Jiezhong Chen et al., |
概要 |
| ほとんどのCOVID-19症例は軽度または無症候性であり、よく回復します。これは、効果的な免疫応答が起こり、SARS-CoV-2ウイルスを首尾よく排除することを示唆しています。ただし、患者のごく一部は、病理学的免疫応答を伴う重度のCOVID-19を発症します。これは、抗炎症メカニズムとバランスの取れた強力な免疫システムがSARS-CoV-2感染からの回復に重要であることを示しています。現在、SARS-CoV-2に対する多くのワクチンが開発されており、SARS-CoV-2感染から保護したり、感染した場合の病気の重症度を軽減したりするための効果的な免疫応答を誘発します。まれではありますが、深刻な罹患率と死亡率は、COVID-19ワクチンの副作用と有効性の欠如の両方に起因しており、有効性のさらなる改善と悪影響の予防が緊急に保証されています。多くの要因が、最適な免疫応答を達成するためのこれらのワクチンの有効性に影響を与える可能性があります。腸内細菌叢は効果的な免疫系応答の発達と維持に関連しているため、腸内細菌叢の調節不全(腸内毒素症)は重要な危険因子である可能性があります。 このナラティブレビューでは、SARS-CoV-2に対する免疫応答、COVID-19ワクチンが防御免疫応答を誘発する方法、COVID-19ワクチンの非効率性と悪影響への腸内細菌叢の関与、およびCOVID-19ワクチンの免疫を改善するため機能性食品による腸内細菌叢の調節について説明します。 |
| 目次(クリックして記事にアクセスできます) |
| 1. はじめに |
| 2. 免疫および腸内細菌 |
| 3. SARS-Cov-2感染に対する免疫応答 |
| 4. 腸内毒素症によるワクチンとメカニズムの非効率性 |
| 5. 危険因子をCOVID-19ワクチンの非効率性に関連付ける腸内毒素症 |
| 6. COVID-19ワクチンの有効性 |
| 6.1 mRNAワクチン |
| 6.2 アデノウイルスベースのワクチン |
| 6.3 不活化または弱毒化ウイルスワクチン |
| 6.4 COVID-19から回復した患者のワクチン接種 |
| 6.5 ワクチン有効性に対する変異体の影響 |
| 7.COVID-19ワクチンの副作用 |
| 8. COVID-19ワクチン接種のための腸内細菌叢を改善するためのアプローチ |
| 9. 結論 |
1.はじめに |
| 重症急性呼吸器症候群コロナウイルス2(SARS-CoV-2)感染によって引き起こされるCOVID-19のパンデミックは、世界的に大きな医学的問題です。感染症と戦うための現在の特定の抗ウイルス薬はありません。ウイルスの蔓延を防ぐためのワクチンは、パンデミックのさらなる拡大を阻止するための最も効果的なアプローチになっています。現在、多数のCOVID-19ワクチンが開発されているか、開発中であり、多数の研究が進行中です(Clinical-Trials-Org、2020–2021)。ほとんどのワクチンは前臨床段階にありますが、11のCOVID-19ワクチンが世界中で緊急使用が承認されています(Vasireddy et al.,2021)。一般に、これらのワクチンの適用により、米国やイスラエルなど、ワクチン接種を受けた人口の割合が高い国での新しい症例が大幅に減少しました。しかし、COVID-19ワクチンの有効性を改善し、副作用を減らすことは、依然として緊急に解決する必要のある問題です。 SARS-CoV-2ウイルスに感染したワクチン接種を受けた人々のごく一部が、重度のCOVID-19疾患を発症しています(Brazal,2021.Frenck et al.,2021)。さらに、ワクチン接種は深刻な副作用(死亡を含む)を引き起こし、多くの人々によるCOVID-19ワクチンの受け入れを減少させています。したがって、SARS-CoV-2感染に対するヒトの免疫応答のメカニズムと、COVID-19ワクチンの有効性および悪影響に関連する要因を十分に理解することが問題を解決するために必要です。 |
| SARS-CoV-2は主に気道から侵入してヒトに感染しますが、胃腸管からも侵入する可能性があります(Chen et al.,2021.Chen et al.,2021.Chen et al.,2021)。ウイルスは、アンジオテンシン変換酵素2(ACE2)受容体に結合することによって細胞に侵入します。ほとんどのヒト細胞はアンジオテンシン変換酵素2を発現するため、ウイルスはアンジオテンシン変換酵素2シグナル伝達による炎症に寄与する多数の臓器に感染し、重度の感染症で多臓器不全を引き起こします(Iwasaki et al.,2021)。感染の結果は、ウイルス量と人間の免疫応答の相互作用に依存します。適切な免疫応答はウイルスを排除し、病気からの完全な回復につながる可能性があります。病理学的免疫応答は、急性呼吸窮迫症候群(ARDS)と多臓器不全を引き起こし、死に至る過炎症とリンパ球減少症を特徴としています。ほとんどのCOVID-19症例は無症候性または軽度であり、ウイルスを排除するのに十分な免疫応答を示しています。 COVID-19ワクチンは、強力でバランスの取れた免疫応答を誘発して、SARS-CoV-2感染を予防するか、まだ感染している場合は病気の重症度を軽減するように設計されています。ほとんどのワクチンは、ウイルスのスパイクタンパク質の受容体結合ドメイン(RBD)に対する抗体を生成して、ウイルスがヒト細胞に侵入するのを防ぎます。 ModernaやPfizerのワクチンなど、一部のワクチンの有効性は95%に達する可能性があり、ほとんどの状況で軽微な副作用がありますが、他のワクチンの有効性は50〜80%に達します。 COVID-19ワクチンは、高齢者や慢性疾患の根底にある人々では効果が低下する可能性があります。 SARS-CoV-2の新しい変異体は、ワクチンの有効性を低下させる可能性があります(Kumar et al.,2021.Kumar et al.,2021.Xie et al.,2021.Zou et al.,2021)。したがって、COVID-19ワクチンの非効率性を引き起こす要因と発生する可能性のある悪影響を研究することが重要です。 |
| このナラティブレビューでは、SARS-CoV-2ウイルス感染に対するヒトの有益な免疫応答とCOVID-19ワクチンに対する免疫応答を要約します。 腸内細菌叢と粘膜免疫の老化、COVID-19のワクチン接種に対する腸内毒素症の影響、およびワクチンの有効性を改善し、悪影響を防止するための腸内細菌叢の調節について説明します。 |
| 2. 免疫および腸内細菌 |
| 腸内微生物叢の組成とその生理学的機能は、宿主の健康、代謝性疾患、老化と密接に関連しています(Turroni et al.,2009)。 腸内細菌叢は、免疫細胞の発達と適切な免疫応答の維持に関連していることがよく知られています(Lynn et al.,2021.Vitetta et al.,2017)。 老化した腸の重要な特徴は、腸のバリアと粘膜免疫系の機能に悪影響を与えるサイトカインIL-6などの炎症誘発性活性の過剰発現です(Man et al.,2015)。 免疫低下は加齢(すなわち免疫老化)の特徴であり、そのため免疫系は重大なリモデリングの影響を受け(Jiang et al.,2013.Weiskopf et al.,2009)、結果として健康と生存に有害な結果をもたらします。 |
| 腸内毒素症は免疫学的不均衡に関連している(BaradaranGhavami et al.,2021)。 免疫学的不均衡は、Tヘルパー2(Th2)の活動亢進およびTreg欠損症として説明されることがよくあります(Baradaran Ghavami et al.,Mezouar et al.,2018)。 腸内細菌叢の関与と免疫関連の結果が最近レビューされ(Zheng et al.,2020)、腸内細菌叢が肺や肝臓などの最終臓器に誘発する特定の影響を表1に示します。 |
 |
| 実験的および臨床的疫学研究(Yu et al.,2016)は、ホメオスタシスを進行させる腸-肺軸を生成する腸と肺の間の極めて重要なクロストークの重要性を強調しています(Dang and Marsland,2019)。 したがって、腸内細菌叢は、肺の健康または病気の機能的メディエーターと見なすことができます。 有害な食事療法と肥満、抗生物質の投与、および疾患プロセス(T2Dなど)による腸内微生物叢の有害な変化(すなわち、腸内毒素症)は、炎症誘発性免疫応答に偏った腸の炎症性緊張を混乱させる可能性があります(表2を参照)。 |
 |
| 最近、肺の微生物叢がレビューされました(Yagi et al.,2021)。 簡単に言えば、独立した分子技術が、多様で少量の微生物が肺および関連する気道に共存することを報告していることをここに追加するだけで十分です。 これは、肺が外部環境に絶えずさらされていることを示しています。 健康な個人の下気道では、Streptococcus, Prevotella、Veillonellaのグループからの細菌の量は少ないものの、多様な存在が報告されています(Dickson et al.,2016,Hilty et al.,2010,Mathieu et al.,2018,Pattaroni et al.,2018,Yu et al.,2016)。 肺の病気の状態では、健康と病気の肺の微生物叢を比較した研究で、有意差が報告されています。 |
| 構成において(Philley et al.,2019,Yagi et al.,2021)、 肺疾患は、1つの分類群または少数の分類群が優勢であるため、細菌の多様性が失われることに関連しています(Tunney et al.,2013)。 |
| さまざまな年齢層の参加者を対象とした糞便微生物叢分析の横断研究では、腸内細菌叢の年齢に関連するプロファイルが、加齢に伴う組成と多様性の変化を示したことが報告されています(Claesson et al.,2011)。コアマイクロバイオータファミリーのBifidobacteriaceae、Bacteroidaceae、Ruminococcaceae、およびLachnospiraceaeは、高齢者グループでは豊富ではなくなります(Biagi et al.,2016,Claesson et al.,2011,Rampelli et al.,2020,Wu et al.,2019)。ワクチンは、感染症を予防するための最も効果的な手段であり続けています(de Jong et al.,2020)。ネズミの研究と因果関係の報告から統合されたもっともらしい提案は、微生物叢がヒトの免疫に及ぼす影響を示しています(de Jong et al.,2020)。それでも、腸内細菌叢全体と免疫系との関係についての議論は指数関数的に増加していますが、ワクチン接種に対する免疫に対する微生物叢の影響はよくわかっていません。 in vitroおよびin vivo実験モデルは、B adolescentisなどの種を含むBifidobacterium属が、Caco-2細胞(訳者注:ヒト結腸癌由来の細胞)およびHT-29細胞(訳者注:ヒト結腸腺癌由来の細胞)へのノロウイルスの結合(Li et al。、2016)とコクサッキーウイルスB3からHeLa細胞(訳者注:ヒト由来の最初の細胞株。 不死化した細胞株として世界各地で培養され、in vitroでの細胞を用いる試験や研究に幅広く用いられている)へ(Kim et al.,2014)の結合を減少させるのに効果的であると報告しています。 Tanと同僚(Tan et al.,2016)は、無菌マウスの腸で、B.adolescentisが腸の炎症に悪影響を与えることなく強力なTh17(訳者注:白血球の一種であるヘルパーT細胞(Th細胞)のサブセットの一つであり、インターロイキン(IL)-17を産生する能力を有する):応答を誘導することを実証しました。 さらに、腸疾患の低慢性炎症動物モデルにおいて、Phillipと同僚(Philippe et al.,2011)は、B.bifidumとB.animalisの菌株が免疫マーカーと腸上皮バリアの完全性を回復したことを報告しました。 B. longum CECT 7347は、グリアジン誘発性腸疾患動物モデルの腸内で、CD4 + T細胞を介した免疫応答を伴う炎症性サイトカイン産生を減弱させました(Laparra et al.,2012)。 10年以上前の研究は、Bifidobacterium属が免疫調節作用を通じて宿主の健康に有益なアウトームを発揮できるという主張を支持してきました。特に最近のレビュー(c.f. Ruiz et al,2017)は、放線菌門内のBifidobacterium属が有意な免疫調節効果を示すという証拠を裏付ける多数の動物およびヒトの研究について包括的に報告しています。さらに、Bifidobacterium属のメンバーは新生児の腸の初期のコロニー形成者であり、そのため、この属は授乳期の乳児で非常に顕著です(Milani et al.,2017)。 |
| 高齢者は感染症にかかりやすくなっています。 初期の臨床調査では、Bifidobacterium属のプロバイオティクス細菌が高齢者の細胞性免疫の特定の側面を強化できることが報告されています(Chiang et al.,2000,Gill et al.,2001)。 小規模な研究ではあるが臨床的証拠を評価した系統的レビュー(Miller et al.,2017)は、Bifidobacterium animalis ssp lactis HN019が健康な高齢者のナチュラルキラー(NK)細胞および多形核細胞の機能を増強すると結論付けました。 |
| さらに、長期生存者の老化は、亜優占種の存在量の増加の特徴を示した細菌種の共起ネットワークの再編成につながる可能性があることが最近報告されました(Rampelli et al.,2020)。 この観察結果は、腸内細菌の濃縮と、ウェルコミクロビウム門(Akkermansia)、Bifidobacteriales、およびChristensenellaceaeなどの細菌の健康に関連する種の存在量の増加と相関していました。 種レベルでは、Bifidobacterium adolescentis、Bifidobacterium longum、Bacteroides Uniformis、Faecalibacterium prausnitzii、Ruminococcus bromii、Subdoligranulum sp。、Anaerostipes hadrus、Blautia obeum、Ruminococcus torques、Coprococcus catus、 Coprococcus、Dorea longicatena、Roseburia sp.、を含む特定の細菌は、腸の健康の改善に関連していると報告されました(Rampelli et al.,2020)。 |
| 3. SARS-CoV-2感染に対する免疫応答 |
| SARS-CoV-2ウイルスに感染したほとんどの患者は、無症候性または軽度の疾患症状を経験します。 これらの患者の免疫反応は、適応免疫反応で効果的でした。 適応免疫の結果は、ウイルスを効果的に排除し、患者が病気から回復することを可能にする適切な応答を伴う免疫系の認識を示していた(Chen et al.,2021,Maecker,2021)。 |
| このような適切な免疫応答の最初の特徴は、免疫応答の経過とSARS-CoV-2感染からの回復を示した事例研究からのものでした(Thevarajan et al.,2020)。軽度から中等度のCOVID-19と診断された女性患者では、SARS-CoV-2感染の初期段階で強い体液性免疫応答と細胞性免疫応答の両方が見られました。ウイルスに対する特異的IgGおよびIgM抗体は、初期段階(症状発現の7日目および9日目)に産生され、抗体分泌細胞の増加(7日目)および循環T濾胞ヘルパー細胞(7日目)を伴っていました(Thevarajan et al.,2020)。細胞傷害性CD8+T細胞は、7日目に増加し、9日目に最高レベルに達し、20日目に減少し、グランザイムA、グランザイムB、およびパーフォリンが増加しました(Thevarajan et al.,2020)。 CD16 + CD14 +単球の血中濃度は、健康なドナーの血中濃度よりも低く、これらの細胞が感染部位にホーミングしていることを示しています。患者と健康な患者の血液サンプルでは、ナチュラルキラー細胞に違いは見られませんでした。わずかに増加した炎症性サイトカインおよびケモカインが見られ、この患者ではサイトカインストームが形成されなかったことを示唆している。 |
| 以下の研究は、SARS-CoV-2感染に対する免疫応答の全体像をさらに示しています。これには、自然免疫応答と適応免疫応答の両方が含まれます(図1)。 COVID-19における自然免疫の重要な役割が示されています。 最も重要なことは、ウイルス複製を阻害し、炎症誘発性応答を調整する一時的なインターフェロンI(IFN I)産生は、軽度のCOVID-19と関連しており、低IFN I産生の遅延は、強力なウイルス複製と重度の疾患を引き起こします(Zhou、Z. et al., 2020)。 持続性/過剰なIFNIおよび過剰な炎症性サイトカインレベルは、複数の組織損傷を引き起こし、重篤な疾患を引き起こす可能性もあります。 |
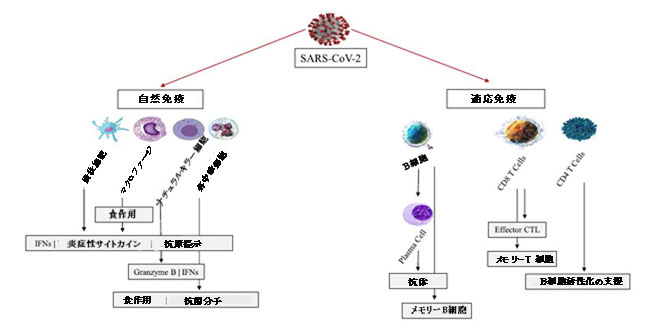 |
| 図1 SARS-CoV-2に対する自然免疫応答 |
| SARS-CoV-2感染は、自然免疫応答と適応免疫応答の両方を誘発します。 樹状細胞(DC)、マクロファージ、ナチュラルキラー細胞、および好中球を含む自然免疫細胞の活性化は、IFN、炎症性サイトカイン、グランザイム、抗菌分子、およびウイルスを含む食作用の分泌をもたらします。 樹状細胞およびマクロファージはまた、適応免疫細胞に抗原を提示して、体液性および細胞性免疫応答を誘発します。 産生された抗体はウイルスを中和することができ、細胞傷害性CD8+T細胞はウイルスに感染した細胞を排除することができます。 メモリーB細胞とメモリーT細胞が産生され、ウイルスのその後の感染に対して迅速な応答を提供します。 |
| 自然免疫細胞はウイルスの侵入を防ぐことができます。樹状細胞およびマクロファージは、適切な量のIFN Iと、炎症性サイトカイン(IL-6、TNF-αおよびIL-1β)およびケモカイン(CCL2およびCCL7)を分泌して、保護効果を発揮します(Zhou et al.,2020、 Zhou et al.,2020)。これらの細胞はまた、適応免疫細胞に抗原を提示します。しかし、サイトカインストームによる過剰反応は病的であり、重度のCOVID-19を引き起こします。 ナチュラルキラー細胞はグランザイムBとパーフォリンを分泌してウイルス感染細胞を排除し、ウイルス複製を減少させることができます(Zuo and Zhao,2021)。好中球は、感染した上皮細胞からの走化性因子によって感染部位に迅速に動員されます。好中球は、食作用によって病因を制限し、抗菌分子を放出し、好中球細胞外トラップを形成する可能性があります(Laforge et al.,2020)。しかし、活性化された好中球は、活性酸素種、プロテアーゼ、および好中球細胞外トラップを生成する可能性があり、過剰な場合、組織の損傷を引き起こす可能性があります(Kang et al.,2021)。適切な自然免疫応答は、SARS-CoV-2感染からの第一線の迅速な保護を提供します。自然免疫応答の失敗は、ウイルス量の増加と過炎症を引き起こし、重篤な疾患につながる可能性があります。たとえば、樹状細胞の低下は、重度のCOVID-19に関連しています。 |
| 体液性および細胞性応答を含む抗原特異的適応免疫応答は、マクロファージおよび樹状細胞を介した抗原提示によって活性化されます。 SARS-CoV-2感染の封じ込めにおける抗体の重要な役割が強調されています(Leslie、2020)。別の研究では、中和抗体レベルが防御能力と関連していることが示されました(Khoury et al.,2021)。中和抗体のレベルがそれぞれ収束平均値の20.2%と3%に達した場合、感染と重度のCOVID-19から宿主を保護するのに十分でした。 Mantus et al(2021)は、入院中のCOVID-19患者の体液性応答を特徴づけ、生成された抗体には、主要なウイルス中和に関連するRBDに対する高レベルのIgGを含む、抗RBD、抗全長スパイク、抗核タンパク質が含まれることを発見しました。 (Mantus et al.,2021)。 Sterlin et al(2021)は、IgAが初期感染で強力に分泌され、IgA抗体がIgGやIgMよりも強力にSARS-CoV-2ウイルスを中和することを発見しました(Sterlin et al.,2021)。 |
| COVID-19におけるT細胞応答のモデルが提案されています。 軽度の疾患では、IL-2、I型およびIII型インターフェロンは、エフェクターT細胞、最終分化T細胞、またはメモリーT細胞となる活性化T細胞のクローン増殖を促進します(Chen and John Wherry,2020)。 一貫して、Liao et al(2020)は、中等度の疾患の症例で高度にクローン増殖したCD8 + T細胞を発見しました(Liao et al.,2020)。 重度の疾患では、IL-6、IL-10、CXCLタンパク質(CXCL8など)およびTNF-αがT細胞プールと活性化T細胞の数を減らし、枯渇または最終分化したT細胞につながる可能性があります(ChenおよびJohn Wherry, 2020)。 これは、PD-L1 / PD-1軸が増加し、T細胞の枯渇を引き起こす可能性があるという最近の発見によっても裏付けられます(ChenとVitetta,2021、ChenとVitetta、2021、Vitte et al.,2020)。 したがって、T細胞応答はSARS-CoV-2感染と戦う上でも重要です。 |
| 回復したCOVID-19患者の回復期の免疫プロファイルは、その後のSARS-CoV-2感染に対するさまざまな保護効果を明らかにしました。風土病のコロナウイルス感染に対する免疫応答は、その後の感染から宿主を保護することはできないが、病気の重症度を軽減することはできることが示されている(Tan et al.,2021)。同様に、無症候性または軽度のSARS-CoV-2感染による回復期の免疫応答は、その後の感染に対して強い免疫を引き出すことができません(Sui et al.,2021)。中等度および重度のCOVID-19感染は、強力な体液性および細胞性免疫応答を誘発する能力をもたらし、その後の感染から宿主を保護する可能性があります(Yan et al.,2020、Zhang et al.,2021)。 Zhang et al(2021)は、重度のCOVID-19と比較して、重度のCOVID-19の回復期血清に高力価の抗S1および抗S2抗体があることを示しました(Zhang、J. et al.,2021)。抗原特異的CD4+T細胞、特にCXCR3 +のサブセットは、抗体価と相関していました。これらの細胞はIL-21とIFN-γも分泌しました。回復期の抗体の持続期間も疑問視されています。 IgGとIgAは、症状が現れてから6〜10週間後に低下し始め、抗体の中和能力は数週間後に低下することが示されています(Beaudoin-Bussièresetal.,2020)。メモリーB細胞とT細胞は、軽度のCOVID-19感染後でも、長期間にわたって保護効果を発揮する可能性があります(Gaebler et al.,2021、Rodda et al.,2021)。これらの細胞は、抗原特異的T細胞およびB細胞の急速な増殖、ならびに二次感染に遭遇したときの抗体の迅速な産生を可能にします(Cox and Brokstad,2020、Sokal et al.,2021、Tavukcuoglu et al.,2021)。 |
| SARS-CoV-2に対する免疫応答がどのように病的になるかは、調査の重要なトピックです。 これは、慢性炎症状態と抗炎症メカニズムの低下に関連している可能性があります(Chen.J. et al.,2021)。 腸内毒素症はそのような状態と密接に関連しており、過炎症の形成を促進します。 重度のCOVID-19では、炎症誘発性マクロファージが豊富にあります(GrauandFélez,1987)。 過炎症はPD-L1を活性化し、CD8 + T細胞の枯渇を引き起こし、ウイルスの排除を遅らせる可能性があります(Chen and Vitetta,2021、Chen and Vitetta,2021)。 同様に、腸内毒素症は、ワクチン接種から誘発される適切な免疫応答を妨げる可能性があります。これについては、後のセクションで詳しく説明します。 |
| 要約すると、自然免疫応答と適応免疫応答は、SARS-CoV-2感染に対する一次および二次防御を提供します。 適切な対応により、侵入するウイルスを排除し、病気からの回復につながる可能性があります。 しかし、ウイルスの急速な複製を伴うI型IFN分泌の遅延や、過炎症を伴う過剰反応免疫などの不十分な反応は、重篤な疾患や死亡につながる可能性があります。 再感染の予防におけるさまざまな回復期の免疫プロファイルは、抗体による短期的な保護と、メモリーB細胞およびメモリーT細胞による長期的な保護によって研究されてきました。 |
| 4. 腸内毒素症によるワクチンとメカニズムの非効率性 |
| さまざまなCOVID-19ワクチンが開発され、SARS-CoV-2感染から防御するための適切な免疫応答を開始します。ただし、SARS-CoV-2に対する免疫応答を妨げる危険因子も、COVID-19ワクチンの有効性を低下させる可能性があります。腸内毒素症は、多くの危険因子によって引き起こされるCOVID-19の重症度と関連しています(Chen、J. et al.,2021)。実際、SARS-CoV-2以外のウイルスに対するワクチン接種に対する免疫応答における腸内細菌叢の役割は、動物実験と臨床研究の両方で十分に実証されています(Lynn et al.,2021、Vitetta et al.,2017)。動物モデルでは、無菌(GF)マウスまたは抗生物質で治療したマウスは、さまざまな面で免疫応答が低下していました。 Kim et al(2016)は、鼻腔内コレラ毒素粘膜ワクチン接種に対する免疫応答が、B細胞数、抗原特異的IgG、想起刺激サイトカイン応答、濾胞ヘルパーT細胞応答の減少を含む無菌マウスまたは抗生物質治療マウスで減少したことを示しました(Kim et al .,2016年、Kim et al .,2019年)。 Ichinohe et al(2011)は、バンコマイシン、ネオマイシン、メトロニダゾール、アンピシリンなどの抗生物質の混合物によるマウスの併用治療が、呼吸器インフルエンザウイルス感染に対する免疫応答を低下させることを明らかにしました(Ichinohe et al.,2011)。 CD4 + T細胞、CD8 + T細胞、およびウイルス特異的抗体価の両方が減少し、肺のウイルス力価は増加しました。 リンパ球性脈絡髄膜炎ウイルス(LCMV)マウスモデルにおいて、Abt et al(2012)は、抗生物質治療マウスがウイルスクリアランスを遅らせ、CD8 + T細胞およびリンパ球性脈絡髄膜炎ウイルス特異的IgG産生を減少させたことを示しました(Abt et al.,2012)。この研究はまた、マクロファージがI型およびII型IFNに対する応答を低下させ、ウイルス複製を制限する能力を損なうことも示しました。これらの動物実験は、ウイルス感染から防御するための効果的な免疫応答には腸内共生細菌が必要であることを示しています。実際、Bifidobacterum longum subsp infantisは、結核、ポリオウイルス、破傷風毒素のワクチンにおける抗原特異的T細胞応答と相関関係があります(Huda et al.,2014)。さらに、タンパク質食の不足によって引き起こされる異生物症は、抗体分泌細胞、CD4 + T細胞、CD8 + T細胞および制御性T (Treg) 細胞の細胞数の減少、ならびにサイトカイン産生の減少など、経口弱毒化ヒトロタウイルスワクチン接種に対する免疫応答の低下ももたらしました(Kimetal.,2020、Michael et al.,2020、Miyazaki et al.,2018)。 |
| ヒトでは、ワクチン有効性における腸内細菌叢の役割は、さまざまなアプローチによって証明されています。同じワクチンが低中所得国(LMIC)と高所得国(HIC)で異なる反応を誘発し、低中所得国の有効性の低さは、集団の腸内毒素症によって説明されました(Lynn et al.,2021)。短期間の母乳育児、栄養失調、下痢のある乳児の腸内毒素症は、経口ポリオワクチンの有効性の低下と関連がありました(Haque et al.,2014)。腸内毒素症は、ワクチン有効性に対するワクチン接種前の免疫状態の影響とも関連していた。ワクチン接種に対する免疫応答は、ワクチン接種前の免疫状態と関連しています。たとえば、インフルエンザワクチンの有効性は、65歳以上の人口ではわずか30〜50%でしたが、若い成人では70〜90%でした(Osterholm et al.,2012)。高齢の呼吸器合胞体ウイルス(RSV)ワクチンの非応答者では、より高い(HLA-DR +)CD4+およびCD8+ T細胞と、CCR7、CD127、およびCD69の発現の増加を伴うベースライン免疫プロファイルがあり、慢性炎症状態を示しています(Lingblometal.,2018)。 Fourati et al(2016)は、ワクチン接種前の炎症がB型肝炎(HepB)ワクチン接種に対する反応低下と関連していることを発見しました(Fourati et al.,2016)。これらは、ワクチン接種に対する免疫応答の低下を引き起こす腸内毒素症によって引き起こされる慢性炎症によって説明される可能性があります。 |
| ワクチン有効性における腸内細菌叢の役割のメカニズムは、フラジェリンやペプチドグリカンなどの腸内細菌分子によって活性化される抗原提示細胞のパターン認識受容体(PRR)に起因しています(Lynn et al.,2021)。 パターン認識受容体は自然免疫細胞に豊富に存在し、腸内細菌分子によって活性化されると急速に刺激に反応します(つまり、免疫トレーニングと呼ばれます)(Smith et al.,1988、Yakabe et al.,2021)。ペプチドグリカンによるパターン認識受容体、ヌクレオチド結合オリゴマー化ドメイン含有タンパク質2(Nod2)の結合は、cAMP産生を増加させ、樹状細胞がサイトカインを分泌するのを促進します(Kim et al.,2016、Kim et al.,2019)。合成アゴニストによるヌクレオチド結合オリゴマー化ドメイン含有タンパク質2の活性化は、無菌マウスまたは抗生物質治療マウスにおけるコレラ毒素誘発免疫応答を再構成し、腸内細菌叢刺激免疫応答におけるヌクレオチド結合オリゴマー化ドメイン含有タンパク質2の重要な役割を示しています(Kim et al.,2016、Kim et al.,2019 )。共生/共生細菌によるヌクレオチド結合オリゴマー化ドメイン含有タンパク質2刺激は、最適なCT(訳者注:CT(Computed Tomography:コンピュータ断層撮影)とは、X線を利用して身体の内部(断面)を画像化する検査です)を介した抗原特異的経口ワクチン接種効果に寄与し、これはIL-1βの誘導とその後のレベルの上昇によって瞑想されたことが報告されました。 Toll様受容体5(TLR5)は、形質細胞の発達と抗体産生のためにフラッグリンによる活性化を必要とします(Oh et al.,2014、Pabst and Hornef,2014)。 TGF-βはクラスターIVおよびXIVaclostridiaによって刺激され、平衡免疫応答を維持するために重要な制御性T細胞(Treg細胞)を活性化することができます(Atarashi et al.,2011)。対照的に、腸内毒素症は、酪酸などの共生細菌代謝物の変化を介して慢性炎症を引き起こし、抗炎症メカニズムを低下させます(Chen.J. et al.,2021)。酪酸は、制御性T細胞を活性化して抗炎症効果を発揮することでよく知られています(Chen and Vitetta,2018、Furusawa et al.,2013)。マウスモデルでは、酪酸はGPR41とGPR109aの活性化、およびヒストンデアセチラーゼの阻害を通じて結腸でのIgA産生を促進しました(Isobe et al.,2020)。酪酸は、腸のバリアの完全性を維持する上でも重要です。酪酸産生の減少は、エンドトキシンと細菌の循環系と腸外器官への移行をもたらし、直接的な組織損傷を引き起こすSARS-CoV-2によって引き起こされる過炎症の形成を促進します(Chen、J. et al.,2021)。過敏反応や炎症によって免疫応答を混乱させる可能性のある腸内毒素症は、COVID-19ワクチン接種によって誘発される免疫応答を妨害し、COVID-19ワクチンの有効性に影響を与える可能性があります(Ferreira et al.,2020a、Ferreira et al.,2020b)。 |
| 5. 危険因子をCOVID-19ワクチンの非効率性に関連付ける腸内毒素症 |
| 腸内細菌叢がさまざまな微生物に対するワクチンの有効性に及ぼす影響と、COVID-19の重症度における腸内毒素症の役割に基づいて、腸内細菌叢がCOVID-19ワクチン接種の有効性にとって重要である可能性があることは驚くべきことではありません( Ferreira et al.,2020a、Stefan et al.,2021) そして、その腸内毒素症は、COVID-19ワクチンの有効性を低下させる可能性があります(Lynn et al.,2021)。重度のCOVID-19と高い死亡率を示す脆弱な集団は、優先的にワクチン接種されるべきであると提案されています(Leong et al.,2021、Russo et al.,2021、Wingert et al.,2021)。脆弱な集団は、高齢、基礎となる慢性疾患(肥満、糖尿病、慢性心血管、肺、肝臓、腎臓の疾患など)および精神障害(ストレス、不安、うつ病など)を特徴としています(Chen et al.,2021、Koff et al.,2021、Stefan et al.,2021)。腸内毒素症は、これらの危険因子が病理学的免疫応答を引き起こすメカニズムの1つです。 SARS-CoV-2感染に対する防御力を低下させる危険因子は、COVID-19ワクチンに対する反応を低下させ、副作用を増大させる可能性もあります。最近の研究では、加齢、肥満、高血圧がファイザー/バイオエヌテック(Pfizer/BioNTech)ワクチンの有効性を著しく低下させることが示されました(Koff et al.,2021、Pellini et al.,2021、Stefan et al.,2021)。 |
| 高齢者、特に基礎疾患のある人々における有効性と副作用は、COVID-19ワクチン接種にとって重大な問題でした(Connors et al.,2021、Soiza et al.,2021)。確かに、老化は重度のCOVID-19感染の最も重要な危険因子であると考えられてきました(Wingert et al.,2021)。 老化は慢性炎症と免疫応答の低下に関連しています。これらの変化は、免疫老化と炎症の両方を引き起こす腸内細菌叢の老化によって引き起こされる可能性があります(Bosco and Noti,2021、Connors et al.,2021)。 研究によると、高齢化した人口は予防接種にあまり反応しないことが示されています(Bosco and Noti,2021)。メタアナリシスは、ワクチンは入院率と死亡率を低下させる可能性があるものの、ワクチンは高齢者に最適な抗体反応を引き起こすことができなかったことを示しました(Almasri and Holtzclaw,2021)。 重要なことに、フレイルインデックスによって示される生物学的老化は、時系列の老化よりも関連する腸内毒素症の変化とより相関しています(Kim and Jazwinski,2018、Maffei et al.,2017)。 Muller et al(2021)は、60歳未満と80歳を超える患者の2つのグループでファイザーワクチンに対する免疫応答を比較しました(Mülleretal.,2021)。後者は抗体価が有意に低かった。 2回目の投与後でも、31.3%の高齢者グループでは中和抗体は検出されず、若いグループでは2.2%しか検出されませんでした(Mülleretal.,2021)。したがって、早期の再ワクチン接種またはワクチン投与量の増加を促進することが提案された。しかし、最近の臨床試験では、アストラゼネカは高齢者の方が忍容性が高く、同様の免疫原性の結果が得られたことが報告されています(Ramasamy et al.,2021)。体系的なレビューでは、COVID-19ワクチンのほとんどの臨床試験では通常、高齢者が除外されており、ワクチンの有効性を分析するにはデータが不十分であることが示されました(Hou et al.,2021)。高齢者グループにおけるCOVID-19の非効率性における腸内毒素症の関与は、広範な研究を保証します。腸内細菌叢の調節は、COVID-19の有効性を高めるためのより良いアプローチを含む可能性があります。 |
| タンパク質の欠乏などの栄養失調は、抗体分泌細胞を減少させ、動物モデルのロタウイルス感染におけるIgAおよびIgG産生の減少につながることが実証されています(Michael et al.,2020、Miyazaki et al.,2018)。 これは、栄養状態がCOVID-19ワクチンの有効性において重要である可能性があることを示唆している可能性があります。 実際、ビタミンC、ビタミンD、ラクトフェリン、オメガ3脂肪酸などの栄養補助食品、および亜鉛、セレン、マグネシウムなどの微量元素は、腸内細菌叢を改善し、COVID-19の重症度を軽減することができます(Chen & Vitetta,2021、Chen & Vitetta, 2021)。 これらの成分の欠乏は深刻なCOVID-19を引き起こします。 |
| 肥満は、COVID-19ワクチン接種の有効性低下の主な懸念事項でもあります(Kipshidze et al.,2021、Ledford,2020、Townsend et al.,2021)。低体重および正常体重の患者は、過体重および肥満の患者よりも抗体価においてはるかに高い有効性を有することが示されているが、他の免疫応答をさらに調査することができる(Pellini R et al.,2021)。慢性炎症を特徴とする肥満は、重度のCOVID-19の危険因子であり、腸内毒素症は、効果のない免疫応答をもたらすメカニズムの1つです(Chen.J. et al.,、2021)。腸内毒素症がCOVID-19ワクチン接種の非効率性にどのように関与しているかは、さらに専用の研究が必要です。腎臓または肝臓移植患者の体液性免疫応答は、通常は臨床試験で除外されているが、減少したことも報告されている(Marjot et al.,2021、Mossad,2021、Sui et al.,2021)。最近の研究では、肝移植患者の61%と他の慢性肝疾患の24%が、COVID-19ワクチン接種に対する抗体反応が不良であることが報告されています(Thuluvath et al.,2021)。肝移植におけるCOVID-19ワクチンの低い免疫原性は、年齢、腎機能、および免疫抑制薬の要因と関連していた(Rabinowich et al.,2021)。対照的に、ある研究では、非アルコール性脂肪肝疾患(NAFLD)がCOVID-19ワクチンの有効性に影響を与えなかったことが示されました(Wang et al.,2021、Wang et al.,2021、Wang et al.,2021)。ワクチンの有効性に関して軽度と重度の肝疾患を区別する必要があるかもしれません(Cornberg and Eberhardt,2021)。ある研究によると、慢性腎臓病は、尿毒症環境、ビタミンD欠乏症、およびエリスロポイチンが原因の抗原提示細胞の機能不全とB細胞数の減少により、ワクチン有効性の低下と関連していることが報告されています(Hou et al.,2021)。尿毒症環境とビタミンD欠乏症の両方が腸内毒素症を引き起こすことが知られています(Chaves et al.,2018、Chen et al.,2021)。 |
| COVID-19の重症度の性差は十分に実証されており、その結果は誘発された免疫応答の違いによるものです(Alwani et al.,2021、Mateus et al.,2021)。 これらの免疫応答は、COVID-19ワクチンの有効性の違いを裏付ける可能性もあります。 性ステロイドは腸内細菌叢を調節することが知られており(Manosso et al.,2021、So and Savidge,2021)、したがって腸内細菌叢を介してワクチンの有効性を仲介する可能性があります。 そのような研究は、COVID-19ワクチンの有効性の性差における腸内細菌叢の役割を調査することが保証されています。 |
| 6. COVID-19ワクチンの有効性 |
| 現在、弱毒生ウイルスワクチン、不活化ウイルスワクチン、タンパク質サブユニットワクチン、複製欠損ベクター、遺伝子ワクチン(DNAおよびRNAワクチン)など、多数のCOVID-19ワクチンが開発されています(Min and Sun,2021)。 現在、世界中で少なくとも10を超えるCOVID-19ワクチンが緊急使用のために承認されています。 これらのワクチンは、臨床試験からさまざまなレベルの有効性でSARS-CoV-2に対する免疫応答を誘発することが示されています。 3つの米国ワクチン(ファイザー/バイオエヌテック(Pfizer / BioNTech)、モデルナ(Moderna)、ジョンソン アンど ジョンソン(Johnson and Johnson))と1つの英国ワクチン(アストロゼネカ(Astrazenica))が西側諸国で広く使用されています。 |
| 6.1 mRNAワクチン |
| mRNAワクチンは最も成功したワクチンです。 mRNAはウイルスタンパク質をコードするように設計されており、ウイルスタンパク質は抗原タンパク質に翻訳されて免疫応答を誘発します。 ファイザー/バイオエヌテックmRNAワクチンはSタンパク質のRBD(訳者注:受容体結合領域は新型コロナウイルスのスパイクタンパク質が細胞の受容体であるアンジオテンシン変換酵素2と結合する領域)をコードし、モデルナワクチンはS-2P抗原をコードします(Noor,2021)。ナノテクノロジーは、人体の急速な分解を防ぐためにmRNAの送達に適用されてきました。脂質ナノ粒子(LNP)は、両方のmRNAワクチンを処方するために使用されます。免疫応答を生み出すmRNAワクチンの経路を図2に示します(Kowalczyk et al.,2016)。ワクチン粒子が樹状細胞に飲み込まれた後(すなわち、食作用)、mRNAはサイトゾルに放出され、そこでmRNAは抗原タンパク質に翻訳されます。抗原は小胞体のMHCIに結合し、細胞表面に提示され、B細胞を活性化して抗原特異的抗体を産生し、CD8 + T細胞を活性化してリンパ節を排出する抗原特異的細胞傷害性T細胞にします(Kowalczyk et al., 2016)。循環中に放出されたmRNAは、Toll受容体 3,7,8およびRIG-I(レチノイン酸誘導性遺伝子)による認識を通じて自然免疫系を活性化し、INF Iの産生を増加させ、Th1細胞を刺激します(CagigiおよびLoré、 2021)。処方中のLNPは、Toll受容体経路を介して免疫応答を誘発することもあります(Chung et al.,2020)。両方のmRNAワクチンは非常に高い有効率に達し、臨床試験ではモデルナが94.1%、ファイザー/バイオエヌテックが95.0%です(Baden et al.,2021、Polack et al.,2020)。これらの臨床試験は、ワクチンに対する反応率が低い可能性が高い免疫抑制された個人を除外しました。たとえば、抗B細胞CD20抗体であるオクレリズマブとリンパ球を隔離する免疫調節剤であるフィンゴリモドを使用すると、ファイザー/バイオエヌテックの体液性免疫応答の有効率がそれぞれ22.7%と3.8%に低下しました(Achiron et al.,2021)。 |
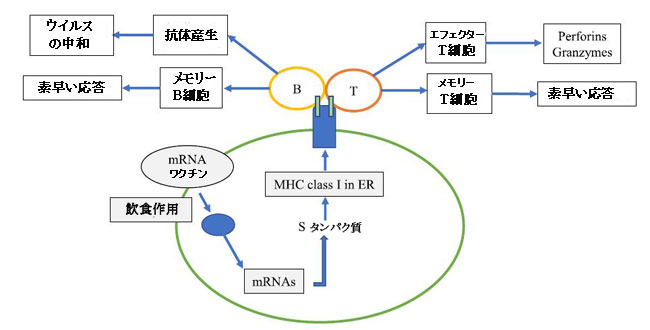 |
| 図2 COVID-19mRNAワクチンに対する免疫応答 |
| mRNAワクチンはエンドサイトーシスを介して抗原提示細胞に入ります。mRNAは細胞質に放出され、Sタンパク質に翻訳されます。 Sタンパク質はERのMHCクラスIに結合し、MHCクラスIによって細胞表面に提示されます。 次に、抗原はB細胞を活性化して抗原特異的抗体を産生し、メモリーB細胞とT細胞はエフェクターT細胞とメモリーT細胞を産生します。 |
| 6.2 アデノウイルスベースのワクチン |
| アデノウイルスベースのワクチンは、強力なアデノウイルスプロモーターを使用して、アデノウイルスバックボーンの導入遺伝子からSARS-CoV-2抗原を発現します。 アストラゼネカ-オックスフォードワクチン(AZD1222)は、オックスフォード大学が開発したチンパンジーアデノウイルス(chAdOx1)を採用しており、ヒト細胞に侵入しますが、複製はしません(Derikvand et al.,2020)。 アストラゼネカ-オックスフォードワクチン(導入遺伝子は、SARS-CoV-2のスパイクタンパク質です。次の第II/III相試験では、70.4%の有効性で以前の結果が確認されました(Ramasamy et al.,2020、Voysey et al.,2021)。 ジャンセンファーマセチカルワクチン(Johnson and Johnson)は、Ad26.COV2.Sを作成しました。これは、1回の免疫化のみが必要です。ベクターは複製能のないアデノウイルス26であり、導入遺伝子は完全長のSARS-CoV-2スパイクタンパク質です(Bos et al.,2020)。単回免疫は、SARS-CoV-2感染から保護するのに十分な抗原特異的抗体で強力な免疫応答を誘発し、全体的な有効性は66.3%であることが示されています(Mercado et al.,2020、Sadoff et al.,2021)。ロシアで開発されたスプートニクVは、COVID-19のスパイクタンパク質をコードするcDNAを運ぶために、2つのアデノウイルスタイプ26と5から作られています(Logunov et al.,2020)。フェーズI/IIの臨床では、安全性と忍容性が示されました。ワクチンは、100%の抗体および細胞性免疫応答と91.6%の有効性をもたらしました(Logunov et al.,2021、Logunov et al.,2020)。スプートニクは現在、世界中の多くの国で使用されています。 CanSino Biologicals(Wu et al。、2020)のAd5-nCoV(Convidecia)は、中国、ハンガリー、チリ、メキシコ、パキスタンなどの一部の国で緊急使用が承認されています。しかし、まだWHOによるものではありません。臨床試験(フェーズI / II)は、それが安全で効果的であることを示しました(Zhu et al.,2020、Zhu et al.,2020)。 |
| 動物モデルは、アデノウイルスベースのワクチンの免疫原性をテストするために使用されています。 最近の成体アカゲザルの実験では、1x1011 Ad26.COV2.Sウイルス粒子(vp)と5x1010 vpの両方が、2週目に強力な免疫応答を生成し、4週目にさらに増加しました。 高用量のウイルス粒子免疫化は、低用量のそれよりも1.6倍高い結合と2.1倍高い中和抗体を持っていた(Solforosi et al.,2021)。 どちらの場合も、結合抗体は中和抗体よりも急速に低下しました。 4週間または8週間後に2回目の免疫を行うと、結合抗体がそれぞれ5.7倍および11.8倍増加し、中和抗体がそれぞれ7.6倍および15.2倍増加しました。 2回目の投与の2週間後に測定された(Solforosi et al.,2021)。 免疫応答はまた、保護能力が弱まるかもしれないが、ほとんどの老化したアカゲザルを保護するのに十分でした。 |
| 6.3 不活化または弱毒化ウイルスワクチン |
| 不活化または弱毒化ウイルスワクチンは、感染力の破壊と免疫原性の保持の両方を必要とする従来のアプローチです(Xia et al.,2020、Xia et al.,2021)。 Sinovac Biotechは、ホルムアルデヒド不活化全ウイルスSARS-CoV-2ワクチン– CoronaVacを開発しました。これは、WHOによって緊急使用が承認されました(Gao et al.,2020、Mallapaty,2021)。ワクチン処方には、アジュバントとしてミョウバンが含まれています。活性化される免疫応答には、単球分泌IFN、CD4 + T細胞、B細胞、抗体、およびCD8+T細胞が含まれます。臨床試験(Zhao Y,2020)は、ワクチンが安全であり、高齢の参加者に許容されることを示しました(Wu et al.,2021)。 Sinopharmは、BBIBP-CorV(WHOによって承認された)と呼ばれる不活化ワクチンも製造しました。これは臨床試験中です(Garcia,2020、Yang,2020)。第III相臨床試験(Yang Y,2020)から報告された結果は、SARS-CoV-2 WIV04およびHB02からの不活化ワクチンがそれぞれ72.8%および78.1%の有効性に達したことを示しました(Al Kaabi et al.,2021b)。 Bharat Biotechによるコバキシンの自家製および製造ワクチンは広く入手可能であり、何百万人もの医療従事者に投与されます(Kumar,V. M. et al.,2021)。第I相臨床試験では、有効性、安全性、忍容性が示されました(Thiagarajan、2021)。 25,800人の参加者を対象とした進行中の臨床試験では、81%の有効性が予測されました(Thiagarajan,2021)。したがって、不活化ワクチンは、mRNAワクチンよりも有効性は低いものの、十分な防御効果を生み出す可能性があります。 |
| 6.4 COVID-19から回復した患者のワクチン接種 |
| COVID-19ワクチンは、SARS-CoV-2に感染した集団の免疫応答をさらに高めることが示されています。 Mazzoni et al(2021)は、ファイザー/ ビオエヌテックワクチン接種の1回投与は、COVID-19回復患者の体腔および細胞免疫を増加させるのに十分であり、2回目のワクチン接種は免疫応答をさらに増加させないことを明らかにしました(Mazzoni et al.,2021)。対照的に、未経験のワクチン接種を受けた患者が高い免疫応答に達するには、2回目のワクチン接種が必要です。 Levi et al(2021)は、ファイザー/ ビオエヌテックワクチン接種の初回投与後、ナイーブ患者と比較して、以前に曝露されたCOVID-19患者で5〜8日後にIgGレベルが指数関数的に増加したことを示しました(Levi et al.,2021 )。しかし、無症候性でナイーブな患者は、最適な免疫応答に到達するためにワクチンの2回目の投与が必要でした。 Sasikala et al(2021)は、以前にSARS-CoV-2に感染した280人の医療従事者において、mRNAワクチンの単回投与により中和抗体が劇的に増加した(1124.73±869.13vs94.23±140.06AU/ ml)ことを示しました(Sasikala et al.,2021 )。したがって、ワクチン接種は、SARS-CoV-2に曝露されたが、十分な免疫応答が誘発されなかった集団の免疫プロファイルをさらに増加させることができます。 |
| 6.5 ワクチン有効性に対する変異体の影響 |
| COVID-19のパンデミックの拡大と人口の感染の増加に伴い、多くのSARS-CoV-2変異体が出現しました。さまざまな変異体が、ワクチン接種によって引き起こされる免疫応答に対する耐性を、これとは異なる程度で生み出すことが実証されています。耐性の範囲に応じて、WHOはバリアントを懸念のあるバリアントと関心のあるバリアントに区別します。現在、B1.1.7、B.1.351、B.1.1.28のP1およびB.1.167を含む4種類の懸念変異株があります(Jia and Gong,2021)。自然感染およびワクチン接種から単離された抗体は、変異体に対して効果が低いことも示されている(Chen et al.,2021、Hoffmann et al.,2021、Jia and Gong,2021、Khoury et al.,2021、Röltgenetal.,2021、Wang et al.,2021、Wang et al.,2021、Wang et al.,2021)。メタアナリシスは、自然感染またはワクチン接種によって誘発された中和抗体が、B.1.1.7に対して1.5倍、B1.351に対して8.7倍、P1に対して5倍減少したことを示しました(Chen et al.,2021、Chen et al.,2021)。新たに発見された変異体B.1.167.1は、野生型よりもこれらの抗体に対して6.8倍耐性がありました(Edara et al.,2021)。 2つのmRNAワクチンは、野生型スパイクおよびヌクレオカプシドタンパク質(回復期血漿よりもさらに)に対する強力なT細胞応答を誘発しましたが、バリアント(B1.1.7、B.1.351、B.1.1.28のP1およびB.1.167)への影響は大幅に減少しました(Gallagher et al.,2021)。 CDCの報告によると、64%のCOVID-19ブレイクスルー症例は変異型でした(「CDCに報告されたCOVID-19ワクチンブレイクスルー感染症-米国、2021年1月1日から4月30日」2021年)。 |
| 7.COVID-19ワクチンの副作用 |
| すべてのCOVID-19ワクチンの副作用は、緊急使用中に報告されています。臨床試験では、ワクチン接種のための患者の選択基準のために、副作用が過小評価される可能性があります(Klimek et al.,2021)。局所的な痛み、発赤、腫れ、発熱、倦怠感、頭痛、筋肉や関節の痛みなどの全身症状など、深刻ではない副作用が頻繁に報告されています(Polack et al.,2020)。 2つの不活化COVID-19ワクチンの最近の臨床試験では、参加者の41.7%から46.5%で非重篤な副作用が発生し、そのほとんどが局所的な痛みを報告していることが示されました(Al Kaabi et al.,021a、Zhao,2020)。深刻な副作用はまれですが、死亡者を出し、予防接種の受け入れに大きな影響を与えています。 COVID-19ワクチン接種後にノルウェーで報告された虚弱な高齢患者の23人の死亡がありました(Torjesen,2021年)。深刻な悪影響をどのように回避するかは、緊急の解決を必要とする重要な問題です。 |
| アレルギー反応は、2つのmRNAワクチンによって引き起こされています(Longo et al.,2021、Klimek et al.,2021、Selvaraj et al.,2021)。 アレルギーはIgEの過剰産生によって引き起こされることが知られています。 IgEは、肥満細胞および好塩基球上の高親和性IgE受容体(FcイプシロンRI)に結合し、ヒスタミン、プロスタグランジン、ロイコトリエン、プロテアーゼ、および炎症性サイトカインの放出を引き起こします。 ワクチン接種参加者向けのガイダンスは、アナフィラキシーを回避するために作成されました(Banerji et al.,2021、Klimek et al.,2021)。 コンピューター化されたモデルは、可能性のあるアレルゲンが、mRNAワクチンによって発現されるRBD(スパイクタンパク質サブユニット)の437-508のアミノ酸残基である可能性があることを示しました(Selvaraj et al.,2021)。 使用されるナノデリバリーシステムのコンポーネントは、PEG、TRIS、糖リン脂質などのアレルギーを引き起こす可能性もあります(Hatziantoniou et al.,2021、Selvaraj et al.,2021、Troelnikov et al.,2021)。 |
| COVID-19ワクチン接種で報告されたもう1つの重篤な副作用は血栓症であり、これも生命を脅かします。 ジャンセンス、アストラゼネカ(インドのCovishield)、スプートニク V(Gamaleya Research Institute、モスクワ、ロシア)などのアデノウイルスベースのワクチンは血栓症に関連しています(Gupta et al.,2021、See et al.,2021)。 Ad5-nCOVのレシピエントと臨床試験では血栓症は報告されていませんが、後者は5×109 vpの用量で1%、1×1011の用量で9%の高熱や重度などの重篤な副作用があります。疲労(Zhu et al.,2020、Zhu et al.,2020)。血小板の活性化が血栓症の原因であると考えられました。ほとんどの血栓症はアデノウイルスベースのワクチンで起こったので、ベクターとコードされた全長スパイクタンパク質が原因物質であると考えられてきました。現在、mRNA誘発性血栓症の1例のみが報告されています(Carli et al.,2021)。ある研究では、血小板第4因子に対する抗体が、アデノウイルスベースのワクチン接種とmRNAワクチン接種の両方で発見されたことが示されました(Thiele et al.,2021)。血小板第4因子と反応して活性化するIgG抗体は、ワクチン接種の参加率が高いことが検出されています(19/281)。しかし、抗体の力価は通常非常に低く、血小板を活性化するのに十分ではありません。 |
| まれに、抗PD-1免疫療法を受けている結腸直腸癌患者にBTN162b2をワクチン接種した後、サイトカインストームが報告されています(Au et al.,2021)。 血小板減少症を伴う炎症性サイトカインCRP、IFN-γ、IL-2、IL-18、IL-16、および抗炎症性サイトカインIL-10が非常に増加しました。 サイトカインストームは、細胞性免疫応答の感受性の増加とともにワクチンの刺激によって説明することができます。 抗PD-1はCD8+T細胞の活性化を増加させる可能性があります。 |
| 8. COVID-19ワクチン接種のための腸内細菌叢を改善するためのアプローチ |
| COVID-19の腸内毒素症は、COVID-19ワクチンの有効性と悪影響に影響を及ぼしているため、腸内細菌叢の改善により、COVID-19ワクチンの有効性が高まり、副作用が軽減される可能性があります(Lynn et al.,2021) 確かに、COVID-19ワクチンで腸内細菌叢の調節をテストしているいくつかの進行中の臨床試験があります。 検出されたCOVID-19に対するワクチンの有効性を高めるためにABBC1と呼ばれる酵母ベースのプロバイオティクスを体液性応答と細胞性応答の両方による臨床試験実施している(MateusRodríguezJA,2020)。 ABBC1には、β1,3/β1,6-グルカンと不活化Saccharomyces cerevisaeのほか、微量元素のセレンと亜鉛が含まれています。 別の臨床試験でも、3つのビフィズス菌を含む処方を使用して、COVID-19ワクチンの有効性を高め、高齢の2型糖尿病患者の悪影響を軽減しています(Mak JWY,2020)。 3番目の臨床試験は、COVID-19ワクチンの有効性を高めるために5-ALA-リン酸を含む機能性食品を調査しています(Darwish A,2020)。 5-ALA-リン酸は腸内細菌叢の恒常性を維持することが知られています(Chang et al.,2021)。 |
| 乳酸菌とビフィズス菌は、COVID-19ワクチン接種に対する免疫応答を強化する可能性があります。これらの細菌からのMAMPは、Toll様受容体を刺激して、自然免疫応答と適応免疫応答の両方を促進することができます(Moradi-kalbolandi et al.,2021、Vlasova et al.,2013)(図3)。また、ワクチン接種および感染した参加者(すなわち、NCT04980560)のマイクロバイオータプロファイルを調べるための臨床試験が進行中です(図3)。プロバイオティクス、プレバイオティクス、シンバイオティクス、栄養補助食品、微量元素、糞便微生物移植(FMT)、FMD(空腹時模倣食)などの食物/エネルギー制限の投与を含む、腸内細菌叢の多様性を改善するためのさまざまなアプローチが利用可能です。これらのアプローチはすべて、さまざまな条件に適用され、さまざまなレベルの成功を収めています。糞便微生物移植は、抗生物質による腸内毒素症に起因するC.difficle感染症の治療に使用されています。糞便微生物移植は、C.difficle破壊的感染から腸内細菌叢を回復するための好ましいアプローチです。しかし、それは病原性感染症などの深刻な悪影響を与える可能性があります。したがって、COVID-19ワクチンのアジュバントアプローチには適していない可能性があります。プロバイオティクス、プレバイオティクス、シンバイオティクスは、炎症性腸疾患などの多くの慢性感染症の治療に使用されてきました(Chen et al.,2021、Walton et al.,2021)。それらは報告された最小限の副作用で非常に安全です。最も一般的に使用されるプロバイオティクスは、乳酸菌やビフィズス菌などの狭い範囲の生物からのものです。一般的なプレバイオティクスには、フルクトースオリゴ糖、ガラクトオリゴ糖、ベータグルカン、難消化性デンプンが含まれます。一部のプレバイオティクスは、直接的な抗ウイルス効果を持つ可能性があります。たとえば、エピガロカテキンガレートは腸内細菌叢を改善するだけでなく、Nsp15(訳者注:新型コロナウイルスの酵素)を阻害することで強力な抗ウイルス効果を発揮します(Hong et al.,2021)。プロバイオティクスとプレバイオティクスの両方がインフルエンザワクチンの有効性を著しく高めることが報告されています(Lei et al.,2017)。示されているように、これらはCOVID-19ワクチンの有効性を改善する可能性があります(Akatsu、2021)。シンバイオティクスは、共生細菌だけでなく、腸内の酪酸産生レベルを高めることができる食物繊維も提供するため、プロバイオティクスまたはプレバイオティクス単独よりも優れている可能性があります(Holscher.2017年)。酪酸はまた、抗原活性化CD8 + T細胞の長寿命メモリー細胞への移行を増加させます(Bachem et al.,2019、Ji and Hu、2019)。さらに、インドール産生細菌と胆汁酸調節細菌も、COVID-19ワクチン接種での使用の可能性について研究することができます(図3)。これまでに85種の細菌が、トリプトファンをインドールに変換するトリプトファナーゼを産生することが確認されています(Lee and Lee,2010)。インドールは、アリール炭化水素受容体の活性化を通じて抗炎症効果があります(Rothhammer et al.,2016)。共生細菌は、3-オキソリトコール酸、イソアロリトコール酸、isoDCAなどの二次胆汁酸のいくつかの誘導体を産生する可能性があり、制御性T細胞の活性化を通じて抗炎症効果があります(Campbell et al.,2020、Hang et al.,2019)。 |
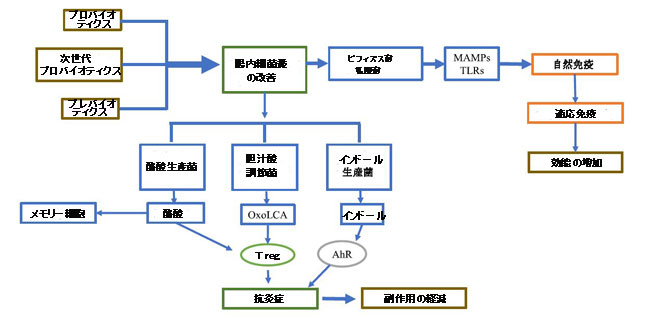 |
| 図3 COVID-19のためのプロバイオティクスとプレバイオティクス |
| プロバイオティクス、次世代プロバイオティクス、およびプレバイオティクスは、腸内細菌叢の存在量を改善し、共生細菌代謝産物を増加させることができます。これにより、生来の免疫応答と適応免疫応答の両方が強化され、COVID-19ワクチンの有効性が高まります。 一部の共生細菌代謝産物は、制御性T細胞を活性化して免疫応答のバランスを取ることもできます。 腸内細菌叢の改善は、COVID-19ワクチンの悪影響を減らすこともできます。 |
| 新世代のプロバイオティクスは細菌の範囲が拡大しており、将来的にはCOVID-19ワクチンを補完するための処方が含まれる可能性があります。 酪酸を直接生成できるF.prausnitziiなどの酪酸生成細菌は、実験的なプロバイオティクス製剤に含まれています(Gautier et al.,2021、O'Toole et al.,2017)、 これは、例えばCOVID-19ワクチンの有効性と安全性を改善する(Moradi-Kalbolandi et al.,2021)など、多くの有益な効果をもたらします(Leylabadlo et al.,2020)。 介入的臨床試験は、F.prausnitziiが許容され、代謝経路を改善したことを示しました(Lorenzon M,2020) Bacteroides acidifaciens、Bacteroides ovatus、Bacillus pumilus、Bacillusmegateriumなどの他の次世代プロバイオティクスはあまり研究されていません。 B. acidifaciensはノトバイオートマウスのIgAを増加させ、B.ovatusはIgMとIgGを増加させました。 これは、C.difficle感染症の治療のための9つのプロバイオティクスの混合物に含まれています(Graham DY.2020)。 この研究は、1人の患者に直腸点滴同種糞便と他の5人の患者に滅菌生理食塩水で希釈した10種類の通性嫌気性細菌と嫌気性細菌の混合物を介して投与した以前のC.difficleによって引き起こされる慢性再発性下痢の治療のため研究(Tvede and Rask-Madsen,1989)の更新です。 最近の研究では、B.pumilusとB. megateriumがマウスモデルでF.prausnitziiを増加させることがわかっており、複雑な細菌の相互作用を示しています(Kotowicz et al.,2019)。次世代のプロバイオティクスがCOVID-19ワクチンに対してより有益な効果を生み出すかどうかは、さらなる研究が必要です。 |
| 腸内細菌叢の調節はまた、COVID-19ワクチンの副作用を減らすことができます。プロバイオティクスはアレルギーを効果的に改善できることが示されています(Jing et al.,2020)。 B. longum、B.breve、L.plantarum、L.casei、L.fermentum、L.rhamnosusを含むいくつかのプロバイオティクスは、動物実験や臨床試験を通じて抗アレルギー効果があることが実証されています(Shu et al.,2019)。 メカニズムは十分に解明されていません(Shu et al.,2019)。 酪酸は重要なメディエーターになる可能性があります。腸内細菌叢における酪酸産生細菌の減少は、食物アレルギーに関連していることが示されています(Lee et al.,2020)。酪酸は制御性T細胞を活性化することが知られており、抗体のIgE産生を低下させる可能性があります。炎症誘発性シグナル伝達経路の阻害は、酪酸が抗アレルギー効果を発揮するためにも重要である可能性があります。次世代のプロバイオティクスは、一部が酪酸を生成する可能性があるため、アレルギー反応に対して強力な予防効果をもたらす可能性があります。代わりに、COVID-19ワクチン製剤に酪酸を含めることで、ワクチンの悪影響を軽減することもできます。プレバイオティクス多糖類は、COVID-19の予防と治療のために提案されています(Barbosa and de Carvalho Junior,2021)。多糖類の種類ごとに、免疫系に刺激または抑制のいずれか異なる影響を与える可能性があります(Barbosa and de Carvalho Junior,2021)。これらの多糖類がCOVID-19ワクチンの有効性にどのように影響するかは、最初に動物モデルで調べることができます。 |
| シンバイオティクス製剤(すなわち、プロバイオティクス+プレバイオティクス)は、COVID-19ワクチンの有効性と副作用の予防に大きな影響を与える可能性があります。 最適な処方を進めることができます。 次世代のプロバイオティクス、特に酪酸産生細菌が含まれる可能性があります。 腸内細菌叢が腸内の免疫応答を高めるための主要なメディエーターである酪酸の生成を促進するために、いくつかのプレバイオティクスを追加することができます。 |
| 9. 結論 |
| 腸内細菌叢は、加齢とともに変化する可能性があることが認識されています(Zhang et al.,2021、Zhang et al.,2021)。 加齢に伴う炎症、免疫老化、ライフスタイルの栄養および身体活動の変化、薬物使用、慢性的な健康状態の存在など、加齢に伴う生理学的および外因性の変化があります(DeJong et al.,2020、Zhang et al.,2021、Zhang et al.,2021)。 これらの変更の範囲はまだ明らかにされていません。 進行しているのは、腸内毒素症(Ragonnaud and Biragyn,2021)が加齢に伴う要因であると認識されており、重度のCOVID-19感染症で観察されています(Chen et al.,2021、Prasad et al.,2021)。 |
| COVID-19ワクチンは現在、先進国で広く投与されており、COVID-19の新規症例を大幅に減らすようになっています。ただし、特に脆弱で高齢者であるか、基礎となる慢性疾患と診断されている脆弱な人々では、ワクチンの有効性をさらに高める必要があるかもしれません。いくつかの研究は、COVID-19ワクチンの有効性が、高齢または慢性疾患の患者で著しく低下することを示しています(Soiza et al.,2021)。ほとんどの副作用は軽微ですが、重篤な合併症や死亡を引き起こす副作用は、主要な未解決の問題を構成します。腸内毒素症は、COVID-19ワクチンの有効性と副作用に関連している可能性があります。確かに、ワクチン接種後の高齢の虚弱な患者の死亡を調査しているノルウェーの研究(Torjesen、2021)は、異生物性腸の関与のもっともらしさを増しています。リスク集団における腸内細菌叢の改善は、ワクチンの有効性を高め、重篤な副作用を軽減する可能性があります。腸内毒素症の影響は、感受性の高い個人や虚弱な高齢者の集団におけるCOVID-19ワクチンの非効率性と悪影響の主な要因である可能性があります(Lynn et al.,2021)。ビフィズス菌および酪酸などの細菌代謝産物による腸内細菌叢の調節(Chen and Vitetta,2020、Lynn et al.,2021)は、虚弱な高齢者におけるワクチンCOVID-19の有効性改善の補助剤としての実用的なアプローチである可能性があります。プレバイオティクスの有無にかかわらず、特に酪酸産生細菌を誘発する次世代プロバイオティクスの組み込みは、重要なアジュバントメディエーターであるCOVID-19ワクチンの有効性とワクチンの悪影響の抑制として、腸内細菌叢の存在量と多様性を改善するために必要な要件となる可能性があります。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
| この文献は、Journal of Functional Foods 87 (2021) 104850に掲載されたThe intestinal microbiota and improving the efficacy of COVID-19 vaccinationsを日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |