、プロバイオティクス菌株を含む食品、つまり発酵食品の健康上の利点は、神経細胞の変化によって部分的に媒介される可能性があることを示唆しています。 . 最近、神経精神障害、微生物叢、および神経活動の複数の側面の調節の間の根底にある関連性が、いくつかの疾患に対する有望な将来の治療法として提案されている(462)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.4.ホルモン経路 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ホルモンは、栄養素の消化と吸収、および食物摂取の調節において長い間確立されてきましたが、腸内微生物叢がこれらの腸内ペプチドのレベルを調節できるという考えは最近になって登場しました。 腸内微生物叢がコレシストキニン、ペプチドYY、GLP-1、グレリンなどの宿主ホルモンに影響を与えるさまざまなメカニズムが提案されています。これには、いくつかの微生物による直接的な産生だけでなく、代謝産物または微生物成分を介した腸内分泌細胞の調節による間接的なメカニズムも含まれます(463、464)。 これらのホルモンの受容体は、脳のさまざまな領域 (465, 466) または迷走神経求心性終末 (467) で確認されており、一部は 血液脳関門 を通過して受容体に直接結合することが示されています (468)。 したがって、それらの機能は、腸の運動性の局所的な調節を超えて、食欲、気分、不安、および抑うつの中央制御にまで拡張することができます (464)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 食事-微生物叢-脳の三角形における腸ホルモンの役割を直接調査した研究はありませんが、微生物代謝産物を介した間接的な経路が提案される可能性があります。. 発酵性の高いプレバイオティクスは、微生物叢によって誘発される GLP-1 と ペプチドYY の変化に影響を与え、動物 (469) と人間 (470) の両方の研究で、満腹感の増加、空腹感の減少、食欲の変化をもたらしました。 食事 - 微生物叢 - 脳の相互作用における腸内分泌機能の関与に関するいくつかの初期の証拠を提供します。 腸内微生物叢の食事操作に関連するホルモンの変化が行動の結果につながるかどうかを調査するより多くの研究が必要です。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.5.代謝経路 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 伝統的に、インスリンは血糖恒常性を維持する機能で最もよく知られています。現在、ますます多くの文献が、インスリンとインスリン受容体の利用可能性が、必要なエネルギー源を提供するためだけでなく、適切なニューロン活動とシグナル伝達回路を確保するためにも、正常な脳機能に関連していることを示唆しています[例えば、ドーパミン作動性およびセロトニン作動性システム( 471)]。したがって、当然のことながら、インスリン抵抗性は神経学的健康と認知障害に関与しており(472)、インスリンシグナル伝達の低下と神経変性疾患の病因との関連が提案されています(471)。微生物叢の組成と末梢および中枢のインスリン感受性との間の確立された直接的な関連性(473)、および高脂肪食が微生物叢の組成を変化させ、代謝変化(インスリン抵抗性およびグルコース恒常性を含む)ならびにうつ病に寄与することを実証する動物研究により、不安様行動 (474)、高脂肪食に関連する微生物の変化、代謝機能不全、および心理的問題の間の根底にあるメカニズムが提案される可能性があります (475)。 2 つの動物の食事介入は、微生物叢の標的化に成功し、代謝を改善し、その結果、認知機能を改善しました。 微生物叢アクセス可能な炭水化物 (MAC)と断続的な断食(320)の補給は、インスリン抵抗性マーカーとシグナル伝達の改善に起因する脳パラメーターを改善しました(446)。 両方の研究で、抗生物質の投与は、代謝および認知パラメーターで観察された食事の影響を無効にしました。これは、微生物叢が食事に関連する改善に必要であることを示唆しています。 微生物代謝産物は、インスリンとニューロンの健康の相互作用においても興味深い可能性があります。一部 (イノシトールなど) はインスリン感作効果があり、それによって CNS の機能に寄与する可能性があることが示されているためです (476)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6.6.視床下部-下垂体-副腎軸(HPA) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 視床下部-下垂体-副腎軸は、哺乳類におけるストレス反応の主要な神経内分泌調節因子です。視床下部-下垂体-副腎軸の調節不全は、うつ病などのさまざまなストレス関連の神経精神障害に長い間関与してきました (477, 478)。 視床下部-下垂体-副腎軸の調節における腸内微生物叢の極めて重要な役割の証拠は、腸内微生物叢の非存在下での視床下部-下垂体-副腎軸の過敏性を示す研究から得られます (218)。 プロバイオティクスまたはプレバイオティクスの補給後にコルチコステロン (479) またはコルチゾール (329) のレベルの低下を観察する前臨床および臨床研究からも同様です。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 栄養介入は、視床下部-下垂体-副腎軸の活動を正常化することが示されています。ビタミンC(480)、魚油(481)、またはポリフェノールが豊富なダークチョコレート(482)の補給は、ヒトコホート研究でコルチゾールレベルと主観的ストレス測定値の低下をもたらしました。 自然食品の食事、特に炭水化物の増加は、太りすぎまたは肥満の女性の唾液中コルチゾールレベルを改善しました (483)。 特定の種類の炭水化物に関する詳細な情報は提供されていませんが、短鎖脂肪酸が 視床下部-下垂体-副腎軸の活性を調節し、急性ストレッサーに対するコルチゾール応答を減衰させることを示す他の研究 (484) は、食物繊維の微生物代謝が視床下部-下垂体-副腎軸の調節に関与していることを示唆している可能性があります。 . 腸内微生物叢の組成が特徴付けられていないか (483)、短鎖脂肪酸」が結腸に直接投与されたため、微生物の作用を迂回し (484)、視床下部-下垂体-副腎軸とメンタルヘルスに対する栄養のこれらのプラスの効果が微生物叢によって媒介されるかどうかはまだ決定されていません。 しかし、微生物叢に向けられた食事介入を通じて視床下部-下垂体-副腎軸の活動を標的にすることが提案されており (485)、いくつかの研究では、腸内微生物叢がストレス応答の栄養調節に関与している可能性があることが示されています。 例えば、前臨床研究では、乳脂肪球膜とプレバイオティクスを補給することで、幼少期の母子分離によって引き起こされる視床下部-下垂体-副腎軸の調節不全と認知機能障害が軽減され、微生物叢の組成にも影響を与えました (486)。 慢性的な予測不可能な社会的ストレスの別の動物モデルでは、プレバイオティクスの投与により、ストレスによる微生物叢の変化とコルチコステロンレベルの上昇が正常化されました(487)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 微生物叢と脳のクロストークの根底にあるメカニズムを媒介する食事の役割を理解し始めていますが、関係を完全に解明するには追加の研究が必要です。 確かに、人間が毎日摂取する食物成分の組み合わせと、食事 - 微生物叢 - 脳シグナル伝達の複雑なメカニズムにより、多数の絡み合った経路が食事 - 微生物叢 - 脳の相互作用の根底にある可能性が最も高い。 例えば、短鎖脂肪酸は消化管で神経伝達物質と腸ホルモンの産生と放出を刺激し (353, 360)、視床下部-下垂体-副腎軸は迷走神経 (488) および免疫系 (489) と密接に相互作用します。 したがって、全食アプローチは、食事によって誘発される微生物プロファイルが脳機能と精神的健康に影響を与えるさまざまなメカニズムを開始する可能性があります。 包括的な新しいレビューでは、腸内微生物叢を介したものを含め、食事がうつ病に影響を与える可能性があるメカニズムに関する最新の知識がカバーされています (490)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7.食事介入に対する応答者と非応答者 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 人間の健康を改善するために微生物叢を対象とする研究の増加に伴い、これらの介入に対する代謝反応の個人間変動がますます説明されており (例えば、491–493)、参加者の 30% 未満が望ましい結果に達していると報告している研究もある (494)。 個人がどの食事から利益を得ることができるかを特定することは、特定の疾患に対する食事療法の開発だけでなく、個別化された栄養アプローチの設計においても重要な考慮事項です。 年齢、性別、遺伝学、運動、ベースライン微生物叢組成、および習慣的な食事パターンを含むがこれらに限定されないさまざまな要因が、個人の微生物および全身の反応を決定する可能性があります (495–498)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ベースラインの微生物叢組成に関しては、細菌グループの比率と特定の微生物の存在が、食事介入の成功の重要な決定要因を構成すると説明されています。 食品成分を代謝する微生物分類群の特殊な能力により、応答性の高い微生物群集の構成は、関心のある食事介入に依存します。 いわゆるエンテロタイプの使用は、食事介入に対する個人の反応を決定する方法として、また個別化された栄養のアプローチとして提案されています (499)。 エンテロタイプは、Prevotella、Bacteroides、または Ruminococcus が優勢な微生物叢のクラスターとして 2011 年に初めて報告されました (88)。 それ以来、これらのエンテロタイプに基づいて参加者を層別化すると、特に代謝の改善と体重減少に関して、食事介入 (特に繊維) に対する反応が予測されることが研究で示されています (70, 500)。 一方、カロリー制限食の成功は、A. muciniphila (68)、一部のclostridial 種 [Eubacterium ruminantium、Clostridium felsineum、や C. sphenoides (501)] または豊富なLactobacillus, Leuconostoc,やPediococcus (502)のベースライン量に依存していました。 人間の胃腸系の微生物は共同体の一部であり、多くの場合、孤立して機能するのではなく、生存と成長のために他の微生物との複雑な相互作用に依存しているため、微生物群と微生物間相互作用の存在について食事介入の成功は偶発的であることが示されました。 線形判別分析を使用して、Zhang et al (503) は、発酵乳製品の介入に感受性のあるラットと耐性のあるラットを区別する 43 の運用分類単位 (OTU) を特定しました。 人間の介入研究では、ネットワークの複雑さの分析により、非応答者では微生物種間の負の相互作用が介入後に増加したことが明らかになりました(70)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ベースラインの細菌の豊富さと多様性は、食事介入の成功のもう1つの重要な予測因子かもしれません(66, 504)。したがって、繊維に対する個人の反応は、最初の標的細菌レベルに依存する可能性があり、ベースラインで繊維摂取量が最も少なく、微生物の豊富さが限られている個人は、繊維摂取量の増加から最大の利益を得て、より大きな微生物叢の変化を示す可能性があります(505)。 また、微生物の安定性が食事介入に対する反応性を決定する可能性があることも示唆されています。 したがって、比較的安定した微生物叢は安定した食事から恩恵を受ける可能性がありますが、不安定な微生物叢は、食事介入に対する柔軟な対応と最適な食事の絶え間ない再評価を意味する可能性があります(506)。 最後に、食物成分を代謝する酵素活性を持つ微生物を宿す微生物遺伝子の豊富さ、および微生物代謝産物の濃度が、食事介入反応の決定的な要因として説明されました。 実際、特定の栄養素(ポリフェノールなど)の有益な全身効果の程度は、栄養素を生物学的に利用可能な化合物に変換する個人の微生物叢の表現型の能力に依存します(507)。. 例えば、ザクロ由来のポリフェノールであるエラグ酸、プニカラギン、エラジタンニンを摂取した後の生物活性代謝物であるウロリチンAの速度と濃度には、かなりの個人差がある(508)。 さらなる例はイソフラボン代謝産物であるエクオールで、かなりのエストロゲン特性を持っていますが、人口の半分以上はそれを産生するための微生物叢組成を欠いています (509)。 食事介入を成功させるためにどの微生物酵素レパートリーが必要かは、研究食に含まれる栄養素に依存します (70, 494, 510)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 微生物叢の構成に対する食事の顕著な影響により、介入前の習慣的な食習慣も微生物叢の応答性の重要な決定要因になる可能性があります。 研究によると、長期間にわたる不適切な食生活は、一部の微生物の潜在的な絶滅につながる可能性があることが示されています (13, 511)。 したがって、健康的な食事 (つまり、高繊維食) に反応する微生物の酵素能力が失われている可能性があります。 したがって、個体が食事介入に応答するためには、不足している微生物の投与が必要になる場合があります。 実際、食事とプロバイオティクスを組み合わせたアプローチは、ハミルトン不安評価尺度によって測定されるように、小規模な研究での各介入単独よりも、不安症状の大幅な減少につながりました(512)。 プレバイオティクス補給研究では、ベースラインで繊維摂取量が多い参加者は、習慣的な繊維摂取量が少ない参加者よりも、イヌリン型フルクタン プレバイオティクスに反応して、より大きな微生物反応を示しました (513)。 他のいくつかの研究では、個人のベースラインの食事が、食事介入に対する全身および微生物の反応を予測したことが報告されています (70, 514)。 これらの研究は、参加者の徹底的な食事評価が微生物叢を標的とした介入において重要であることを示しています。しかし、多くの研究では、介入開始前の習慣的な食事摂取に関する情報が十分に得られていません (515)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 食事介入に対する個人の反応を予測することは進歩していますが、認知転帰への影響を予測する要因を調査する調査研究は不足しています。 微生物叢の組成 (民族性、年齢、性別、習慣的な食事パターンなど)、一貫したサンプル収集、配列決定技術、および他の「オミクス」アプローチの使用に影響を与えることが知られている定義済みの交絡因子を使用した、より大規模で強力な追加の臨床試験(トランスクリプトミクス、プロテオミクス、およびメタボロミクス) 微生物バイオマーカーを発見することは、食事介入に対する応答者と非応答者をさらに区別し、このアプローチを臨床ケアに適用できるようにするために必要です (495, 516)。 食事パターンが宿主の反応をどのように導くかについて明確なコンセンサスがまだないため、ベースラインの食事習慣の広範な特徴付けも重要です. たとえば、ベースラインでの食物繊維の摂取量が少ないと、プレバイオティック サプリメントに対する反応性が高まる可能性があることが提案されています。 それを使用して繁栄し、より強い宿主反応をもたらすか、または複雑な炭水化物を使用する細菌の酵素能力がないために反応性を低下させることができます(516。) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8.微生物叢は食事が脳と行動に及ぼす影響を調節していますか? | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
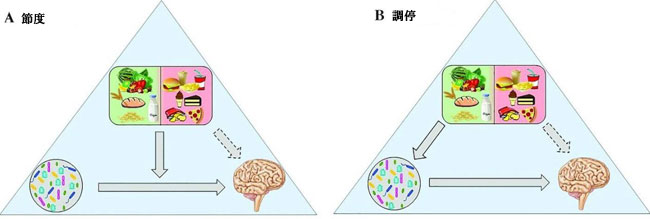
| 食事介入の有益な効果が微生物叢を介したものであるか、宿主に対する食事成分の直接的な影響によるものか、またはその2つの組み合わせによるものかは、現在の議論のトピックです。 微生物叢は、宿主の反応に対する食事の効果の仲介者または調整者である可能性があることが示唆されています (517)。 メディエーターとして、食事は微生物叢の組成と機能を直接変化させ、宿主に影響を与えますが、モデレーターとして、宿主の応答に対する食事の影響は微生物叢に依存しませんが、微生物叢は関係を強化する可能性があります (図 3)。 媒介効果の一例は、食物繊維の摂取と脳のエンドポイントです。 宿主がアクセスできない食物繊維は、腸内微生物叢によって発酵され、短鎖脂肪酸産生細菌の増殖と活動をサポートし、短鎖脂肪酸の産生を増加させます。 次に、短鎖脂肪酸は直接または間接的に脳にシグナルを送り、脳の生理機能と行動に影響を与えることができます。 一方、ω-3脂肪酸やポリフェノールの影響では緩和効果が見られました。 これらの栄養素は、それ自体が脳の生理機能の強力な調節因子であり、神経保護作用があります。 しかし、バイオアベイラビリティと一部の生物学的活性は、腸内微生物叢による変換にも依存している可能性があり (413)、これは、ポリフェノール、微生物叢、および関連する有益な効果の間の相互作用の場合に当てはまります (507)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 図 3 食事、微生物叢、および脳の間の相互作用を緩和することと比較した仲介 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 食事、微生物叢、および脳の間の緩和および仲介関係の両方が提案される可能性があります。 (A) 調節関係では、食事は微生物叢と脳の相互作用を強化または弱める可能性がありますが、(B) 仲介関係では、食事は微生物叢の組成と機能を直接変化させ、脳のプロセスに影響を与えます。 脳のプロセスに対する食事の潜在的な直接効果は、破線で描かれています。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 表 2に示すように、全食アプローチが腸内微生物叢および行動または認知の結果に与える影響を調査する研究は、食事 - 微生物叢 - 脳のクロストーク間の潜在的な相関関係または因果関係を解読するための証拠を提供し始めています。 相関分析は、微生物叢の食事誘発性の変化が、生化学的および行動の結果と遺伝子発現の変化と関連していることを明らかにした(314, 316)。 しかし、食事と脳の関係に対する微生物叢の潜在的な媒介効果に関する最も説得力のある証拠は、無菌マウスまたは抗生物質治療を使用した動物研究から得られます。 最近の研究では、微生物叢がケトジェニックダイエットの抗発作効果に必要であることが明らかになりました (341)。 著者らは、無菌状態または抗生物質治療がケトジェニックダイエットの抗発作効果を無効にしたことを示しただけでなく、ケトジェニックダイエットによって誘発された微生物叢の糞便移植も発作保護を誘発した。 同様に、糖尿病マウスモデルの認知障害に対する間欠的絶食の有益な効果は、抗生物質カクテルの投与後に部分的に無効になった(320)。 さらに、高脂肪食を与えられたマウスから従来のマウスに微生物叢を移植すると、レシピエントマウスが通常の固形飼料を与えられた場合でも、行動が変化し、神経炎症が増加しました(518)。 同様に、高フルクトース食によって誘発された海馬の神経炎症は、マウスに広域抗生物質を投与することによって抑制されました(342)。 最後に、高脂肪食を与えられたマウスの糞便ライセートを使用した in vitro 追跡調査の結果は、高脂肪食によって誘発されたグラム陰性菌の LPS 成分が、in vivo での神経細胞機能の障害に寄与したことを示唆しています(318)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 動物モデルでは食事効果における微生物叢の必要性の研究が可能ですが、これらの相互作用は臨床研究で解読するのがより困難です。 それにもかかわらず、人間のコホートにおける食事介入の成功は、ベースラインの微生物叢の組成に部分的に依存する可能性があるという事実は、人間の集団においてさえ食事の利点を媒介する上での微生物叢の重要性を示唆しています。 同様に、地中海式食事の抗炎症特性のいくつかは、微生物叢の調節によって媒介される可能性があることが以前に示唆されていました (519)。 微生物叢が健康転帰に対する食事の有益な効果を仲介していることを示す他の報告 (300、501、520) や、特に微生物叢を対象とした食事介入研究 (食物繊維や発酵食品の増加など) は、メンタルヘルスのいくつかの側面の改善を示しています(332) 。 同様の関係が人間のコホートの食事-微生物叢-脳の三角形で観察されるかもしれないという仮説を立てる理由を与えます。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9.結論と今後の方向性 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 微生物叢に対する食事の影響と、脳の機能と行動における微生物叢の重要な役割についての証拠が、文献に示されています。 前臨床研究は、食事と微生物叢と脳の相互作用を解明し始めていますが、これまでのところ、この複雑な関係を調査する人間の研究はほとんどありません. 上記で概説したように、ほとんどの研究は、動物モデルを使用した微生物叢と脳のインターフェースに対する高脂肪、高糖、または高カロリーの食事の有害な影響に焦点を当てており、食事–腸–脳軸と微生物叢の潜在的なメカニズムを理解し始めたばかりです。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 多くの異なる食事パターンが精神的健康の改善に関連しているという事実は、植物性食品が多く超加工食品が少ない全体的な食事パターンよりも、食事の個々の構成要素がメンタルヘルスにとって重要ではない可能性があるという事実を裏付けています。 ただし、植物を増やして超加工食品を減らすことの利点はすべてに当てはまりますが、研究では、個々の微生物叢の変動に影響される、特定の食品に対する個々の代謝反応の変動性が引き続き強調されています。 したがって、食事の特定の成分が腸内細菌叢にどのように影響し、メンタルヘルスを含む健康上の結果にどのように影響するかを理解することは、継続的に不可欠です。 食事と脳のプロセスの間のインターフェースにおける微生物叢の役割に関する証拠が出現しており、特に動物研究から説得力のある結果が得られていますが、この分野の研究はまだ始まったばかりであり、結果を過度に解釈しないように注意する必要があります。 同様に、動物モデルでの食事研究は、研究で使用される動物の食事製剤が人間集団の 1 日摂取量を超える用量を提供することが多いため、常に人間集団に翻訳できるとは限らないため、これらの研究の翻訳能力を考慮する必要があります。 微生物叢と脳のコミュニケーションを回復するための健康的な食事パターンの使用と、人間の介入に対するその有効性に関する多くの未回答の質問が残っており、脳に対する食事成分の直接的な影響を無視することはできません。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 微生物叢を標的とした人間の介入の開発を推進するためには、特定の介入に対する個人の反応を予測する決定要因をさらに理解することも重要です。 微生物の絶滅は、加工食品や揚げ物の消費の増加、繊維の摂取量の減少など、不健康な食事パターンに部分的に起因するという知識が増えていることを考えると、将来の栄養介入は、食事介入を効果的にするために、特別に設計されたプロバイオティクスと食事アプローチを組み合わせる可能性があります。 一部の研究では、低微生物叢アクセス可能な炭水化物食に続く何世代にもわたる微生物の多様性の喪失は、不足している微生物を投与した場合にのみ回復可能であることがすでに示唆されています。 したがって、不足している微生物は、必ずしもプロバイオティクス サプリメントによって供給される必要はありませんが、発酵食品などの有益な細菌を含む食品を通じて消費される可能性があります。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ヒトでの介入研究からの証拠は限られていますが、既存のデータは、植物性食品の摂取量と種類を増やし、超加工食品を減らすか排除することを一貫して支持しています。 この意味で、精神と腸の両方の健康に関する食事の推奨事項は、他のほとんどの健康状態に関する推奨事項と一致しています。 したがって、MyNewGutコンソーシアムは、うつ病患者に、穀物/繊維、発酵食品、および魚を多く含む植物ベースの食事を摂取するよう奨励することを推奨しています(521)。 さらに、プロバイオティクスをサプリメントとして使用することにはいくつかの証拠がありますが、食事を通じて微生物叢を操作することは、長期的にはプロバイオティクスのサプリメントよりも経済的に安価な解決策である可能性があります。 メンタルヘルスの治療戦略として食事の改善を採用することの追加の利点は、うつ病の症状を軽減する地中海スタイルの食事介入の有効性を調査する 2 つの画期的な試験の説得力のある経済的評価を考えると、その費用対効果です(522, 523)。 これは、一般的に精神障害と併存する慢性疾患を含む、全体的な健康と機能のための食事改善のプラスの利点に関連している可能性があります. 一般集団と比較した精神障害者の平均死亡率の差が 20 歳であることを考えると、身体的健康と精神的健康を改善する介入は、精神疾患に罹患した多くの人々の健康と精神的健康の両方の結果に大きな利益をもたらす可能性があります(138、524)。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 私たちの健康を改善する多くの機会は明らかに私たちの微生物叢にありますが、因果関係を確立し、「健康な微生物叢」を明確に定義し、個別化された栄養アプローチの可能性と限界を理解し、メカニズムの関係を解読するには、さらに研究を行う必要があります。 微生物群集の全体的な機能をよりよく理解するために、微生物叢のメンバーを特徴付けるだけでなく、マルチオミクスアプローチを統合することも重要です。 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| この文献は、Adv Nutr. 2021 Jul; 12(4): 1239–1285.に掲載されたDiet and the Microbiota–Gut–Brain Axis: Sowing the Seeds of Good Mental Health. を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |