Kayla Jaye et al., |
概要 |
| 近年、がんの抑制と進行に対する腸内微生物代謝産物の役割は、抗がん研究において大きな関心を集めています。腸内微生物叢は、腸内微生物代謝物を介して媒介されることが多いさまざまな種類のがんの発生、治療、および予後において極めて重要な役割を果たすことが確立されています。たとえば、バクテリオシン、短鎖脂肪酸、フェニルプロパノイド由来の代謝物を含む腸内微生物代謝物は、さまざまな分子メカニズムを介して直接的および間接的な抗がん活性を示しています。報告された抗がん活性にもかかわらず、二次胆汁酸を含むいくつかの腸内微生物代謝産物は、発がん促進特性を示しています。このレビューは、腸内微生物代謝産物の発がん性および抗がん活性を実証する現在の研究の重要な要約と評価を引き出し、これらの代謝産物と免疫系および腫瘍微小環境との相互作用を分子機構および臨床研究でさらに調査する必要性を強調しています。 |
| 目次(クリックして記事にアクセスできます) |
| 序章 |
| バクテリオシン |
| 短鎖脂肪酸 |
| フェニルプロパノイド由来代謝物 |
| プレニルフラボノイド |
| 天然プリンヌクレオシド |
| 二次胆汁酸 |
| 発がん性 |
| 抗がん作用 |
| まとめと今後の方向性 |
| 序章 |
| 過去 10 年間、腸内細菌叢は人間の健康維持に関連して広く調査されてきました。 腸内微生物叢は、腸の恒常性の調節と、腸の免疫にとって重要な上皮バリアの保存を通じて、宿主との相互に有益な関係を維持することが観察されています (1)。 これらの微生物は、宿主の自然免疫系の正常な生理学的機能と構造において重要であり、腸の健康に多くの影響を及ぼします(2)。 新たな証拠は、腸内微生物叢とその構造成分に加えて、腸内微生物群集によって生成される無数の代謝産物も、代謝反応のシグナル伝達分子および基質として作用することにより、宿主の生理学と健康に影響を与えることを示しています(3)。 私たちの最近のレビューでは、腫瘍に対する腸内微生物代謝産物の直接的および間接的な影響を強調しながら、最も一般的な5つのがんの予防、治療、および臨床転帰における腸内微生物叢の極めて重要な役割を強調しています(4)。 文献のいくつかの研究では、がんを含むさまざまな疾患における腸内微生物代謝物の潜在的な影響も実証されています。 このレビューは、がんの発生、予防、および治療における主要な腸内微生物代謝産物の役割を理解するために実施された現在の研究の重要な要約と評価を提供します。 このレビューはまた、がん研究のこの新興分野におけるいくつかの潜在的な将来の方向性を提案しています。 |
| 腸の代謝産物は、がんに対してさまざまな発がん促進効果および抗がん効果を示すことが示されています。 最近のレビューでは、大腸がん(CRC) の促進と予防における腸内代謝産物の二重の役割が示されました (5,6 )(図 1)。 また、大腸がんの進行を予測および予防するために、微生物代謝産物の累積的な影響を考慮する必要があることも示唆されています(5). さらに、大腸がんリスクの増加は、酪酸などの健康を促進する代謝産物と、二次胆汁酸 (BA) を含む潜在的に発がん性の代謝産物との間の不均衡によるものであると仮定されています(7)。 二次胆汁酸の発がん促進活性に加えて、微生物代謝は、農村部のアフリカ人やアフリカ系アメリカ人の集団で観察される結腸がんの進行に対する食事性脂肪 (肝臓での胆汁酸の合成を増加させる) の役割も浮き彫りにする可能性があります(7)。結腸の健康に対する食事の影響は、結腸の代謝プロファイルに対する高タンパク低炭水化物食の影響を評価した研究でさらに観察されました(8)。 著者は、高タンパクで低炭水化物の食事は、糞便中の抗がん代謝産物の観察可能な減少を引き起こすことにより、結腸の健康に有害な影響を与えることを報告しました。 発がん性代謝物の濃度が上昇し、長期にわたってこの食事を順守する個人の大腸がんのリスクが増加します(8)。 この傾向を裏付けるさらなる証拠は、健康に有益な短鎖脂肪酸(SCFA)を生成する炭水化物発酵と比較して、遠位結腸でのタンパク質発酵が有毒なアンモニア、アミン、フェノール、および硫化物を生成することにより宿主の健康に有害な影響を与えるという理解につながりました(9)。 . 2012 年の Windey らによるレビュー (9) では、肉が豊富な食事は、脂肪、ヘムおよび複素環式アミンの摂取量の増加に加えて、タンパク質の発酵を増加させることも示しました。 ただし、大腸がんに関連するタンパク質発酵と腸の健康との関係を理解するには、より詳細なメカニズム研究が必要です。 |
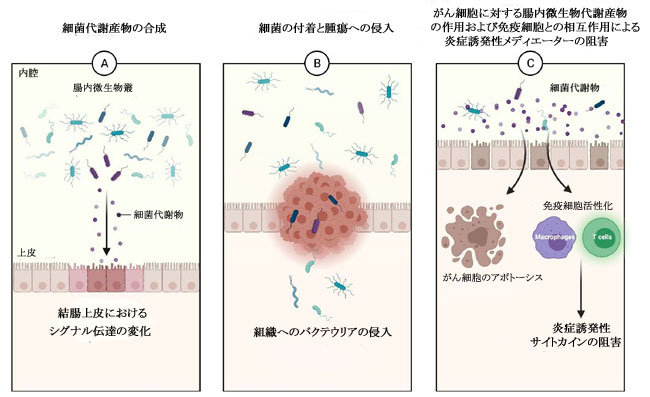 |
| 図1. (a) 結腸上皮に対する腸内微生物代謝産物のシグナル伝達効果、(b) 結腸直腸がん組織への細菌種の侵入の開始の概略図(6)。 さらに、(c) 腸内微生物代謝産物はがん細胞のアポトーシスを誘導します。 免疫細胞を活性化して、炎症誘発性サイトカインを阻害します(5)。 |
 |
| 図 2. 腸内微生物代謝産物と、子宮頸がん、乳がん、結腸がん、前立腺がん、頭頸部がん、および肝臓がんに対する作用の概要。 これらの代謝産物は、アポトーシスの誘導を介してがん細胞または腫瘍の増殖と生存を阻害します。 |
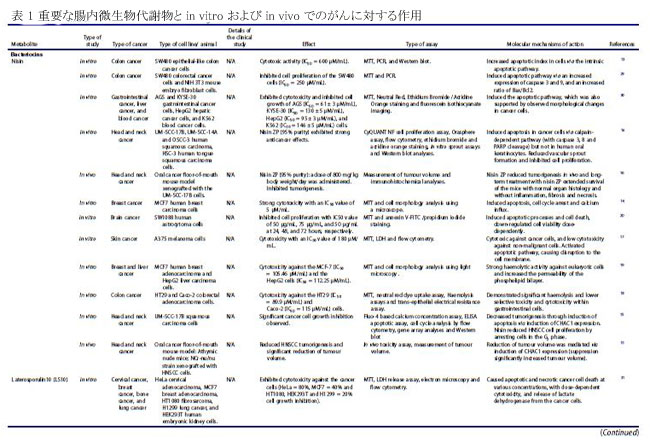
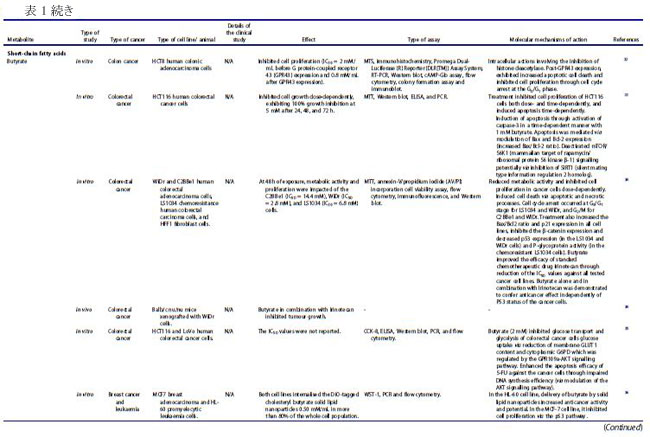
| 全身の健康を改善することを目的としたプレバイオティクスおよびプロバイオティクスベースの戦略により、腸内微生物群集による炭水化物発酵の副産物(二次代謝産物)である酪酸、酢酸、およびプロピオン酸を含む短鎖脂肪酸の役割に対する関心が高まっています(10)。 短鎖脂肪酸は食物繊維の発酵中にがんを保護する特性を示しますが、二次 胆汁酸は結腸内で生理的レベルが高くなると、結腸の炎症を誘発することで反対の効果を引き起こします(10,11)。 大量の二次胆汁酸は高脂肪食と相関しており、胆汁酸にさらされると活性酸素種が生成され、細胞膜とミトコンドリアが破壊される可能性があります(12)。 バクテリオシンなどの他の細菌代謝産物は、食品業界で食品保存料として安全に使用されており、結腸、頭頸部、乳房、脳、皮膚、および肝臓のがんに対する潜在的な治療薬として浮上しています(13–20)。 ナイシンは十分に研究されたバクテリオシンであり、in vitro および in vivo の両方で、アポトーシスの誘導を介して大腸がんおよび頭頸部扁平上皮がんに対する細胞毒性効果を示しています(13,16)。 報告されている潜在的ながん促進および抗がん作用にもかかわらず、がんの進行と予防における腸代謝物の役割をよりよく理解し、精密な抗がん療法を開発するには、さらなる動物および臨床研究が必要です。 |
| フェニルプロパノイドは、アミノ酸のフェニルアラニンとチロシンから合成される植物二次代謝産物の多様なファミリーです。 フェニルプロパノイド(訳者注:赤ワインのポリフェノール(タンニン)、ダイズのイソフラボンやブルーベリーのアントシアニン(フラボノイド)、ゴマのセサミン(リグナン)など、最近、いわゆる健康に良いとされる植物成分の多くがフェニルプロパノイドおよびその関連化合物です)からなる植物ベースの食事は、人間の健康を改善するために提案されています。 しかし、特定の微生物種は、フェニルアラニン、チロシン、トリプトファンの 3 つの芳香族アミノ酸 (AAA) を発酵させてフェニル酢酸 (PAA) にする能力を持っています。 および4-ヒドロキシルフェニル酢酸(4-ヒドロキシPAA)は、タンパク質発酵が結腸内のフェニルプロパノイド由来の副産物の可能性のある供給源であることを示しています(21)。 2013 年の Russell らによる研究( 21 )では、腸内微生物叢がタンパク質を発酵させて主要なフェニルプロパノイド由来の代謝産物を生成できることが示されましたが、2011年初めの同じ研究グループは、高タンパク質で低総炭水化物および繊維食は、糞便がん保護代謝物を大幅に減少させ、有害な代謝物の濃度を上昇させる可能性があることを示しました(8)。 |
| バクテリオシン |
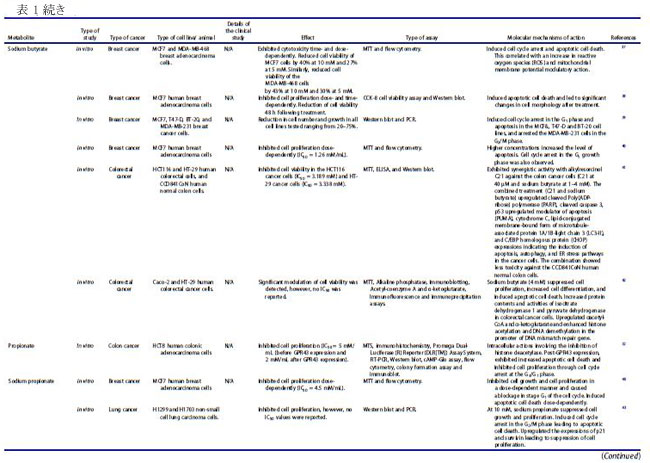
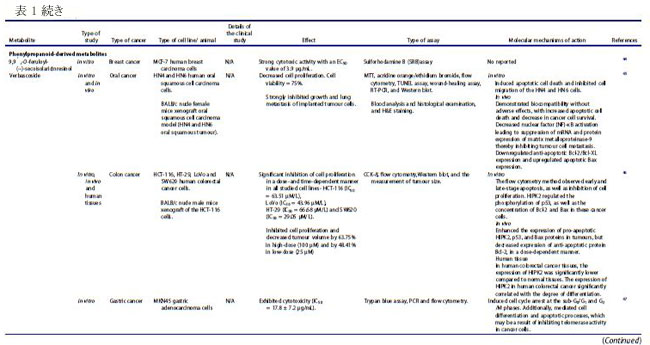
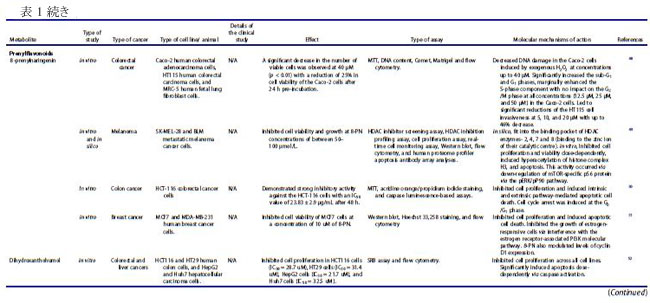
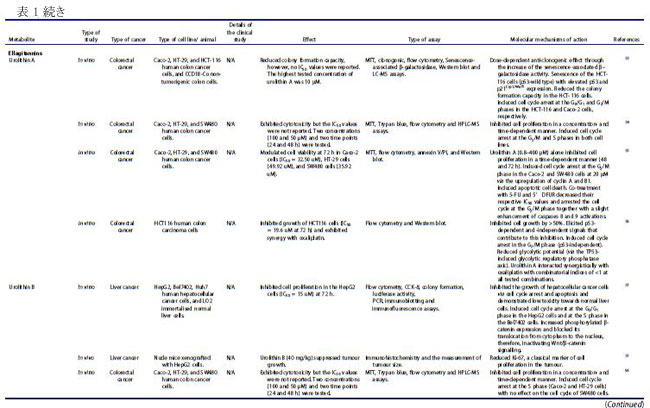
| バクテリオシンは、リボソーム活性を介して腸内の特定のプロバイオティクス細菌によって産生されるカチオン性ペプチドであり、構造的に細菌性抗菌ペプチドとして分類されます(22–24)。 バクテリオシンは、抗生物質耐性株などの他のバクテリアに対して、狭い範囲から広い範囲の活性で有意な阻害を示すことが実証されています(22)。 これらの抗菌ペプチドは、腸内の病原菌を阻害することもできるため (25)、腸の恒常性を維持する上で重要です。 乳酸菌は、バクテリオシンの最も重要な供給源の 1 つであり、特にFirmicutes門に属するEnterococcus属です(26,27)。 ある臨床研究では、大腸がん患者における病原性大腸菌株によるバクテリオシン産生の有病率がプロファイルされました(28)。 この臨床試験には、結腸直腸がん患者 30 人、結腸直腸腺腫患者 30 人、および健常対照者 20 人が含まれ、バクテリオシン - コリシン Ia、コリシン M、マイクロシン mH47、マイクロシン mV、およびマイクロシン mM を評価しました。 この研究では、進行期の 大腸がん患者は、より毒性の強い大腸菌株を示し、これは、進行期ではない診断と比較して、バクテリオシンの産生増加と相関していることが観察されました(28)。 バクテリオシンは、in vitro でがん細胞に対して有意な細胞毒性を示し (表 1)、正常な腸上皮細胞に対しては低い細胞毒性を示しました(18)。 さらに、バクテリオシンの抗がん活性は、腸内微生物組成の免疫調節と同様に、「定着抵抗性」として知られる現象で競合する病原性細菌株の定着を阻害する能力にも起因しています (図 3)。 細胞膜は真核細胞におけるバクテリオシンの主要な標的であり、バクテリオシンはがん細胞の負に荷電した細胞表面分子の発現を増加させ、細胞毒性を促進します(67)。 この活性の作用機序として提案されているのは、アポトーシス細胞死の誘導と、細胞膜の透過性の変化をもたらす細胞膜の脱分極であり、非受容体調節プロセスを示しています(67)。 がん細胞に対する選択的細胞毒性に加えて、バクテリオシンの非免疫原性および生分解性は、それらを新しい抗がん療法の有望な候補にします(67)。 したがって、腸内のプロバイオティクス細菌によるバクテリオシンの産生を調節する可能性と、それらのバイオエンジニアリングも、臨床応用のために提案されています(22)。 |
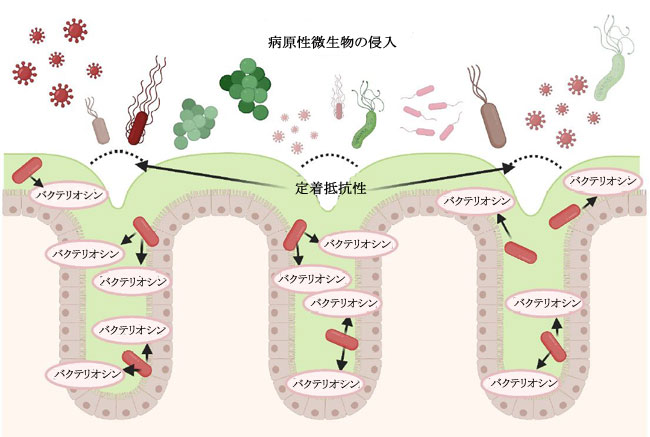 |
| 図 3. 乳酸菌が結腸上皮表面でバクテリオシンを産生し、病原性微生物の定着を防ぐことによって開始される「定着抵抗性」プロセスの簡略図。 |
 |
 |
 |
 |
 |
 |
 |
 |
| バクテリオシンは、宿主の免疫系を調節するだけでなく、腸内微生物叢に直接影響を与えることが示されているため、これらの代謝産物は、腸内での発がんを抑制するプロセス、および抗がん治療の有効性とがんの臨床転帰を決定する上で重要な役割を果たしている可能性があります。 しかし、バクテリオシンの抗がん効果に関する in vivo 研究は非常に限られており、ほとんどの研究は in vitro で行われています。 さらに、生存に関連するいくつかの制限が存在します。 in vivoでのバクテリオシンの機能は、a) 腸内での細菌株の生存、b) バクテリオシンの特異性、c) 使用する動物モデルの種類など、さまざまな要因にほとんど依存しているためです(66)。 これは、バクテリオシンが 腸管(GIT )内で合成されない、または大量に利用できない可能性があることを示唆する研究でさらに検証されましたが、既存の研究ではバクテリオシン生産の有効性に関して不確実性があります(68)。 バクテリオシンの生産と利用可能性との矛盾を考えると、微生物生態学の分析は、バクテリオシン治療の有効性を高めるための新しい創薬プロセスの不可欠な段階であるべきです(68)。 バクテリオシンのプロバイオティクス効果は、これらの代謝物を既存の抗生物質治療やファーマバイオティクスの新しい代替手段として組み込む可能性を実証しています(69)。 バクテリオシン種の特異性により、特定の病原体を確実に標的とすることができます。これは、従来の抗生物質、特に薬剤耐性の病原性株に取って代わる可能性があります(69)。 これを調査するために、かなりの数の胃腸がんの原因であるHelicobacter pylori菌種のさまざまな菌株(70) に対する乳酸菌によって産生されるバクテリオシンの抗菌活性を in vitro 研究で評価しました。 著者らは、Lactococcus lactis によって産生されるバクテリオシン ラクチシン(bacteriocins lacticin ) A164 および BH5 が、Helicobacter pylori菌株に対して最も実質的な抗菌活性を示したことを特定しました。 これらの発見は、バクテリオシンの阻害活性がHelicobacter pylori感染および関連疾患の制御におけるプロバイオティクスの使用を支持することを認めた別の研究によってさらに裏付けられました(71)。 バクテリオシンの構造的および機能的能力、ならびに免疫調節活性は十分に研究されていますが、腸系でのバクテリオシン産生を調節する要因をより包括的に理解するには、さらなる研究が必要です。 まとめて、利用可能なレポートに基づいて、バクテリオシンは、特にHelicobacter pyloriを含む発癌の原因となる微生物を標的とする、新しい治療法の開発に有望である可能性があります。 |
| ナイシンは、がん細胞に関するいくつかの研究で明らかなように、抗がんの可能性を持つ最も研究されているバクテリオシン (ランチビオティックとしても知られています) の 1 つです。 これまでのところ、L. lactis とS treptococcus uberis から、ナイシン-ナイシン A、Z、Q、および U の 4 つの変異体が発見されています(72)。 ナイシンは、Staphylococcus aureus,(黄色ブドウ球菌)やListeria monocytogenes(リステリア菌)を含む広範囲のグラム陽性菌に対する抗菌活性で知られる細菌発酵プロセスによって生成される多環式ペプチドであり(73)、結腸直腸がん細胞に対する潜在的な抗がんペプチドとして最近研究されています(13, 16,74)。 ナイシンの正常な生理学的機能は、細胞表面および他の細胞資源のコロニー形成について他のグラム陽性菌と競合することであり、乳製品の安全な食品保存料として広く使用されることに貢献しています(75)。 その安全性プロファイルにより、ナイシンは 50 カ国以上で食品防腐剤としての使用が承認されており、一般に世界保健機関によって人間にとって安全であると見なされています(67)。 最近の研究では、in vivo および in vitro の両方で、頭頸部扁平上皮がん細胞の腫瘍形成の阻害におけるその使用が調査されています(16)。 ナイシンは、固有のアポトーシス経路を介してがん細胞株のアポトーシス指数を増加させることが示されています(13)。 これは、特定のナイシンバリアントの濃度の増加が、アポトーシスによるがん細胞死のレベルの増加および頭頸部がん細胞の細胞増殖の減少と相関することを観察した別の研究によってさらに裏付けられました(16)。 ナイシンは、アポトーシス促進性陽イオン輸送調節因子であり、腫瘍形成に関連するアポトーシスメディエーターと考えられている CHAC1 (ChaC Glutathione Specific Gamma-Glutamylcyclotransferase 1) を介して、がん細胞に対してこれらの細胞傷害効果を及ぼすことが以前に提案されていました.15 ナイシン誘導性の CHAC1 発現は、カルシウム流入を増加させ、G2 期での細胞周期停止を誘導し、アポトーシスと腫瘍細胞増殖の減少をもたらしました (図 4)(15)。 しかし、著者らはまた、がん治療におけるナイシンの潜在的な使用のために最適な治療用量を決定しなければならないことも認めた(15)。 さらに、頭頸部扁平上皮がん細胞 (HNSCC) に対するナイシン ZP の関連性に関する in vivo 研究では、ナイシン ZP がマウスモデルの腫瘍形成を抑制し、ナイシン ZP による長期治療により、正常な臓器組織学を有するマウスの生存が延長されたことが実証されました(16 )(表 1)。 これは、ナイシンがCHAC1遺伝子発現のアップレギュレートによるアポトーシスの誘導を通じて頭頸部扁平上皮がん細胞の腫瘍形成を減少させた別のin vivo研究によってさらに裏付けられました(15)。 同じ研究は、80 μg/mL のナイシンが、24 時間後の G2 期における細胞周期停止を介して、UM-SCC-17B頭頸部扁平上皮がん細胞細胞の増殖を阻害することを示しました.(15)。 最近の研究では、3 つのバクテリオシンナイシン、エンテロシン、およびエピデルマイシンに由来する融合タンパク質を大腸菌でクローニングおよび発現させ、融合タンパク質を胃がんに利用する可能性を探っています(76)。 さらに、ナイシンは、表 1 に示すように、主にアポトーシスの誘導を通じて、血液、乳房、脳、結腸、胃腸、肝臓、および皮膚のがん細胞の増殖を前臨床的に阻害することが示されています。 これらの研究で示されているように、その有望な抗がん活性と安全性プロファイルにもかかわらず、ナイシンは単独で、または標準的な抗がん療法と組み合わせて臨床的に評価されていません. 正常細胞と比較して癌細胞に対する選択的な毒性があるため 67、ナイシンの治療の可能性を探るには、さらなる臨床研究を実施する必要があります。 現在の文献では、がん治療の代替アプローチとして、標準的な化学療法薬と組み合わせてバクテリオシンを使用する可能性が示されています。 最近の研究では、併用療法としてのナイシンと 5-FU の適用が、in vivo で 7,12-ジメチルベンズ(a)アントラセン誘発皮膚がんに対して相乗作用を示すことが確認されました。 また、A431 類表皮癌細胞に対する IC50 値を 8 倍低下させ、組み合わせ指数値は 0.188.77 でした。 併用療法の抗がん作用は、アポトーシス、血管新生、および細胞増殖経路の調節によって媒介され、その研究では単剤療法と比較して、腫瘍のサイズと数 (平均腫瘍体積と平均腫瘍量) が大幅に減少しました(77)。 さらに、食品および製薬業界では、バクテリオシンが抗生物質に取って代わり、多剤耐性病原体との闘いに役立つことが観察されています(24)。 このアプローチは、発がん性病原体を腸から排除するのにも役立つ可能性があります。 しかし、バクテリオシンの抗がん療法における単剤および標準化学療法との併用レジメンの有効性を理解するには、さらなる研究が必要です。 |
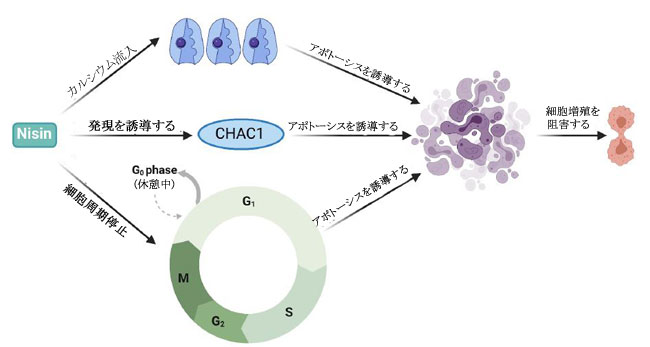 |
| 図 4.カルシウム分子の流入、アポトーシスメディエーターである CHAC1 陽イオン輸送レギュレーターの発現、細胞周期停止の誘導を含む、がん細胞株に対するナイシンの作用の分子メカニズムの図表示(15)。 |
| 短鎖脂肪酸 (SCFA) |
| 短鎖脂肪酸は、特に結腸がん細胞および乳がん細胞に対する阻害効果について、近年よく研究されています (表 1)。 食事の栄養因子が大腸がんの進行に重要な役割を果たしていることが確立されており、特定の因子が存在しないと、腸系内の代謝および恒常性経路が乱れ、腫瘍形成が促進される可能性があります(78–80)。 短鎖脂肪酸は食物繊維の分解に由来し、消化されない炭水化物の発酵が下部消化管系、盲腸および大腸で嫌気性盲腸および結腸微生物によって産生する(.81)。 この発酵プロセスにより、一次代謝産物として 短鎖脂肪酸を含む代謝産物のグループが生成されます.81 短鎖脂肪酸の産生に関与する優勢な細菌種は、Faecalibacterium prausnitzii、Clostridium leptum、Eubacterium rectale、および Roseburia 種、ならびに Anaerostipes 種および Eubacterium hallii を含む、乳酸と酢酸から 短鎖脂肪酸を合成する乳酸利用種です。 大腸がん患者に関するプロファイリング研究では、Bifidobacterium種の支配的なグループが消失し、非大腸がん参加者 (n = 14) と比較して大腸がん患者 (n = 14) に異なるスペクトルのBifidobacteriumが存在することが観察されました。 この観察結果は、非大腸がん群と比較して大腸がん患者の短鎖脂肪酸レベルが有意に低いことと相関していました(83)。 同じ研究グループは、大腸がん患者 (n = 14、インドネシア市民、18 歳以上) は、非大腸がん参加者 (n = 14、インドネシア市民、18 歳以上)、大腸がん発症の予防における短鎖脂肪酸の間接的な貢献を示している(84)。 いくつかの重要な短鎖脂肪酸は、嫌気性細菌と炭水化物の発酵によって合成される酪酸、酢酸、プロピオン酸など、結腸発がんに対する保護作用を示しています( 21,79,85,86 )(図 5)。 特に、繊維の消費による健康への主な利点は、結腸で起こる発酵プロセスによる短鎖脂肪酸の生成に起因すると考えられています(79)。 分子レベルでは、酪酸は細胞増殖を阻害し、がん細胞のヒストン過剰アセチル化の開始を通じてアポトーシスと細胞分化を誘導することが観察されています(5,78,86,87)。 さらに、短鎖脂肪酸は、結腸細胞やマクロファージや T 細胞などの免疫細胞の表面にある G タンパク質共役受容体 GPR41、GPR43、GPR109A を認識することができます。 これは、ヒストンの過剰アセチル化に加えて、結腸の制御性 T 細胞の総数と、抗炎症性サイトカインであるインターロイキン-10 (IL-10) およびトランスフォーミング増殖因子-β (TGFβ) のレベルを高めます (図 1)(5)。 |
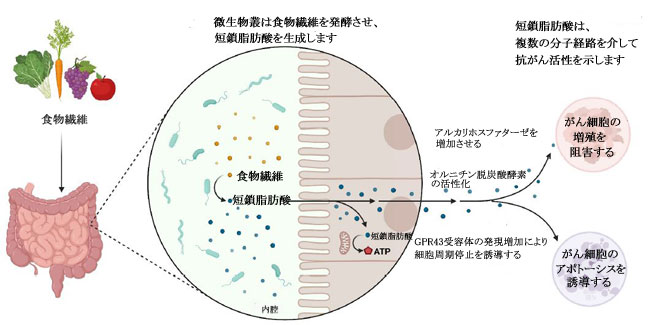 |
| 図 5. 腸内微生物種による食物繊維の発酵による短鎖脂肪酸(SCFA)の合成、およびさまざまな分子メカニズムによるこれらの代謝産物の抗がん作用の図式表示。 |
| ある研究では、HT29 ヒト結腸直腸腺がん細胞の増殖に対する 3 つの 短鎖脂肪酸すなわち酪酸、酢酸、およびプロピオン酸の効果が調査されました。 そして、酪酸とプロピオン酸は、観察可能な効果のない酢酸と比較して、HT29 細胞の増殖を阻害するのにより効果的であることを示しました(85)。 酪酸、プロピオン酸、および吉草酸 (短鎖脂肪酸の一種) がヒト結腸がん細胞を阻害し、酢酸およびカプロン酸 (短鎖脂肪酸の一種) については活性が報告されていない別の研究(86)で、同様の観察が行われました。 この研究の酪酸は、がん細胞のアポトーシスも有意に増加させました(86)。 がん細胞の増殖に対する酪酸とプロピオン酸の阻害効果は、ポリアミン代謝の重要な酵素であるオルニチン脱炭酸酵素の活性化と関連していましたが、これらの短鎖脂肪酸はポリアミン経路だけに作用するわけではないことも注目されました(85)。 その研究からのデータは、酪酸とプロピオン酸がアルカリホスファターゼ活性の増加を引き起こすことを明らかにし、それらが結腸の正常な生理機能において重要な役割を果たしている可能性があり、また結腸発癌に対する食物繊維の保護影響の寄与因子である可能性があることを示しています(85)。 短鎖脂肪酸、特に酪酸の腫瘍抑制機能は、不活性なプロカスパーゼ-3 を触媒的に活性なプロテアーゼ (アポトーシス) に変換するヒストン過剰アセチル化を介した経路によって引き起こされると考えられています (図 6) (32,86,87)。 この短鎖脂肪酸の抗腫瘍効果は、繊維が豊富な食生活が大腸がんリスクの減少と相関していることを強調する疫学的研究によっても支持されています(32,88)。 酪酸栄養補助食品のその他の健康上の利点には、高脂肪食によって引き起こされるインスリン抵抗性と肥満の予防が含まれます。これは、末梢組織の肥満の減少とインスリン感受性の増加によって達成されます(79)。 これは、栄養補助食品としての酪酸の投与が食事誘発性インスリン抵抗性を予防および治療することを観察したマウスモデル研究でさらに裏付けられました(89)。 著者らは、酪酸の作用機序がミトコンドリア活性の誘導とエネルギー消費の促進に直接関連していることも認めており、これは酪酸の抗がんメカニズムを理解する上で重要な進歩でした(89)。 |
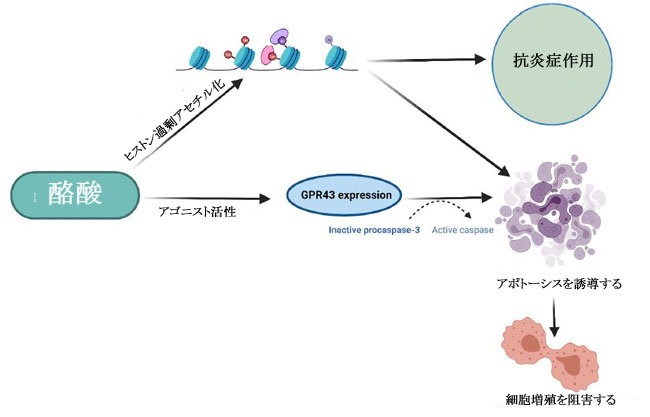 |
| 図 6. Gタンパク質共役受容体43(GPR43)および免疫細胞の調節を介したがん細胞に対する酪酸の生物学的活性の図式的描写(32,86,87)。 |
| 前述のように、細胞への酪酸の導入は、ヒストン脱アセチル化酵素活性の阻害を介してヒストンの過剰アセチル化を引き起こします(5,78,86,87)。 したがって、酪酸は、クロマチンの構造と機能におけるヒストンアセチル化の役割を決定する重要な要因として機能する可能性があり、がん細胞の増殖の防止と管理における酪酸とヒストンデアセチラーゼ阻害剤の強い関連性を示しています(90)。 この阻害活性の作用メカニズムも研究されており、GPR41/GPR43受容体シグナル伝達経路の短鎖脂肪酸媒介活性化とヒストン脱アセチル化酵素の阻害との間に密接な関連があることが示されました(91)。 GPR43 は 短鎖脂肪酸によって認識され、この受容体の発現は主に大腸および造血組織で見られ、結腸がん細胞株では頻繁に失われます(32)。 HCT8 ヒト結腸腺癌細胞における GPR43 受容体発現の回復は、G0/G1 細胞周期停止後のアポトーシス癌細胞死の増加につながることが確認されています(32)。 特に、HCT8 がん細胞を酪酸とプロピオン酸で処理すると、GPR43 受容体の発現とアポトーシスによる細胞死が増加しました (図 6)(32)。 したがって、GPR43 受容体は、大腸がんにおける短鎖脂肪酸のアポトーシス効果を媒介する機能的な腫瘍抑制因子として機能すると推測されています32,85,92,93。 これは、GPR43 受容体の酪酸とプロピオン酸の両方に対するアゴニスト活性を観察した別の研究によってさらに裏付けられました。この研究では、特定の病態生理学的条件下で免疫細胞において GPR43 の最高濃度が確認されました(92)。 別の報告では、短鎖脂肪酸は p21 欠失 HCT-116 結腸がん細胞に対して無効であるため、結腸がん細胞における短鎖脂肪酸を介した増殖停止には p21 遺伝子が必要であることが示唆されました。 正常な細胞では、p21 は細胞周期阻害剤および抗増殖エフェクターとして機能しますが、一部のがんでは調節不全になります(94)。 がんにおける p21 の役割は、p53 転写因子活性下の腫瘍抑制タンパク質としていくつかの報告で確立されています(94)。 近年、腸内細菌叢、免疫系、および食事要因の間の複雑な相互作用が研究されています(4)。 特定の代謝標的を含む、短鎖脂肪酸と T 細胞の免疫代謝との関連性を適切に判断するには、さらなる調査が必要ですが、短鎖脂肪酸が豊富な食事は、これらの二次代謝産物によるサイトカイン発現および T 細胞機能の調節を介して達成される可能性がある、T 細胞媒介性自己免疫応答に対する抑制作用を示すことが示されています(91)。 既存の研究で確認された酪酸の有望な抗腫瘍作用にもかかわらず、研究では大腸がんの発症におけるその前腫瘍効果も観察されています。 病原性細菌が大腸がんの発生に及ぼす影響を評価した研究では、酪酸の腫瘍促進効果または抗腫瘍効果は、治療への曝露の期間と量、および研究対象の細胞型を含む複数の要因に依存することが認められています(95)。 その研究は、「酪酸のパラドックス」という用語を作り出しました。酪酸の効果はその濃度によって決定され、低レベルでは腫瘍形成が促進され、高レベルでは腫瘍が阻害されます(95)。 別の研究では、マウスモデルを利用して「酪酸のパラドックス」を検証し、酪酸の明らかな抗腫瘍活性にもかかわらず、低濃度の酪酸が結腸上皮細胞の増殖を増加させることにより大腸がんの発症を促進することを観察しました(5)。 多くの研究で、動物とヒトにおける酪酸の同様の発がん効果が観察されていますが、宿主の遺伝学、微生物組成、およびこの逆説的な効果を構成する他の腸代謝産物の存在の間の相互作用を理解するには、さらなる調査が必要です(96)。 低レベルの酪酸は、潜在的な病原体を抑制し、酪酸産生種によるコロニー形成を促進することにより、腸内微生物組成を破壊する宿主内の炎症誘発性環境を開始することも示されています(5)。 濃度に依存する酪酸活性の複雑さは、潜在的な抗がん治療薬としての使用において重要な考慮事項です。 標準的な化学療法薬に関連して、ある研究では、酪酸が結腸がん細胞に対する 5-フルオロウラシル (5-FU) の有効性を大幅に改善し、5-FU によって引き起こされる DNA 合成の障害が増加したことが確認されました(97)。 並行した観察が、標準的な化学療法薬を用いてコンジュゲート標的送達システムを形成することを報告した以前の研究で行われました。 ドキソルビシンのようなものであり、短鎖脂肪酸は標準的な薬剤の有効性を改善し、薬剤耐性の発生を制限し、より効率的に腫瘍の微小環境を標的にすることができます(98)。 酪酸は、ヒストン脱アセチル化酵素 (HDAC) の阻害を通じて、標準的な化学療法の臨床効果を改善し、毒性を軽減するのに有益である可能性があります(99)。 |
| 全体として、短鎖脂肪酸は特に結腸がんの状況で有望です。 今後の研究では、膵臓がんや胃がんを含む他の種類のがんに対する短鎖脂肪酸の効果を評価して、それらの分子作用機序を理解する必要があります。 研究では、標準的な化学療法の有効性と安全性、およびがんの予後に対する 短鎖脂肪酸の影響も調査する必要があります。 |
| フェニルプロパノイド由来代謝物 |
| フェノール酸などのフェニルプロパノイド由来の代謝産物は、植物の二次代謝の重要な構成要素であり、いくつかの研究で、多くの分子経路を介してさまざまながん細胞タイプの増殖を阻害することが実証されています(100–103)。 植物の二次代謝の誘導体および植物性食品が豊富な食事の成分として、フェニルプロパノイド(訳者注:赤ワインのポリフェノール(タンニン)、ダイズのイソフラボンやブルーベリーのアントシアニン(フラボノイド)、ゴマのセサミン(リグナン)など、最近、いわゆる健康に良いとされる植物成分の多くがフェニルプロパノイドおよびその関連化合物です)は宿主において化学予防、抗酸化、抗炎症、および抗有糸分裂活性を示しました(104–106)。 フェニルプロパノイドの生合成は、これらの二次代謝産物の発生に関与する上流および下流の酵素の科学的理解を深めるために、過去 10 年間に広く研究されてきました(104)。 特殊なフェニルプロパノイド製品は、フェニルプロパノイド代謝酵素の認識された機構的基盤から開発できると推測されています(104)。 ヒトの糞便サンプルに関するある研究では、フェニル酢酸と 4-ヒドロキシルフェニル酢酸 (検出された 2 つの最も豊富な代謝産物) を含むフェニルプロパノイド由来の化合物が、植物が豊富な食事と結腸内の芳香族アミノ酸の微生物発酵の両方から合成されることがわかりました(21)。 特に、フェニル酢酸 (PAA) や 4-ヒドロキシルフェニル酢酸 (4-ヒドロキシPAA) などのフェニルプロパノイド由来の化合物は、Bacteroidetes (Bacteroides thetaiotaomicron、Bacteroides egerthii、Bacteroides ovatus、Bacteroides fragilis、Parabacteroides distasonis)、および Firmicutes (Eubacterium hallii および Clostridium bartlettii) による結腸での芳香族アミノ酸- フェニルアラニン、チロシン、トリプトファンの微生物発酵によって生成されます。 結腸直腸癌患者のプロファイリング研究では、診断が進んだ患者(ステージ III および IV CRC)で Bacteroides fragilis レベルの上昇が検出され、これらのレベルは直腸よりも結腸で高かった(107)。 この研究には、2 つのコホート (平均年齢 59 歳、性別は均等) からの患者が参加しました。 最初のコホートは、大腸がんに加えて事前に選択された状態のない 55 対の 大腸がん患者サンプルで構成され、2 番目のコホートは、散発性マイクロサテライト不安定性と診断された 18 人の患者で構成されていました(107)。 フェニルプロパノイド由来の代謝産物の産生に関与する細菌の存在の増加は、腫瘍の高度な進行と抗腫瘍バクテリアおよび代謝産物産生の増加の間に潜在的な因果関係を示していた(107)。 Lactobacillus rhamnosus のプロバイオティクス代謝物に関する in vitro 研究では、4-ヒドロキシルフェニル酢酸がミトコンドリア調節アポトーシスを活性化し、HepG2 肝癌細胞株に対する細胞増殖を誘導することが観察されました(108)。 同様に、in vitro 研究では、フェニル酢酸と 4,4'-ビピリジン リガンドを組み合わせた新規の Zn(II) 錯体の活性が調べられ、この錯体は、アポトーシスの誘導を通じて HeLa 子宮頸がん細胞株を阻害することが確認されました(109)。 フェニルプロパノイドは、微生物の多様性と生合成起源でよく知られていますが、これらの化合物の作用メカニズムと抗菌活性、特に発がん性細菌やウイルスに対する抗菌活性をよりよく理解するには、さらなる研究が必要です(110)。 フェニルプロパノイド化合物である食事中のベルバスコシドは、その抗菌性と抗酸化性により、腸の形態に影響を与えることが示されています(111)。 著者らは、ベルバスコシドが消化管の味覚受容体のタンパク質発現を調節することにより、食欲を刺激する可能性のある酸化ストレスから消化管を保護することを実証しました(111)。 ベルバスコシドは、細胞周期のサブ G1 期および G2/M 期で MKN45 胃上皮がん細胞を停止させることもわかっています (図 7)。 |
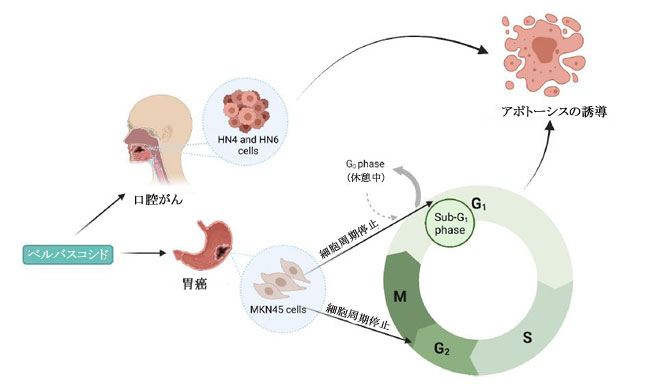 |
| 図 7. HN4およびHN6ヒト口腔扁平上皮がんおよびMKN45胃上皮がん細胞に対する、腸内微生物代謝産物であるベルバスコシドの作用の分子メカニズムの概略図(45,47) |
| ある研究では、ヒトとラットの腸内微生物叢が、ベルバスコシドの一種であるアクテオシドを、8つの分解代謝物、腸内細菌および腸酵素サンプル中の2つの異性体、および4つの親代謝物を含む14の代謝物に分解できることが示されました(112)。 アクテオシドはまた、濃度依存的にマウス腹腔マクロファージの リポ多糖誘発 PGE2、一酸化窒素、および TNF-α を阻害することにより、顕著な抗炎症効果を示しました(113)。 これらの化合物の抗炎症効果を理解するにはさらなる研究が必要ですが、ポリフェノールは複数の炎症成分を標的とし、炎症誘発性サイトカインの合成、免疫細胞の調節、および遺伝子発現を介して免疫プロセスを調節することが示唆されています(114)。 これらの生物学的に活性な化合物は、前臨床実験モデルと臨床研究で観察されたように、免疫系をサポートし、慢性疾患の発症を防ぐことにより、がんを含むいくつかの慢性炎症性疾患の健康上の利点を促進します(114)。 近年、研究は、最も費用対効果が高く、副作用が少ない天然物質の使用に焦点を当てることを優先してきました(115)。 既存の研究では、がんの治療におけるフェニルプロパノイドと標準的な化学療法剤の併用も評価されています。 HeLa 子宮頸がん細胞に対する 5-FU と組み合わせた 8 つの異なるフェニルプロパノイドの複合治療の可能性を評価することを目的とした in vitro 研究では、オイゲノール、フェルラ酸、およびカフェイン酸が 5-FU と組み合わせた場合に相乗効果を示すことが確認されました(116)。 さらに、フェニルプロパノイドは、ヒト赤血球に対して最小限の溶血活性を示し、宿主内で毒性を引き起こすことなく、これらの化合物を医薬品として使用することを支持しています(116)。 がんに対する広範な生物学的特性と腸内微生物叢の調節にもかかわらず、腸内微生物叢によるフェニルプロパノイドの代謝および異化経路と、これらの化合物が炎症および微生物プロセスを調節するメカニズムを理解するには、さらなる研究が必要です(117)。 |
| プレニルフラボノイド |
| キサントフモール (XN) は、ホップに含まれるプレニル化フラボノイドであり、近年有望な抗がん作用を示しています(52)。 腸内微生物叢はキサントフモールを代謝して、非常に強力な植物性エストロゲンである 8-プレニルナリンゲニン (8-PN) を生成することができます(52)。 主に、8-PN は複数のがん細胞株にわたって強力な抗がん作用を示し (表 1)、他の健康上の利点が認められています(49,118)。 両方の化合物が末梢血単核細胞の細胞生存率の増加に同様の効果があることを示す臨床試験にもかかわらず、8-PN の生物活性は、その異性体 6-PN よりも健康な個人での経口バイオアベイラビリティが高いことに起因する可能性があります(118)。 ある研究では、SK-MEL-28 および BLM ヒト転移性メラノーマ細胞に対する 8-PN の抗がん作用が in silico および in vitro で評価され、ヒストン脱アセチル化酵素の阻害を介して抗がん作用を媒介することが観察されました(49)。 以前の研究では、8-PN がアポトーシスの誘導を通じて MCF7 ヒト乳がん細胞の増殖を阻害したことも示されました(51)。 この活性の提案された作用機序は、エストロゲン受容体関連の PI3K 経路との干渉による、8-PN 代謝産物によるエストロゲン応答性細胞の増殖の増加でした(51)。 8-PN の強力なエストロゲン様作用は、ゲニステイン、ダイゼイン、およびクメストロールを含む他の確立された植物性エストロゲンよりも高い活性を示した他の研究でも観察されました(119)。 別の研究では、8-PN の抗がん性とアポトーシスの可能性をプレニルフラバノンの他の側鎖バリアントと比較し、8-PN が多剤耐性白血病細胞を標的にし、ミトコンドリア依存性アポトーシスを誘導できることを発見しました(120)。 結腸直腸腫瘍形成のさまざまな段階に対する 8-PN の in vitro 作用も調査されました。 この代謝産物は、用量依存的に HT115 細胞の細胞増殖を阻害し、未処理の対照と比較して、最大 46% の成長低下を示しました(48)。 著者らは、8-PN が結腸直腸腫瘍形成のさまざまな重要な段階で抗がん活性を発揮し、大腸がんの予後不良を改善するのに有益である可能性があることを実証しました(48)。 より最近の研究では、HCT-116 結腸がん細胞に対する 8-PN の抗がん活性を具体的に評価し、それが外因性および内因性経路媒介アポトーシスの誘導を介して抗増殖活性を付与することを決定しました(50)。 8-PN の抗癌活性に関するこれらの in vitro での知見は、その作用機序と天然の抗がん剤としての潜在的な用途を調べるためのさらなる in vivo および臨床研究を正当化します。 |
| 天然プリンヌクレオシド |
| 天然のプリンヌクレオシドは、さまざまな分子メカニズムを介して抗がんの可能性を示しています。 ヌクレオシドベースの抗がん剤の有効性は、細胞内外への薬物の移動を調節する細胞トランスポーターによって決定されます(121)。 さまざまな寄生生物種の分析により、アデノシン トランスポーターとイノシン トランスポーターという 2 つの顕著なプリン ヌクレオシド トランスポーターが特定されました(122)。 イノシンは、アデノシン化合物の異化作用によって合成される代謝産物であり、アデノシン受容体に直接作用することにより、生体内で多様な抗炎症および免疫調節効果を示しました(123)。 これらの効果の作用の調節機構として提案されているのは、アデノシン A2A 受容体 (A2AR) によるものであり、A2AR のイノシン調節活性化が cAMP 産生と、キナーゼ 1 および -2.123 の細胞外シグナル調節リン酸化を開始します。 in vivo 研究の結果は、イノシンがアゴニストとして ERK1/2 バイアス シグナル伝達を開始し、A2AR 活性化を増幅および拡張できることを確認しており、これには重要な薬理学的意味があります(123)。 イノシン代謝産物に関する別の in vivo 研究では、チェックポイント遮断免疫調節の効果を増幅しながら、結腸直腸、膀胱、および黒色腫のがんタイプにおける T 細胞の抗腫瘍活性を増強することが認められました.58 著者らはまた、ヒトの免疫チェックポイント遮断 (ICB) 療法に対する反応性に関連する細菌 Akkermansia muciniphila が、その ICB 促進効果のためにイノシン-A2AR シグナル伝達を使用していることを確認しました(58)。 膀胱癌に対するイノシンの提案された作用機序は、併用療法としてCpG(メチル化されていないデオキシシチジリル-デオキシグアノシンジヌクレオチド124を含むDNAオリゴヌクレオチド)と一緒に投与した場合(58)、腫瘍全体の重量を減らすだけでなく、抗 CTLA-4 の機能を強化して、IFN-ɣ+CD4+ および IFN-ɣ+CD8+ T 細胞の腫瘍への浸潤を増加させることでした。 この研究では、Bifidobacterium pseudolongum によって産生されるイノシンが cDC 依存性の TH1 細胞回路の活性化を増加させ、腸および上皮腫瘍のマウスモデルにおける ICB 療法の全体的な効果を高めたことも報告されています.58 イノシンは、他の免疫療法と組み合わせて有望な抗がん作用を示していますが、宿主内での単独の抗がん活性と作用機序については、さらなる調査が必要です。 |
| 二次胆汁酸 |
| 発がん性 |
| 二次胆汁酸は、大腸の嫌気性細菌による一次胆汁酸の脱ヒドロキシル化から代謝されます。一次胆汁酸は、腸系に放出される前に肝臓肝細胞のコレステロールから最初に合成されます(125,126)。 主に二次胆汁酸の産生に関与する細菌種は、C. scindens、C. hiranonis、C. hylemonae、および C. sordellii を含む Clostridium 属のメンバーです(127)。 短鎖脂肪酸とは異なり、胆汁酸は発がん促進活性を示すことが示されています。 研究によると、胃腸 (GIT) 管細胞が高レベルの二次胆汁酸にさらされることは、胃腸がんの発症につながる主要な危険因子であり、高レベルの胆汁酸は、食事による脂肪の摂取量が多い人に最も一般的に見られることが示されています(.12,125,128,129)。 高脂肪食は、デオキシコール酸 (DCA) やリトコール酸 (LCA) を含む腸肝循環の二次胆汁酸のレベルの増加につながり、どちらも結腸の炎症やがんの誘発の危険因子となる可能性があります(11)。 この二次胆汁酸、特に デオキシコール酸の蓄積は、ヒトの肝臓が門脈を介して 7α-ヒドロキシル化二次胆汁酸を戻すことができないためであり、「西洋食」を摂取しているヒトに高レベルの蓄積を引き起こします(127)。 デオキシコール酸が、さまざまな疾患の発症に関与する細胞シグナル伝達経路を開始できることは十分に確立されています(127)。 これを裏付けるために、大腸がん患者の代謝物濃度をプロファイリングする臨床研究では、参加者の糞便、血清、および胆汁中のデオキシコール酸レベルの上昇が観察されました(130)。 同様に、高脂肪食を与えられたマウスは、他の微生物種と比較してより高いレベルの C. sordellii を示したことも報告されており、デオキシコール酸レベルの上昇におけるその役割を示唆しています(127,131)。 胆汁酸への過剰な曝露は、さらに 活性酸素種の生成につながり、続いてミトコンドリアと細胞膜が破壊され、DNA が損傷する可能性があります(12)。 細胞を 胆汁酸に長時間さらすことによって引き起こされる酸化的および DNA 損傷関連のストレスは、細胞内のゲノム不安定性を開始し、アポトーシス耐性の発達と最終的ながんの発症につながります(12,129)。 核内受容体は二次胆汁酸の作用を促進する腫瘍からの保護において転写因子として作用するため、胆汁酸代謝と解毒の調節に直接関連していることが報告されています(132)。 これは、二次胆汁酸がWnt/β-カテニンシグナル伝達経路の開始を介して腸腺腫-腺癌シーケンスを加速する可能性がある、ヒトの遺伝子突然変異素因の状況における重要な考慮事項です(133)。 二次胆汁酸の発がん促進活性に関連する遺伝的および環境的要因は、これらの代謝産物ががんの起源と予後において果たす役割をさらに調査するための基盤を提供します。 |
| 脂肪分の多い食事をしている個人に見られる一般的な二次性胆汁酸として、デオキシコール酸は 大腸がんの発症における重要な環境トリガーとして機能します(134)。 腸の腫瘍形成に対するデオキシコール酸の正確な作用機序についてはさらなる調査が必要ですが、デオキシコール酸が腸の粘膜バリアを破壊し、腸内の炎症誘発性サイトカインの産生を増加させることが観察されています。 腸内微生物群集は、腸の恒常性の調節に本質的に関与しており、腸内の高濃度のデオキシコール酸によって誘発される腸の腫瘍形成と腸内細菌叢の異常が直接関連しています(133)。 in vivo 研究では、デオキシコール酸で処理されたマウスは、腸バリアの障害、炎症プロセス、および腫瘍形成と相まって(133)、腸内細菌叢の組成に変化が見られました。 その研究の発見は、腸へのデオキシコール酸の導入が腸内微生物組成の乱れを引き起こし、腸の発がんを促進したという証拠を提供しました。 これらの発見は、他の利用可能な研究と一致して、二次胆汁酸と腸内微生物叢との間の相互作用が腸の発癌の開始に関与していることを強調しており、これは特に胃腸がん新しい治療戦略を開発するために重要です(133,135)。 結腸粘膜の過剰増殖は、結腸直腸発がんの進行の前段階であり、動物モデルでは、デオキシコール酸が腫瘍促進活性で結腸直腸上皮細胞の増殖を刺激することが観察されました(136)。 別の研究では、Caco-2 ヒト結腸がん細胞の遊走に対する デオキシコール酸の効果が評価され、がん細胞の遊走はプロテイン キナーゼ C と関連している可能性が高いことがわかりました。 デオキシコール酸の腫瘍プロモーター活性は、デオキシコール酸が BHK-21 線維芽細胞の原形質膜への Ca2+ 流入の増加を介してプロテインキナーゼ C およびホスホリパーゼ C を活性化できることを示す別の研究でさらに検証されました.138 1999 年の結腸癌の有無にかかわらず 19 人の患者に関する生検研究では、結腸直腸増殖の有意な増加が血清デオキシコール酸レベルと相関していたが、リトコール酸、コール酸、ケノデオキシコール酸、およびウルソデオキシコール酸などの他の 胆汁酸の血清レベルとは相関していないことが示された.136 デオキシコール酸レベルの低下が発がんリスクを低下させるかどうかを調べるには、より多くのヒト介入研究が賢明です. アメリカ人の大腸がんのリスクが高いのは、高脂肪・高タンパクの食事が一因である可能性が高く、発がん性の可能性がある二次胆汁酸を生成する可能性のある微生物種の促進につながることが実証されています(80)。 同様に、より最近の調査では、複数の作用機序と、胆汁酸およびその誘導体による大腸がん発生の促進に関与する多様なシグナルが存在することが確認されており、大腸がんの予防において一次および二次胆汁酸を標的とする可能性が強調されています(139)。 ただし、これまでのところ、これらのメカニズムとシグナル伝達経路を調査する研究は限られているため、がんの予防と治療を支援するための胆汁酸に関する将来の前臨床および臨床研究の必要性が強調されています。 |
| 抗がん作用 |
| 二次胆汁酸の主な発がん性にもかかわらず、コール酸の誘導体であるリトコール酸(LCA)は、さまざまながん細胞株に対して抗増殖作用を示しました(表1)(.61–64,140)。 ヒト乳腺がん(MCF-7 および MDA-MB-231)、ヒト前立腺がん(PC-3、LNCaP および DU-145)、肝がん(HepG2) および神経芽腫がん (BE(2)-m17) に関する in vitro 研究、SK-n-SH、SK-n-MCIXC および Lan-1) 細胞は、リトコール酸が異なる分子メカニズムによってがん細胞の増殖を阻害することを発見しました (表 1)(59–61,63,64)。 特に、リトコール酸は、カスパーゼ 3、8、および 9 を介したアポトーシスを介して、ヒト前立腺がん (LNCaP および PC-3) 細胞の増殖を抑制しました(60)。 リトコール酸はまた、HepG2 肝がん細胞において、小胞体 (ER) ストレスおよびトランスフォーミング増殖因子-β (肝細胞のアポトーシスおよび肝線維症を誘導する強力な線維形成促進因子) を誘導することができた(59)。 HepG2 肝がん細胞の細胞毒性と小胞体ストレスは、二次胆汁酸の疎水性に大きく依存しており、試験した二次胆汁酸の中でケノデオキシコール (疎水性の二次小胞体) が最大の活性を示しました(59)。 別の研究 62 は、リトコール酸が in vitro (MCF7、4T1、および SKBR3 細胞に対して、初代線維芽細胞に影響を与えずに) および in vivo (4T1 異種移植雌 BALB/c マウス) で乳癌細胞に対して抗増殖性であることを示しました。 酸化的リン酸化とTCAサイクルを誘導し、上皮間葉転換、血管内皮増殖因子Aの発現を阻害し、抗腫瘍免疫を高めました。 リトコール酸の作用の分子メカニズムは、TGR5 受容体を介したものであることがわかった. 一部の二次胆汁酸の抗がん活性に関するこれらの新しい発見にもかかわらず、がん細胞におけるミトコンドリア機能障害と細胞毒性の詳細なメカニズムはまだ確認されておらず、さらなる調査が必要です。 |
| まとめと今後の方向性 |
| 腸の恒常性と免疫の健康の維持における腸内微生物叢の重要な役割は、文献で十分に文書化されています。 この認識は、特にがんに関連して、腸内微生物代謝産物が人間の健康に及ぼす影響の調査につながりました。 特定の腸内微生物代謝産物とがん細胞増殖の進行または阻害との間の複雑なクロストークは、抗がん研究の新たな領域です。 in vitro および in vivo で実施された研究は、宿主におけるこれらの腸代謝産物の抗がん作用およびがん促進作用を理解する上で非常に重要です。 たとえば、短鎖脂肪酸やバクテリオシンなどの腸内代謝物は、さまざまな種類のがん細胞に対して細胞傷害活性を有することがますます報告されています。 短鎖脂肪酸は、がん細胞の増殖を抑制するのに役立つ、宿主の健康に対する抗酸化作用と抗炎症作用で一般的に知られています。 さらに、短鎖脂肪酸は、結腸直腸がんの発症において重要な考慮事項である、肥満によるインスリン抵抗性の抑制において重要な役割を果たしていることが示されています。 同様に、バクテリオシンは、直接的(がん細胞のアポトーシス経路の誘導による)および間接的(病原性微生物を阻害するために上皮表面にコロニー形成抵抗性を付与することによる)作用を介して抗腫瘍の可能性を示しました。 さらに、フェニルプロパノイド由来の代謝産物は、植物が豊富な食事を摂取する個人で高濃度で観察され、抗酸化、抗炎症、およびがん細胞増殖に対する阻害活性について、過去 10 年間にわたって広く研究されてきました。 比較的、二次胆汁酸類は、宿主で発がん促進活性を発現することがわかっています。 これらの有毒な副産物は、細胞内で遺伝的不安定性を引き起こし、がん細胞の増殖を増加させるアポトーシス耐性を助長する可能性があります。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
| この文献は、Gut Microbes. 2022; 14(1): 2038865.に掲載されたThe role of key gut microbial metabolites in the development and treatment of cancer.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |