|
Duygu Agagündüz et al., |
|
概要 |
| 発酵乳製品は、健康を促進する可能性が十分に特徴づけられている有益な微生物であるさまざまな種類の生きた乳酸菌(LAB)の優れた供給源であります。伝統的に、発酵乳製品の食事摂取は、とりわけ抗菌活性や免疫系の調節を含むさまざまな健康促進効果に関連してきました。近年、新たな証拠は、非感染性疾患の予防と治療における乳製品乳酸菌の貢献を示唆しています。生きている細菌細胞またはその代謝物は、生理学的反応に直接影響を及ぼしたり、より複雑なコミュニケーションを仲介するシグナル伝達分子として機能したりする可能性があります。このレビューは、食品-腸-健康軸の概念を議論することにより、乳製品から分離された乳酸菌(乳製品乳酸菌)と人間の健康との間の相互作用に関する最新の知識を提供します。特に、乳製品乳酸菌のいくつかの生物活性とプロバイオティクスの可能性は、脳腸軸への関与に関して提供されています。そして、非感染性疾患は、主に肥満、心血管疾患、糖尿病、炎症性腸疾患、および癌の治療における可能性に焦点を当てています。 |
|
1.はじめに |
|
発酵乳と乳製品は、世界中の食生活のマイルストーンです。それらの栄養的および官能的特性を超えて、発酵乳製品の健康上の利点は長い間知られていました[1]。 特に、発酵乳製品は、さまざまな種類の生きた乳酸菌の優れた供給源であり、有益な微生物は、そのプロバイオティクスの可能性について十分に特徴付けられています[2]。プロバイオティクスは生きた微生物であり、適切な量を投与すると、宿主に健康上の利益をもたらします[3]。プロバイオティクスの摂取は、栄養素や結合部位を求めて病原体と競合し、抗菌代謝物をその場で生成することにより、腸および泌尿生殖器環境の共生微生物の組成を調和させます[4]。 さらに、プロバイオティクスは、宿主の免疫応答を調節し、炎症性腸疾患(IBD)で観察されるような炎症性胃腸状態の症状を制御することによって粘膜バリア機能の改善に貢献します[5]。あまり明白ではないのは、婦人科、生殖、代謝、心血管、骨粗鬆症、アポトーシス制御など、さまざまな人間の病気の予防と治療に乳製品乳酸菌が及ぼす可能性のある貢献です[6]。 |
| 乳酸菌株の複雑な酵素的遺産は、乳製品マトリックスまたは腸内のいずれかへのいくつかの生物活性代謝物の放出に寄与し、したがって、食物-腸-健康軸の魅力的な概念を促進します。これらの代謝物は、さまざまな生理学的プロセスに直接影響を与えるか、周囲の微生物へのシグナル伝達分子として機能します。たとえば、αおよびβカゼイン、アルブミン、およびグロブリンの異化作用から主に乳製品乳酸菌によって生成される暗号化された生物活性ペプチドは、抗菌剤、高コレステロール血症、オピオイドおよびオピオイド拮抗薬、アンジオテンシン変換酵素阻害薬、抗血栓薬、免疫調節剤、細胞調節剤、および抗酸化剤として報告されています[7]。ヨーグルト、培養発酵乳、ケフィアなどのプロバイオティクスを含む乳製品の摂取は、コレステロール代謝やアンジオテンシン変換酵素(ACE)阻害、抗菌活性、腫瘍抑制、創傷治癒の速度の増加、および免疫系の調節など、さまざまな健康上の利点に関連しているという証拠があります[8,9]。最近、ラクトバチルス菌株によって発酵された乳製品が、マウスモデルの腸骨軸を調節することが報告されました。これは、生きている乳酸桿菌細胞と同じ乳酸菌によって発酵された乳製品によって調整できる効果です[10]。食-腸の複雑な生態系では、乳製品の乳酸菌は、細菌全体またはその成分によって媒介される信号のネットワークを誘発する可能性があります。腸-脳-微生物叢軸は、神経、内分泌、免疫、および代謝のリンクの関与により、腸-微生物叢から脳へ、および脳から腸内細菌叢へのシグナル伝達を介した両側コミュニケーションシステムに基づいています[11]。予備研究で、Butlerと共著者は、乳製品の摂取に基づく食事の変化が乳酸桿菌の豊富さを促進し、脳腸モジュールアプローチを使用して予測神経活性の可能性を測定することにより、参加者の心理的状態との関連を示唆していることを観察しました[12 ]。したがって、最近の調査では、ミルク、ヨーグルト、ケフィアなどの乳製品の摂取が、Lactobacillusの個体数を増やすことによって腸内細菌叢を調節する可能性があることが示されました[13]、一方、プロバイオティクスのLactobacillus casei菌株を含む発酵乳製品を摂取すると、高齢者の呼吸器感染症の期間が短縮されます[14]。ただし、宿主の遺伝学、年齢、性別、ライフスタイルなどの他の要因と組み合わせた乳製品の消費の影響は、腸内細菌叢の組成と機能に影響を与えます。このレビューでは、一般に、乳製品から分離された乳酸菌(つまり、乳製品乳酸菌)と人間の健康との間の複雑な相互作用に焦点を当て、食品-腸-健康軸の新しい概念を提案しています。特に、この記事では、乳製品乳酸菌の発酵および/またはプロバイオティクスの可能性と、脳-腸軸および非感染性疾患(NDC)への関与に焦点を当てています。 そして主に、肥満、心血管疾患、糖尿病、炎症性腸疾患、および癌の治療におけるそれらの保護の可能性についてです。 |
| 2.乳酸菌の定義といくつかの特徴 |
|
乳酸菌は系統発生的および機能的に多様な細菌のグループであり[15]、微生物の食用培養物として広く知られています[16]。定義上、乳酸菌は系統発生的および機能的に多様な細菌の分類学的順序を構成します[15]。これらの多様な微生物には共通の特徴があります。炭水化物を発酵させる過程で最終的に乳酸を生成します[17–19]。伝統的に、それらは特に乳製品における人間の食品の発酵と密接に関連しています[16]。乳酸菌で見られる一般的な特性を表1にまとめています。 |

|
| 3.健康の観点からの乳製品における乳酸菌の重要性 |
| 乳酸菌は、発酵乳や乳製品の健康へのプラスの効果に重要な役割を果たします。 乳酸菌は、一部の乳製品に自然に含まれる場合があります。また、スターターカルチャーとして追加したり、不明確な問題がある場合でも、特に製品のプロバイオティクスの可能性のために機能性を高めるための新しい成分/添加物として追加したりすることもできます。乳酸菌は、 Lactobacillus, Lactococcus, Pediococcus, EnterococcusおよびStreptococcusなどの属を含む発酵乳製品によく見られる細菌のグループです[21]。プロバイオティクスとして使用されるほとんどの細菌は、乳酸菌種であるLactobacillus属(Lactobacillus acidophilus,Limosilactobacillus fermentum, Lacticaseibacillus casei, Limosilactobacillus reuteri, Lactocaseibacillus rhamnosus, Lactobacillus helveticus, Lactococcus lactis,Lactobacillus crispatus, Lactobacillus gasseri, Lactiplantibacillus plantarum)およびEnterococcus属(Enterococcusfaecalis,Enterococcusfaecium)に属しています[22]。乳酸菌スターターカルチャーは食品や牛乳の発酵を行うために長年使用されてきたため、これらの細菌は一般に「一般的に安全と認められている」(GRAS)と見なされています[23]。 |
| 乳製品の栄養価に影響を与える主な要因は、使用されるミルクベースの供給源(動物の種類、食事、年齢、泌乳期間など)と食品加工プロセス(温度、保管条件など)です。さらに、発酵に使用されるスターターカルチャーおよび/またはプロバイオティクスタイプは、発酵乳製品の栄養価に直接影響する要因です[24]。プロバイオティクスは一般に、乳酸菌ベースの従来のプロバイオティクス、非乳酸菌プロバイオティクス、および次世代プロバイオティクスの3つのグループに分けられます。発酵乳製品には他の微生物が使用されていますが、乳酸菌の方が優れています。 乳酸菌が使用される主な乳製品は、発酵乳、ヨーグルト、乳児用調製粉乳、チーズ、バターとクリーム、およびアイスクリームです[25,26]。 一般に、乳製品は、自然に検出されるか、後で追加されるプロバイオティクスとして、乳酸菌の主要な食事源と見なされます[27]。食品中のプロバイオティクス株の健康への影響を保証できる特定の細胞数レベルはありませんが、プロバイオティクスの有益な効果から利益を得るのに十分な量として、少なくとも106〜108 cfu / gが報告されています[25]。これは、プロバイオティクスの可能性を示すことができるカルチャーの存在が、製品がプロバイオティクスになることを保証するものではないことを明確に示しています。多くの要因が乳製品のプロバイオティクスの生存率と安定性に影響を与えます。これらは、滴定可能な酸性度、pH、均質化、溶存酸素含有量、H2O2、保存温度、プロバイオティクスの種類、乳酸濃度、および関連生物の種と菌株です[28]。ただし、プロバイオティクスの実行可能性を維持するためにいくつかの方法が開発されています。これに関連して、マイクロカプセル化およびプレバイオティクス添加技術を使用してプロバイオティクスの生存率を高めるための研究があります[28,29]。 |
| 発酵乳製品では、Lactobacillus, Lactococcus, Leuconostoc, Pediococcus, Bacillus,Propionibacterium およびBifidobacterium属に属する細菌が主に顕著です[30]。Lactobacillus属は、314種の最大の属であり、過去20年間でプロバイオティクスと最も関連性の高いキーワードになっています。しかし、プロバイオティクスとしていくつかの乳酸菌属を受け入れることについて、科学者の間でコンセンサスはありません[31]。文献で最も研究されているプロバイオティクス乳酸菌のいくつかは、 Lactocaseibacillus rhamnosus GG (ATCC53103), Lactocaseibacillus rhamnosus HN001, Lacticaseibacillus casei Shirota,Lacticaseibacillus casei Zhang, Lactobacillus acidophilus NCFM, Lactobacillus acidophilus LA-5 および Lacticaseibacillus casei DN-114 001です。最近、Limosilactobacillus reuteri およびLactobacillusjohnsoniiなどの新しいプロバイオティクス乳酸菌が、機能性乳製品の開発に使用されています[25,32]。ミルクおよび乳製品のコーデックス委員会の定義によると、ヨーグルトおよびその他の発酵乳製品の特定のスターターカルチャーを図1に示します[33]。プロバイオティクスの有益な効果は菌株特異的であるため、同じ種の異なる菌株は宿主に完全に異なる効果を引き起こす可能性があります。したがって、新しい乳酸菌株のプロバイオティクスの可能性と、乳製品のよく知られたスターターカルチャーを理解するには、さらに多くの研究が必要であると述べられています[34]。 |
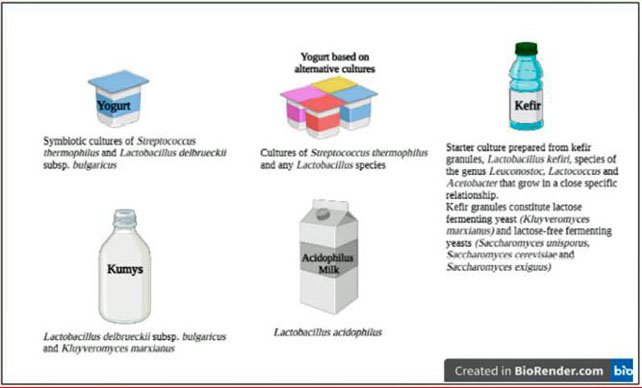 |
| 図1.一部の乳製品の特定のスターターカルチャー(Codex Alimentarius、2011年から採用) |
| 乳製品から分離されたいくつかの乳酸菌株、それらの潜在的な生物活性/プロバイオティクスの可能性、および安定性の問題を表2にまとめています。発酵乳製品から分離された乳酸菌株は、一般に低酸性条件とさまざまな胆汁酸濃度で生存します。ただし、一部の乳酸菌は、同じ条件下で低い安定性を示すことも報告されています[35]。さらに、多くの乳酸菌は、ヒトの腸細胞に接着する優れた能力と重要な自己凝集特性、および抗菌作用と免疫調節作用を持っていると報告されています[36–38]。入手可能なデータは、新しい機能性食品生産における乳酸菌の高い可能性を裏付けています。ただし、乳業での新しいプロバイオティクス製品の開発において、プロバイオティクス菌株と新規プロバイオティクス菌株の理想的な組み合わせと環境を選択するために、さらなる研究が推奨されます。 |
 |

|
| 近年のヒトの微生物叢とプロバイオティクスに関する重要な研究の結果、腸はその微生物叢により「第二の脳」として新たな評判を得ています[46]。間違いなく、脳-腸軸の発見がこれの根底にあります。脳-腸相関(HPA)は、腸管神経系と中枢神経系を接続する双方向通信ネットワークです。このネットワークは解剖学的であるだけでなく、内分泌、体液性/代謝、および免疫コミュニケーション経路も含まれます[47,48]。この通信ネットワークには、中枢神経系(CNS)、脳と脊髄の両方、自律神経系(ANS)、腸管神経系(ENS)、腸と脳を互いに接続する視床下部-下垂体-副腎軸(HPA)が含まれます [11,47]。 このように腸と脳の間に確立された通信ネットワークは、腸の活動や胃腸機能、代謝性疾患、認知能力、メンタルヘルスなどの多くの重要な健康パラメータに関連している可能性があることが示唆されています[46,47]。 |
| 腸内細菌叢は、人間の脳のように、腸管神経系と中枢神経系を調節する能力を持ち、多くの神経伝達物質を生成する能力を持っています[49]。腸内細菌叢によって局所的に産生される神経伝達物質の中には、γ-アミノ酪酸(GABA)、ノルアドレナリン、ドーパミン、セロトニン(5-ヒドロキシトリプタミン-5-HT)、メラトニン、ヒスタミン、アセチルコリンなどの局所神経伝達物質があります。内腔に生物学的に活性なカテコールアミンが形成され、それによって神経系の活動に影響を与える可能性があります[50、51、52]。この調節は、迷走神経、脳-腸相関、炎症性サイトカイン、神経伝達物質など、多くのメカニズムによってもたらされます[53]。さらに、乳酸桿菌などの特定の微生物によるカプサイシン感受性神経線維のバニロイド受容体と相互作用する一酸化窒素などのいくつかの分子の生成は、この調節に寄与します[54]。この変調は、遺伝学、ライフスタイルの習慣(栄養、薬物使用、運動など)、および多くの環境要因(ストレス、恐怖、社会的相互作用など)の影響を受けます。腸内細菌叢の組成は、この調節において重要な役割を果たします[49]。実際、腸内細菌叢における食事療法の有効性に関する証拠が近年増加しているため、双方向通信として知られる「脳-腸軸」ネットワークは、「食物-腸-脳軸」として3方向と呼ばれるようになります[48,55]。 |
| この軸に関連する人間に関する限られた研究は、ほとんどが乳酸菌で行われ、精神的パフォーマンス、心理的および関連する免疫学的パラメーターに関連していることがよくあります[47,56,57]。 Lactobacillus helveticus IDCC3801を含む発酵乳製品は、健康な高齢者(60〜75歳)で12週間後に認知能力を改善することが確認されました[56]。別の研究では、Lacticaseibacilluscasei DN-114001で発酵させた乳製品とヨーグルト培養を6週間定期的に摂取すると、大学生の免疫応答(リンパ球とCD56細胞数)が学術的ストレス下で変化したことが報告されました[57]。 Lactobacillusは、低温殺菌されていないミルクと製品の消費後に増加したこともわかりました(対照としての低温殺菌されたミルクと比較せずに)。脳-腸相関アプローチを使用して推定神経活性電位を測定することによって決定されるように、微生物叢のプロファイルと機能、および糞便の吉草酸のレベルが増加します[12]。したがって、これらすべての研究の結果は、牛乳および乳製品に由来する乳酸菌がこの軸で重要な役割を果たし、したがって健康パラメータに関連していることを示しました。ただし、乳酸菌カウントと精神的および心理的測定値との相関関係は報告されていません。 したがって、脳-腸軸に対する乳製品乳酸菌の直接的な影響の証拠はまだ不足していると言えます。 |
| 4. 非感染性疾患の調節における乳製品乳酸菌の関与 |
| ミルクと乳製品は、味、食感、栄養価の点で多種多様な食品グループであるため、世界中で60億人以上が消費しています[58]。体重や胴囲の減少、2型糖尿病(T2DM)、高血圧、心血管疾患(CVD)のリスクの低減など、牛乳とその製品の健康への有益な効果を支持する研究の数は増えています。近年、これらの効果を支持しない研究が文献にあります[59,60,61,62]。 これらの一貫性のない結果は、乳製品間の栄養素の多様性の大きな違いによるものと考えられています[58]。 乳製品の発酵やこの発酵プロセスで使用されるスターターカルチャー、発酵の期間、周囲温度などの多くの要因が、この栄養素とフレーバーの多様性の原因です[63]。 |
| 発酵乳製品は、発酵プロセス中に形成される微生物代謝物(生体または生物活性効果)の影響、およびそれらの組成から分離された特定の乳酸菌株のプロバイオティクス効果により、健康促進効果を示す場合があります[64,65]。発酵プロセス中に乳酸菌活性によって形成される最も重要な生物活性化合物は、ペプチド、エキソポリサッカライド(EPS)、バクテリオシン、いくつかのアミラーゼ、プロテアーゼ、リパーゼ酵素、および乳酸であると言われています[66]。ただし、すべての乳酸菌株がこれらの化合物のすべてを生成できるわけではありません。一部の乳酸菌株は、アンジオテンシン変換酵素阻害ペプチドを産生することにより降圧効果を示しますが、一部の菌株は、エキソポリサッカライドを産生することにより、抗糖尿病、コレステロール低下、抗腫瘍、および免疫調節効果を示します[66,67,68]。このため、非感染性疾患の予防と治療のサポートに効果的な乳製品乳酸菌株を調べることは非常に重要です。この点で、非感染性疾患は乳製品の乳酸菌株に関連しており、これらの細菌によって調節されると考えられています(図2)。 |
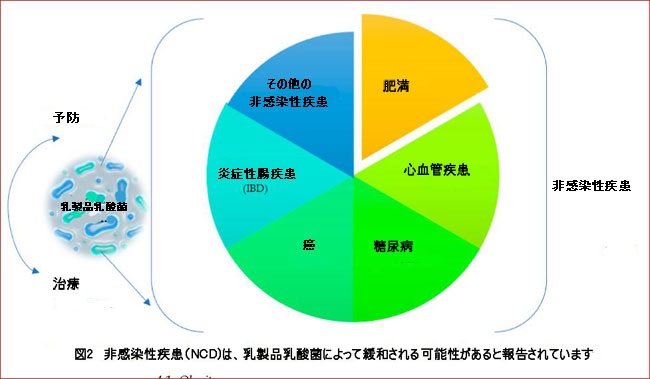 |
| 4.1 肥満 |
| 肥満は、健康を損なう可能性のある異常または過剰な脂肪蓄積として定義され、エネルギー消費の減少やエネルギー摂取の増加などの多くの要因から生じます[69,70]。これは、世界中の先進国および発展途上国における深刻な公衆衛生問題であり、その有病率は日々増加しています[71]。世界保健機関(WHO)は、肥満の有病率は1975年以来ほぼ3倍になっていると述べています[70]。 2型糖尿病、高血圧、心臓病などの一般的な肥満関連の併存疾患は、社会的および財政的システムに影響を及ぼします[71,72]。このため、効果的な治療法を適用することによる肥満の治療と予防は、個人の健康と生活の質、そして国の経済の両方にとって非常に重要です。同様に、肥満から保護し、その治療をサポートすることができる食品や栄養素の研究は、近年増加しています。これらの食品の中にある発酵乳製品は、発酵に使用される特定のスターター乳酸菌株のプロバイオティクスの可能性と、さまざまな生体代謝物(エキソポリサッカライド、生物活性ペプチドなど)の形成によってこれらの効果を示すと言われています[64]。 |
| 乳酸菌活性の結果として形成されるタンパク質加水分解物は、グルカゴン様ペプチド1(GLP-1)、グルコース依存性インスリン分泌性ポリペプチドまたは胃抑制ペプチド(GIP)、コレシストキニン(CCK)などの腸ホルモンに対する刺激効果を示すことができます。これらは食物摂取に反応して腸で分泌され、満腹信号としても機能します[73]。 高脂肪食(HFD)を摂取し、発酵ホエー飲料(LactiplantibacillusplantarumDK211およびLactococcuslactis培養)を摂取しているマウスを、高脂肪食、食物摂取量、血糖値、血清インスリン、レプチン、およびグレリンレベルに加えて比較した研究では、消費されたホエイ飲料は、高脂肪食のみを消費したマウスと比較して有意に低いことがわかりました[72]。 |
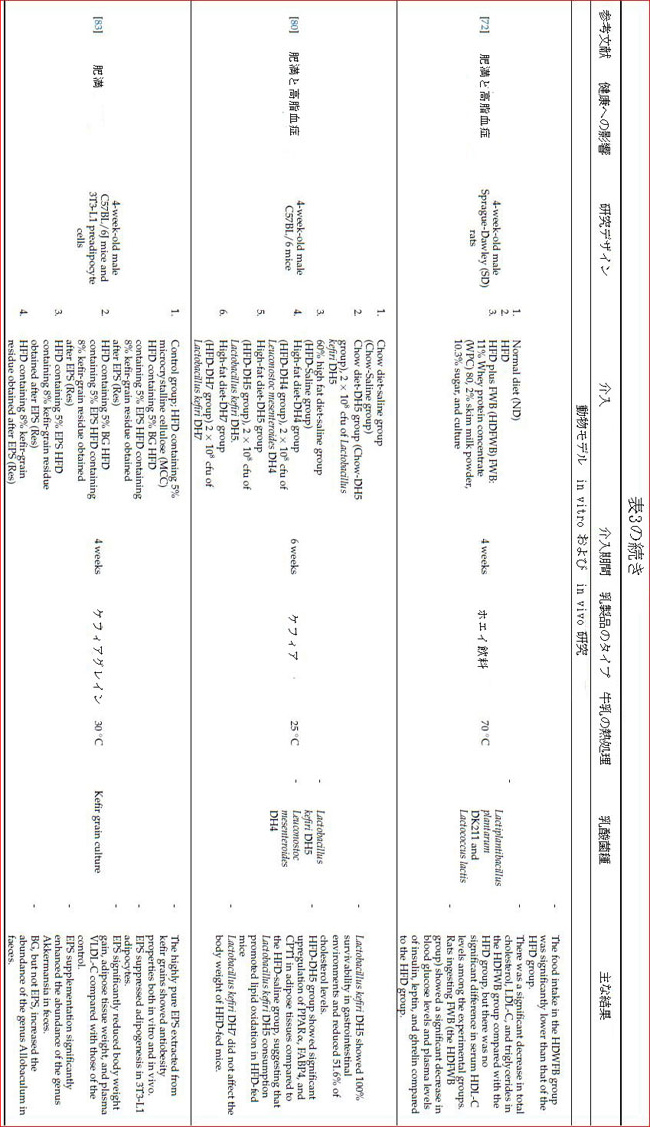
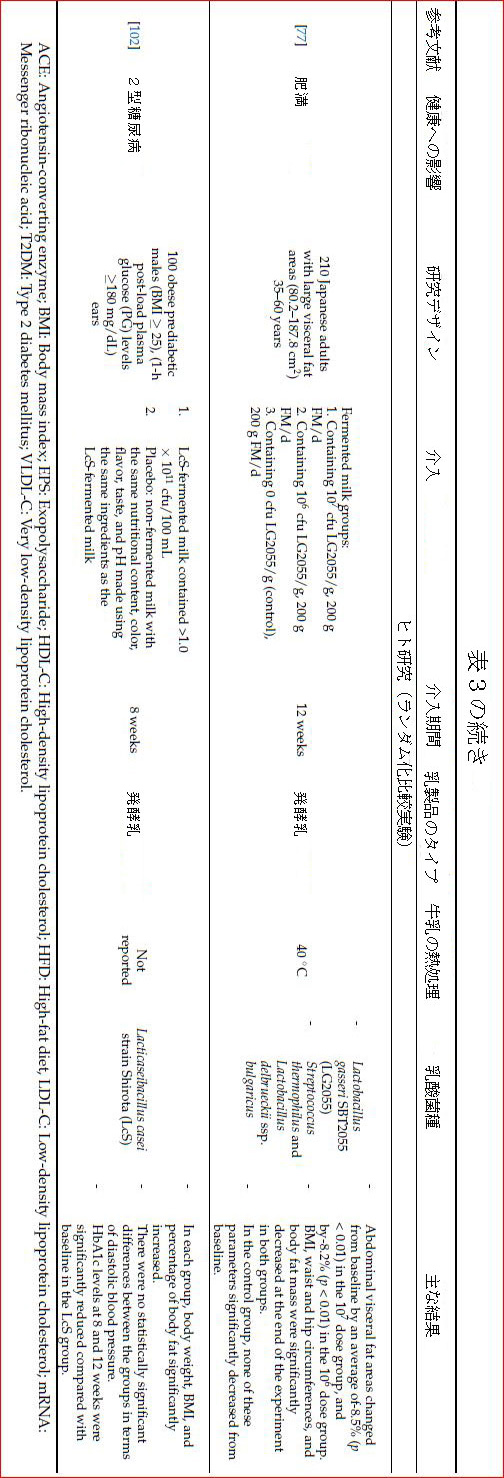
| 肥満と食事による高脂肪摂取は、ファーミキュラス/バクテロイデス門の比率の増加と腸内毒素症の発症を引き起こし、腸透過性の増加、代謝性内毒素血症、脂肪組織の炎症、および代謝性疾患を引き起こします[74,75,76]。さらに、腸内細菌叢における病原菌の増加は、上皮細胞からのリポ多糖(LPS)の産生と分泌を刺激します[75]。 リポ多糖は、脂肪細胞で炎症性サイトカインの放出を引き起こすサイトカイン受容体に結合することにより、代謝を炎症過程に導くことができます。プロバイオティクス特性を備えた乳製品乳酸菌株は、病原菌と競合し、腸内毒素症および炎症関連遺伝子発現のこれらの影響を軽減する可能性があります[75]。KadookaらはLG2055(訳者注:ガセリ菌SP株)を含む発酵乳を106および107cfu / g /日の用量で内臓肥満のある人に摂取すると、内臓脂肪症、肥満度指数(BMI)、体脂肪量、ウエスト、および股関節周囲が大幅に減少することが示されました[77 ]。 LG2055のこの効果は、腸上皮細胞(IEC)による認識と、腸上皮細胞の抗炎症および上皮の完全性保護メカニズムのサポートによって説明されます[78]。乳酸菌によって産生されるバクテリオシンは、腸内毒素症を引き起こす腸内細菌叢の細菌に対して抗菌効果を示すことにより、潜在的な抗肥満効果をもたらします[79]。 |
| ケフィア培養から分離された乳酸菌株、特にLactobacillus kefiri DH5株は、高脂肪食を与えられたマウスの体重と肥満を減らし、胃腸系(GIS)全体で生存率を維持しました[80]。Lactobacillus kefiriDH5は、腸管腔でのコレステロール吸収を減らし、PPARα(訳者注:ペルオキシソーム増殖剤活性化レセプター)、カルニチンパルミトイルトランスフェラーゼ-I(CPT1)、および脂肪組織で脂肪酸結合タンパク質4(FABP4)の発現を増加させることにより、これらの抗肥満効果を発揮することが確認されています。このメカニズムをさらに解明するために、カルニチンパルミトイルトランスフェラーゼ-Iおよび脂肪酸結合タンパク質4遺伝子はPPARαによって媒介されるため、PPARα活性の増加はこれら2つの標的遺伝子のアップレギュレーションを誘導します[80]。カルニチンパルミトイルトランスフェラーゼ-Iはミトコンドリアの外膜にある酵素で、脂肪酸のβ酸化のためにアシル基の輸送を触媒します[81]。脂肪酸結合タンパク質4は、脂肪生成を抑制し、脂肪組織の脂肪分解を促進する細胞内タンパク質です。その結果、Lactobacillus kefiri DH5を含むケフィアの摂取は、カルニチンパルミトイルトランスフェラーゼ-Iおよび脂肪酸結合タンパク質4遺伝子の発現をアップレギュレートし、それによって脂肪酸のβ酸化と脂肪分解を増加させ、代謝における脂質生成を減少させ、抗肥満効果をもたらす可能性があります[80]。 |
| 乳製品乳酸菌は膵臓リパーゼ酵素を抑制し、エネルギー源として使用する脂肪の吸収を防ぎ、糞便中の排泄を増加させます。この抑制機能を備えたバクテリアを含む製品は、肥満の治療に使用できると言われています[82]。 Lacticaseibacillus casei、Limosilactobacillus fermentumおよびLactobacillus helveticusのさまざまな菌株が、膵臓リパーゼを2〜25%阻害できることが確認されました。 発酵乳製品であるダヒから分離されたLacticaseibacillus casei(NK9)株は、最も高い阻害を示しました[82]。ただし、同じ細菌種の異なる菌株間でもまったく異なる結果が得られました。 |
| 抗肥満効果は、発酵プロセス中に一部の乳製品乳酸菌株によって生成されるエキソポリサッカライドの生物学的効果の結果としても発生する可能性があります。ケフィア粒に含まれるLactobacillus kefiranofaciensによって生成される水溶性ヘテロ多糖エキソポリさっぱライドであるケフィランは、生物学的効果が最も研究されている乳製品エキソポリサッカライドの1つです。高脂肪食に加えてケフィア分離エキソポリサッカライドを摂取したマウスは、高脂肪食のみを摂取したマウスよりも体重と脂肪の増加が有意に低かった[83]。エキソポリサッカライドの抗肥満効果は、一般的にその粘性と水溶性の構造に関連しています。しかし、エキソポリサッカライドの抗肥満効果は、エキソポリサッカライドグループとより粘度の高いベータグルカンを投与したグループの体重と脂肪の減少を比較すると、大幅に優れていることがわかりました[83]。さらに、Akkermansia属は、エキソポリサッカライドサプリメントを摂取したグループの糞便で増加したことが確認されました[83]。Akkermansia属の種は、ムチン層の厚さと杯細胞の厚さを増加させ、代謝内毒素血症を減少させることにより、腸の完全性を保護し、肥満を防ぐプロバイオティクス効果があると言われています[84]。言い換えれば、その粘度と体積増加効果に加えて、その潜在的なプレバイオティクス特性は、エキソポリサッカライドの抗肥満メカニズムにおいても重要な位置を占めています[83]。乳製品乳酸菌株によって生成されるエキソポリサッカライドは、ヒトの消化酵素によって加水分解されることなく結腸を通過し、腸上皮と直接接触し、生物学的活性を持つ代謝物を生成することにより、これらのプレバイオティクス効果を示すことができます[85]。 |
| また、乳製品の乳酸菌株は、発酵プロセス中にミルクの組成に含まれる共役リノール酸(CLA)の量を増やすことにより、抗肥満効果をサポートすることも示されています[86]。共役リノール酸は、食物摂取、増殖、脂肪細胞分化、脂質生成を減少させ、脂肪分解、脂肪酸のβ酸化、エネルギー消費を増加させることにより、体重を減らす効果があります[86]。 |
| 4.2 心血管疾患 |
| 心血管疾患(CVD)は、世界の主要な死因の1つであり、その有病率は日々増加しています。 それらはまた、最も一般的な死因であり、毎年推定1,790万人の死亡を占めています(全死亡の31%)[87]。アメリカ予防心臓学会(ASPC)は、心血管疾患の最も重要な危険因子は、不健康な食事、運動不足、脂質異常症、高血糖、高血圧、肥満、患者間の個人差(年齢、人種/民族、性別)、血栓症/喫煙、腎機能障害、および遺伝性/家族性高コレステロール血症であると指摘しました[88]。 近年行われた研究では、発酵乳製品の消費と心血管疾患のリスクとの間に負の関係があることが示されました[59,62]。 心筋梗塞後の患者では、脂肪含有量に関係なく、ヨーグルト消費量が25g / 日増加するごとに、心血管疾患関連の死亡リスクが4%減少し、全体的な死亡リスクが2%減少することがわかりました。 一方、ミルクやチーズなどの他の乳製品ではそのような関連性は観察されませんでした[89]。 |
| 心血管疾患の病態生理に関与する最も重要な危険因子の1つは、高コレステロール血症です。 臨床研究は、プロバイオティクス乳酸菌株の定期的な投与がコレステロール低下作用をもたらす可能性があることを示しています[90]。人間の腸から分離された乳酸菌およびビフィズス菌はコレステロール同化効果を示す可能性があり、乳酸菌によって体内のコレステロール含有量が1%減少すると、心血管疾患の発生率が2%から3%減少します[91]。さらに、Lactobacillus acidophilus L1株を含む発酵乳製品を定期的に摂取すると、血中コレステロールが2.9%減少することが報告されています。したがって、冠状動脈性心臓病のリスクを6〜10%減らす可能性があります[92]。さらに、Lacticaseibacillus paracasei subsp paracasei NTU 101、Lactiplantibacillus plantarum NTU 102およびLactobacillus acidophilus BCRC17,010株で発酵させた牛乳を摂取すると、総コレステロール、低密度リポタンパク質コレステロール(LDL-C)、高比重リポタンパク質コレステロール、密度リポタンパク質コレステロール(HDL-C)、 高コレステロール食を与えられた高脂血症マウスのトリグリセリドレベルの低下に効果的であることがわかりました[93]。 |
| プロバイオティック特性を示す乳酸菌は、基本的に、コレステロール同化、細胞表面や膜などの細胞成分へのコレステロールの結合/付加、胆汁酸塩ヒドロラーゼ(BSH)による胆汁酸の酵素的脱共役、プロバイオティクスによって生成される短鎖脂肪酸(SCFA)によるコレステロールデノボ合成の抑制などのメカニズムを通じてコレステロール低下効果を示します[94,95]。ただし、発酵乳製品のすべての乳酸菌株がコレステロール低下作用を示すわけではありません。 Streptococcus thermophilusとEnterococcusfaecium菌株を一緒に使用して発酵させたヨーグルトは血清コレステロールレベルの有意な減少(8.4%)をもたらしましたが、StreptococcusthermophilusとLactobacillus acidophilusまたはLactobacillus rhamnosus菌株を一緒にまたは単独で使用して製造されたヨーグルトは低コレステロール血症効果を示しませんでした [96]。消化およびコレステロール低下能力におけるinvitro生存のための58の潜在的なプロバイオティクス乳酸菌株の検査の結果、最高の性能を発揮する菌株は、Lacticaseibacillus casei, Lacticaseibacillus paracasei, Lactiplantibacillus plantarum, Enterococcus faecium, and Enterococcus lactisおよびEnterococcus lactisであり、いくつかの菌株は、ブロス中のコレステロール値を42〜55%低下させました [97]。 別の研究では、プロバイオティックヨーグルト(Streptococcus thermophilus,Lactobacillus bulgaricus,Lacticaseibacillus casei subsp casei; 108cfu/g)と従来のヨーグルト(Streptococcus thermophilus,Lactobacillus bulgarus; 10 8 cfu/g)で、グループ間で有意差は観察されませんでした。したがって、プロバイオティクスと従来のヨーグルトの両方を4週間定期的に摂取すると、健康な女性の血漿中の脂質プロファイル(総/ HDLおよびLDL / HDLコレステロール比)にプラスの効果があることが示されています[98]。 |
| プロバイオティクス乳製品乳酸菌株であるLacticaseibacilluscaseiShirota(LcS)の経口投与は、動物モデルにおけるインスリン抵抗性、耐糖能異常、2型糖尿病、脂肪肝、高脂血症などの肥満関連代謝異常を改善することが示されています[99,100,101]。 LcS YIT9029で発酵させたミルクを消費する肥満の前糖尿病患者では、この乳製品乳酸菌株は、ステロールを結合および/または同化することによって食事中のコレステロール吸収を減らし、高コレステロール血症の個人を防ぐ可能性があります。 100mL /日の発酵乳を8週間摂取した人は、プラセボ群と比較して、総LDLおよび非LDLコレステロールレベルが有意に低かった[102]。 |
| 一部の乳酸菌株には胆汁酸塩ヒドロラーゼ(BSH)酵素があり、この酵素は抱合型胆汁酸塩の遊離胆汁酸塩とアミノ酸残基への加水分解を触媒します[103,104]。 腸管腔での胆汁酸塩ヒドロラーゼの発現は、溶解度と吸収性の高い抱合型胆汁酸塩を溶解度の低い遊離胆汁酸塩に変換します。したがって、胆汁酸塩の再吸収と再利用が減少し、肝臓でのコレステロールからの胆汁酸塩のデノボ合成が増加します。この合成には、血液から肝臓へのコレステロール輸送が必要であり、それによって総血清コレステロールレベルが低下します[103]。 たとえば、Lactiplantibacillus plantarum SC70およびSC80株で発酵させた乳製品は、高いリパーゼ阻害(>35%)および胆汁酸塩ヒドロラーゼ活性を示すことにより、コレステロール値を下げることができます[103]。 さらに、Lactiplantibacillus plantarumGKM3をマウスの高脂肪食に6週間添加した結果、肝臓の総脂質、コレステロール、およびトリグリセリドのレベルが低下し、糞便中の脂質、コレステロール、およびトリグリセリドの排泄が大幅に増加しました[105]。糞便中のコレステロールと脂質の排泄の増加は、リパーゼ阻害と胆汁酸塩ヒドロラーゼ酵素活性の効果を示唆しています。 |
| 乳製品乳酸菌が血中コレステロールレベルの調節に効果的である別のメカニズムは、3-ヒドロキシ3-メチルグルタミルCoA(HMG-CoA)レダクターゼ酵素を阻害することによってコレステロール合成を抑制することです。乳製品の乳酸菌種の1つであるLactobacillusacidophilusは、体内の内因性コレステロール生合成に関与する律速酵素である3-ヒドロキシ3-メチルグルタミルCoAレダクターゼを阻害し、この酵素は腸内の胆汁酸を脱共役することによってコレステロール濃度の低下を引き起こす可能性があります[106]。 |
| ただし、一部の細菌はコレステロール表面に付着し、コレステロールが吸収されにくくなります。 ある研究では、ケフィアから分離されたLactobacillus kefiriJK17の活性細胞と死細胞のコレステロール結合特性を比較したところ、コレステロールが死細胞膜に結合していないことが観察されました。一方、少量のコレステロールが活性細胞に結合していました[107]。このメカニズムに関連する研究は多くありませんが、現在の研究の結果は、乳製品乳酸菌がコレステロール結合特性を示すために腸内での生存能力を維持する必要があることを示唆しています。 |
| LDLの酸化的修飾は、アテローム性動脈硬化症の開始と進行に重要な役割を果たします。LDLには細胞毒性の可能性があり、アテローム性動脈硬化症のプラークにおける内皮細胞の損傷とマクロファージの変性の原因となります[108]。このため、LDL酸化の指標であるマロンジアルデヒド(MDA)-LDLおよび/またはマロンジアルデヒド-LDL /LDL-コレステロールレベルは、心血管疾患の優れたリスク指標となる可能性があります[109,110]。 二重盲検プラセボ対照試験では、抗酸化活性の高いStreptococcus thermophilus YIT 2001を含む発酵乳の摂取が、LDL酸化と血圧に及ぼす影響を調べました。発酵乳グループの血清マロンジアルデヒド-LDL、マロンジアルデヒド-LDL / LDLコレステロール、収縮期血圧(SBP)、および拡張期血圧(DBP)値の低下は、対照グループ(スキムミルク)よりも有意に高いことが観察されました。 [108]。別の研究では、酸化ストレスとマロンジアルデヒドレベルが減少し、血清GSH-Px(訳者注:グルタチオンペルオキシダーゼ)、SOD(訳者注:活性酸素を除去する酵素)、およびCATレベルは、老齢マウスでLactiplantibacillus plantarum YW11によって形成された高用量(1日あたり50 mg / kg)のエキソポリサッカライドへの曝露の結果として増加しました。 [90]。エキソポリサッカライドの生成は、コレステロール結合効果を示し、抗酸化防御をサポートすることにより、高脂血症を軽減する可能性があります。エキソポリサッカライド産生および非エキソポリサッカライド産生Lactococcus lactis subsp cremorisのコレステロール結合能力を比較した研究は、エキソポリサッカライド産生株が非エキソポリサッカライド産生株よりも多くのコレステロールに結合できることを示しました。これらの多糖類は、可溶性食物繊維と同様に、コレステロールを結合し、その吸収を減らし、糞便中排泄を増やすことによって血中コレステロールレベルを下げることができると言われています[111]。 |
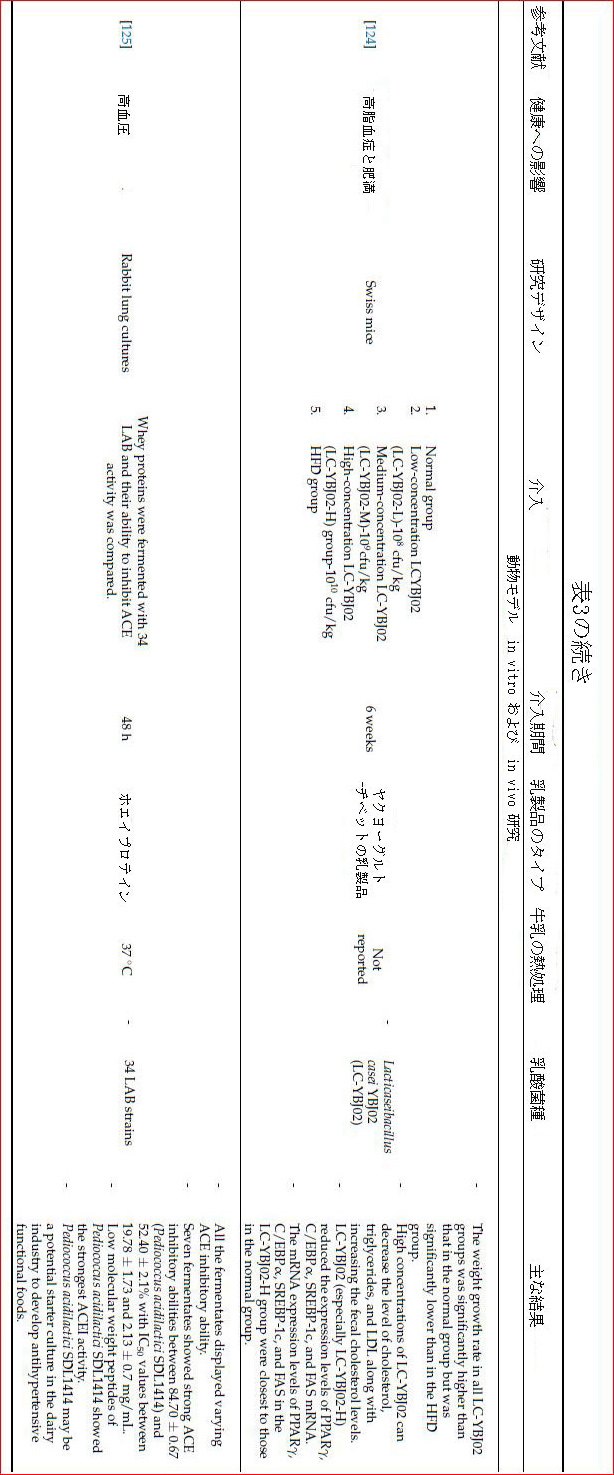
| 高血圧は、血圧値が正常限界を超えることを特徴とする慢性変性疾患です[112]。さらに、それは他の心血管疾患、脳卒中、腎不全、脳血管事故、および他の多くの合併症の発症の重要な危険因子です[113]。 フラミンガム心臓研究は、拡張期血圧の2mmHgの減少が、高血圧の有病率の17%の減少、脳卒中のリスクの15%の減少、および冠状動脈性心臓病のリスクの6%の減少と関連していることを報告しました[114]。アンジオテンシン変換酵素は血圧の制御に重要な役割を果たし、アンジオテンシンIをアンジオテンシンIIに変換します。アンジオテンシンIIは血管収縮作用を持ち、ブラジキニンやカリジンなどの血管拡張ペプチドを加水分解して血圧を上昇させます[113]。この酵素を阻害すると、血管拡張反応が高まり、血圧を下げる効果が見られます。 発酵乳製品の発酵過程で乳酸菌によって形成される生理活性ペプチドは、アンジオテンシン変換酵素を阻害することにより血圧調節効果があると言われています。これらのバクテリアは、タンパク質分解効果を示すことによってミルク組成物中のタンパク質を加水分解し、Val-Pro-Pro(VPP)やIle-Pro-Pro(IPP)などのアンジオテンシン変換酵素阻害剤オリゴペプチドを形成します。同時に、タンパク質分解活性により、カソキニンやラクトキニンなどの降圧ペプチドを生成することができます[1,64]。 Lactococcus lactis NRRL B-50571で発酵させたミルクを摂取し、カプトプリル(アンジオテンシン変換酵素阻害剤)を6週間摂取すると、収縮期血圧と拡張期血圧に同様の効果があり、これらの値は対照群とは有意に異なっていた。さらに、6週間の終わりに、発酵乳製品を消費したグループの収縮期血圧値は49.9±14.2mmHg減少しましたが、カプトプリルを消費したグループの収縮期血圧値は45.2±23.6mmHg減少しました。血漿アンジオテンシン変換酵素レベルは5.5倍低く、血漿一酸化窒素(NO)レベルは対照群と比較して発酵乳とカプトプリルを投与された患者で1.6倍高かった[115]。一酸化窒素は、内皮機能と血管拡張を改善し、血圧を制御する重要な生体調節分子です。体内の炎症過程と酸化ストレスの増加は、内皮一酸化窒素シンターゼ(eNOS)酵素活性を低下させ、内皮機能障害を引き起こし、一酸化窒素バイオアベイラビリティを低下させ、その結果、高血圧を引き起こす可能性があります[116]。 |
| ある研究では、Val-Pro-ProとIle-Pro-Proを5日間適用した結果、内皮一酸化窒素シンターゼ遺伝子の有意な誘導(1.89倍)が一酸化窒素レベルを増加させ、シクロオキシゲナーゼ(COX-1)遺伝子の発現をわずかに増加させ、NF-kB(訳者注:転写因子として働くタンパク質複合体)およびPPARγ(訳者注:ペルオキシソーム増殖因子活性化受容体γ、核内受容体スーパーファミリーに属するタンパク質であり、転写因子としても機能する)遺伝子の発現を減少させることが決定された[117]。 5週間後に発見された高血圧前症患者、収縮期血圧(116.55±12.26 mmHg vs.124.77±11.04mmHg)および拡張期血圧(80.7±9vs.84.5±8.5mmHg)は、対照群と比較して、発酵乳を摂取した人で有意に低かった[118]。 介入終了時の収縮期血圧と拡張期血圧の値が、発酵乳を消費している人のベースラインと比較して大幅に減少したとしても、この効果は介入が中止されてから1週間後に消えました[118]。 別の研究では、高血圧ラットは、Lactococcus lactis NRRL B-50572 および Lactococcus lactis NRRL B-50571 株、カポプチル、および水で4週間発酵させたミルクを摂取しました。その結果、Lactococcus lactis NRRL B-50572 発酵乳を摂取したグループの収縮期血圧および拡張期血圧値は、Lactococcus lactis NRRL B-50572および水を摂取したグループと比較して有意に低かった。一方Lactococcus lactis NRRL B-50571および水を消費するグループも同様であることがわかりました。 しかし、介入の終わりに、最も低いLDL-コレステロールおよびTG値(中性脂肪)は、Lactococcus lactis NRRL B-50572FMを消費するグループにあることがわかりました[119]。脂質異常症はまた、内皮損傷を引き起こすことによって高血圧の発症に役割を果たす可能性があります[115]。それにより異なるメカニズムにより、両方のLactococcuslactis 菌株は降圧特性を有し、心血管疾患に対する保護効果を発揮すると結論付けることができます。 ただし、この結果をすべてのLactococcus lactis株について一般化することはあまり正確ではありません。 Lactobacillus helveticusNK1、Lactocaseibacillus rhamnosus F、およびLimosilactobacillus reuteriLR1株で発酵させたミルクのタンパク質分解、抗酸化、およびアンジオテンシン変換酵素阻害活性を調べた研究では、細菌株のタンパク質分解活性に応じて抗酸化活性が増加しました。一方、アンジオテンシン変換酵素阻害剤とタンパク質分解活性の間にそのような関連はありませんでした。さらに、Lactobacillus helveticusNKのタンパク質分解活性はLactocaseibacillus rhamnosusFの2倍でしたが、これらの菌株のアンジオテンシン変換酵素阻害は類似していることがわかりました。さらに、発酵乳サンプルのペプチドプロファイルを調べたところ、アンジオテンシン変換酵素阻害特性はαS2-カゼインのC末端からなるペプチドと関連していた[48]。カゼインおよびカゼイン画分から生成されたペプチドの強力な降圧およびアンジオテンシン変換酵素阻害特性は、それらの構造においてイオン性カルシウム(Ca2 +)と関連しています[120]。 |
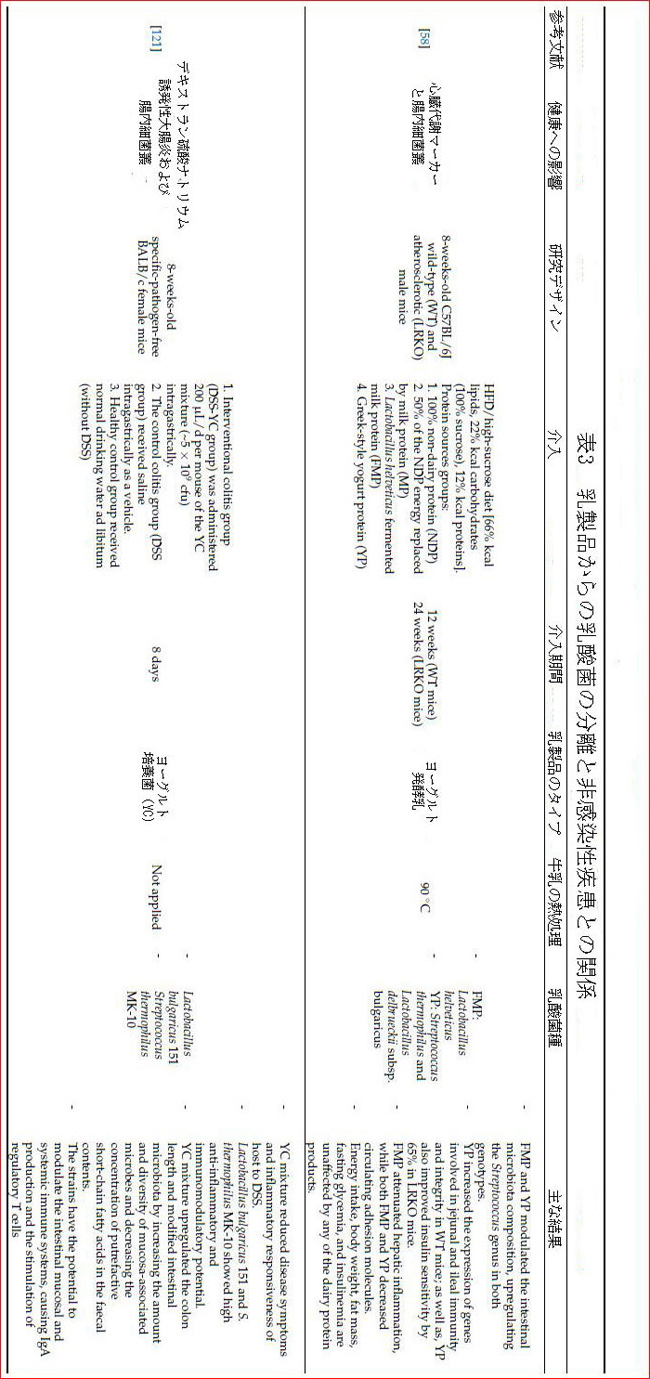
| ただし、場合によっては、アンジオテンシン変換酵素阻害ペプチドまたはそれらを構成する細菌を含む発酵製品の摂取がアンジオテンシン変換酵素阻害効果を示さないことがあります。これは、アンジオテンシン変換酵素阻害ペプチドが胃腸の消化中にさらに分解されるため、標的臓器で作用できないという事実によって説明されました[115]。さらに、発酵させるミルクに適用されるプロセスは、血清アルブミン、β-ラクトグロブリン、およびα-ラクトアルブミンの濃度を低下させ、タンパク質の構造に変化をもたらすことにより、同様の効果を引き起こす可能性があります。ミルクの過度の熱処理およびアルカリ処理は、難消化性ペプチド結合につながる加水分解酵素の作用に耐性のある分子間共有結合を形成することにより、アンジオテンシン変換酵素阻害剤であり抗酸化特性を有する生物活性ペプチドの形成を防ぐことができます[64]。滅菌ミルクと90°Cまで加熱されたミルクの使用は、健康を保護する効果がありました(表3)。 |
|

|
|

|

|
|
|
|
| 4.3 糖尿病 |
| 糖尿病は、深刻な健康、社会的、経済的影響を伴う世界的な大流行として定義されています。過去30年間で、2型糖尿病の有病率は2倍以上になりました[133]。2019年の世界の2型糖尿病の有病率は9.3%(4億6300万人)であり、この率は2030年までに10.2%(5億7800万人)、2045年までに10.9%(7億人)に増加すると推定されています[134]。2型糖尿病は成人でより一般的であり、すべての糖尿病症例の約90%を占めています。 糖尿病は、インスリン抵抗性または膵臓β細胞によるこのホルモンの不十分な分泌によって引き起こされる高血糖を特徴とする慢性疾患です[135]。世界のさまざまな地域での実験により、身体活動や健康的な栄養によるライフスタイルの変更が2型糖尿病の発症を遅らせたり妨げたりする可能性があることが明らかになりました[136]。乳製品の消費と糖尿病の発生率との関連を調べた研究は、一貫した結果はありませんが、逆の関連を示しています[137,138,139]。結果の不一致は、乳製品がそのサブグループ(脂肪、無脂肪、発酵など)に従って分析されていないという事実によって説明されます。乳製品の中で、特に低脂肪および発酵乳製品は、さまざまな研究で糖尿病の発生率を低下させることが示されています[139,140]。 これらの効果は、プロバイオティクス特性を示し、生物活性ペプチドを形成する乳製品乳酸菌株の結果として達成され、それによって酸化ストレスを軽減し、腸内細菌叢を調節し、免疫調節および抗炎症効果を示します[141]。 |
| 脂肪組織から放出されるFFA(訳者注:遊離脂肪酸)、アディポカイン、およびさまざまな炎症誘発性サイトカインは、2型糖尿病の病態生理において重要な役割を果たしています。循環する遊離脂肪酸とその酸化生成物の増加は、セリン/スレオニンキナーゼカスケードを活性化し、インスリンシグナル伝達の障害を引き起こす可能性があります[142]。インスリンシグナル伝達におけるこの欠陥を補うために、膵臓β細胞からのより多くのインスリン分泌は、小胞体ストレス、タンパク質の誤った折り畳み、そして最終的にはβ細胞のアポトーシスをもたらす[142]。乳製品の乳酸菌株の1つであるLactobacillus acidophilusは、膵臓細胞への損傷の原因となる活性酸素代謝物とサイトカインの産生を阻害し、血清インスリンレベルに調節効果をもたらします[106]。高フルクトース食でプロバイオティクス細菌(Lactobacillus acidophilus, Lacticaseibacillus casei および Lactococcus lactis biovar diacetylactis)を含む発酵乳製品(ダヒ)を摂取しているラットの空腹時血糖、インスリン、糖化ヘモグロビン(HbA1c)、および循環遊離脂肪酸レベル8週間は、高フルクトース食のみを与えられたものと比較して有意に低く、通常の食餌を与えられたものと同様でした[143]。 |
| 腸内毒素症を引き起こすグラム陰性菌の増加と腸内バリア機能の低下の結果として、これらの細菌に関連するリポ多糖(LPS)の循環レベルが増加します。ただし、リポ多糖レベルの増加は、NF-kB経路を活性化し、炎症誘発性バイオマーカー(CRP、TNF-α、IL-1β、およびIL-6)の循環レベルを増加させ、2型糖尿病の患者ではインスリン抵抗性の病因に関与します[106,144,145]。小腸細菌異常増殖(SIBO)の高齢者がLactobacillus johnsoniiLa1含有ヨーグルト109CFUを4週間毎日300g摂取すると、血漿リポ多糖パターン認識受容体とエンドトキシンレベルの低下が検出されたことが報告されています[145]。乳製品乳酸菌株は、これらの炎症性サイトカインの産生を阻害することにより、膵臓細胞に保護効果を発揮することができます[146]。ある研究では、Lactiplantibacillus plantarum OLL2712(> 5×109細胞/ヨーグルト112g)を含むヨーグルトを摂取しているわずかに肥満の人では、空腹時血糖値と血清IL-6レベルが大幅に低下することがわかりました[147]。2型糖尿病の患者では、300 gのプロバイオティクスヨーグルト(Lactobacillus delbrueckii subspbulgaricusおよびStreptococcusthermophilus Bifidobacterium animalis subsp lactisBb12(DSM 10140)およびLactobacillus acidophilus La5株; 3.7×106cfu /g)を摂取すると、空腹時血糖、IL-6、高感度CRP値に有意な変化を引き起こすことはありませんがHbA1cおよびTNF-αレベルの低下を示しました [146]。 研究間のこれらの異なる結果は、ヨーグルトに追加された乳酸菌株間の違いに起因する可能性があります。ただし、乳酸菌株を含むプロバイオティクスヨーグルトには、抗炎症作用と抗糖尿病作用がありますが、炎症誘発性マーカーは異なると結論付けることができます。 |
| 2型糖尿病またはインスリン抵抗性の場合、脂肪および筋肉組織へのインスリン依存性グルコース取り込みの失敗は、血糖濃度の増加およびインスリン非依存性組織へのグルコース取り込みの増加をもたらす。これは、終末糖化産物(AGE)などの酸化剤生成物の増加につながり、マクロファージの活性化に伴い、炎症性サイトカインと酸化ストレスを増加させ、β細胞の損傷を引き起こします[106,146]。発酵した種類のミルクは、その組成に抗酸化ペプチドがあるため、食事の天然の抗酸化源の1つです。最も重要な生物活性ペプチドの1つであるαs-カゼイン由来の生物活性ペプチドは、フリーラジカル捕捉、酵素的および非酵素的脂質過酸化阻害効果を有すると言われています。さらに、ホエイプロテインに由来する抗酸化ペプチドは、その構造中のシステインリッチアミノ酸がグルタチオン合成を助けるため、細胞内抗酸化特性を持っていることが示唆されています[141]。ある研究では、高脂肪食を12週間与えたラットで、糖質ケフィアから分離されたLactobacillus mali APS1株(5×108–109)を摂取すると、腸内細菌叢の酪酸菌の数が増加しました。そしてグルカゴン様ペプチド1(GLP-1)の活性化により、SIRT-1(訳者注:サーチュイン1、寿命を制御する分子のひとつとして働いている可能性が高い)およびPGC-1α(訳者注:転写活性化補助因子、この物質が筋肉中に少ない人は糖尿病になる危険性が高い)が上方制御され、SREBP-1(訳者注:肝臓において脂肪酸、中性脂肪の合成を支配する転写因子)が下方制御されます。 |
| これらの結果、血糖値が調節され、HOMA-IR値(訳者注:空腹時の血糖値とインスリン値から算出され、インスリン抵抗性(インスリンが効きにくく糖尿病になりやすい状態)を評価します。日本人では1.6以下が正常で2.5以上の場合は、インスリンに対する抵抗性があると考えます)が低下し、脂質代謝が調節されることが観察されました。さらに、SIRT-1のアップレギュレーションは、肝臓でのNrf2(訳者注:転写因子、炎症を増悪させるサイトカイン※3であるインターロイキン6(IL-6)やインターロイキン1β(IL-1β)の遺伝子の発現を阻害することで、炎症を抑えている)の活性化と抗酸化酵素の産生を増加させることにより、肝臓の酸化ストレスも抑制しました[148]。別の研究では、ケフィアの介入が糖尿病患者のNrf2経路を介して抗酸化酵素を活性化することも観察されました[149]。ミルク発酵中の25の乳酸菌株の抗酸化活性の変化に関連する別の研究では、Leuconostoc mesenteroides subsp cremoris、Lactococcus lactis、Lactobacillus acidophilus、Lactobacillus jenseniiおよびLactobacillus helveticus菌株がラジカルスカベンジング活性と脂質過酸化を阻害します。 そしてこの活動は発酵プロセスの間に増加します。発酵プロセス中に出現した生物活性ペプチドは、Leuconostoc mesenteroides subsp cremoris, Lactobacillus jenseniiおよびLactobacillus acidophilus株の抗酸化活性に有効であることが観察されました。しかし、すべての生物活性ペプチドがこの効果を持っているわけではないことが確認されています。たとえば、タンパク質分解活性の高いLactobacillus helveticusは、中程度の抗酸化作用を発揮します。さらに、混合乳酸菌培養を使用した発酵乳製品は、単一の細菌株を使用したものよりも高いラジカル捕捉活性を示しました[150]。 |
| 食事中の炭水化物は、α-アミラーゼ、パンクレリパーゼ、およびα-グルコシダーゼ酵素によって小腸から加水分解され、血糖値を調節します。腸上皮にコロニーを形成する乳酸菌は、加水分解によって形成されたグルコースをエネルギー源として使用することにより、グルコース吸収を低下させ、血糖値の調節を助けることができると言われています[146]。さらに、膵臓アミラーゼとα-グルコシダーゼの阻害は、2型糖尿病の効果的な治療法である可能性があります[151]。さまざまなブランドのプレーンヨーグルトとフルーティーヨーグルトのα-アミラーゼ抑制効果を評価したところ、ブランドによってはプレーンヨーグルトのα-アミラーゼ抑制効果が最大68%になる可能性があることがわかりました[151]。さらに、Limosilactobacillus fermentumM2およびM7スターターカルチャーで発酵させたミルクは、α-アミラーゼ(それぞれ65.3%および63.5%)およびα-グルコシダーゼ(それぞれ11.3%および13.7%)酵素に対して抑制効果を示したと言われています。この効果の主な理由は、発酵プロセスにおけるホエーやカゼインなどの乳タンパク質からなるペプチドでした[82]。乳酸菌株のタンパク質分解活性によって形成される生理活性ペプチドは、さまざまなメカニズムで抗糖尿病効果を発揮することが示されています。例えば、 Lactococcus lactisのタンパク質分解活性の結果として、α-およびβ-カゼインからなる生物活性オリゴペプチドは、インスリンおよびソマトスタチンの放出を刺激し、栄養素の胃腸吸収を遅らせると述べられました[1]。別の研究では、ケフィアペプチドで治療されたマウスのHOMA-IR、血糖、および空腹時インスリンレベルが対照群と比較して有意に低下したことが観察されました[152]。したがって、ケフィアペプチド治療は耐糖能を改善し、インスリン抵抗性を低下させることができることが示唆されています[152]。 |
| アディポネクチンは脂肪細胞由来の血清タンパク質であり、インスリン感受性の増加に重要な役割を果たします[153]。アディポネクチンは、エネルギーバランスの維持、グルコース恒常性、脂質代謝などの多くの代謝プロセスを調節し、肥満の人では一般的にアディポネクチンレベルが低くなります[127]。高脂肪食とLimosilactobacillus fermentumTSIで発酵させたヨーグルトを与えた肥満ラットは、高脂肪食のみを与えたラットよりもアディポネクチンレベルが有意に高いことがわかりました[127]。 |
| 文献における乳製品消費と糖尿病の関係は、主に従来のヨーグルトと、 Lactobacillus acidophilus La5やBifidobacterium lactis Bb12 などの菌株を含むプロバイオティクスヨーグルトを比較しました[106,141,146]。これらの研究の結果、プロバイオティクスヨーグルトは、血糖値、HbA1c、TNF-α、抗酸化能などの糖尿病関連パラメーターに対して有意に効果的であることが観察されました[141,146]。しかし、乳酸菌株(Lactobacillus bulgaricus と Streptococcus thermophilus)のみを含む従来のヨーグルトは、糖尿病関連のパラメーターの改善を示しませんでした[106,141,146]。 |
| 4.4 癌(がん) |
| 癌は、遺伝的および後成的突然変異に起因する悪性細胞の増殖として定義されます。 WHOのデータによると、癌は世界で2番目に多い死因です[154]。ほとんどの種類の癌は、時間の経過とともに細胞に蓄積する遺伝的変化または損傷によって引き起こされます。この損傷は、遺伝性突然変異、ホルモン特性、免疫状態などの個々の特性、および喫煙、感染性微生物、化学物質、食事成分などの環境要因によって引き起こされる可能性があります[155]。観察研究によると、癌の症例の約30〜40%は、栄養関連の要因を調節することで予防できることが示されています[156]。乳製品には多種多様な食品が含まれているため、乳製品の消費と癌のリスクとの間に一貫した関連性を得ることができませんが、発酵乳製品とその生理活性化合物に含まれる乳酸菌と、ミルクの組成に含まれるカルシウムは、癌のリスクを減らすのに効果的であることが示唆されています[157,158]。たとえば、発酵乳製品に含まれる乳酸菌やビフィズス菌などのプロバイオティクスは、グルクロニダーゼ、β-グルクロニダーゼ、アゾルトルターゼ、ニトロレダクターゼなどの糞便酵素の活性を変化させ、免疫細胞活性を高めることにより、結腸癌のリスクを軽減します[159]。β-グルクロニダーゼは、腸管腔内のグルクロニドの加水分解に関与する酵素です。この反応は、肝臓でのグルクロニドの形成によって無毒化され、胆汁を介して腸に入る有毒で発癌性の物質を生成します[159]。アグリコン構造の有毒な化合物は、細菌のβ-グルクロニダーゼによって腸内で生成される可能性があります。ヒトでは、糞便中のβ-グルクロニダーゼ活性は、健康な対照よりも結腸直腸癌患者の方が高く、この酵素が発癌に関与していることを示唆しています[159]。結腸癌の予防に重要な別の酵素はニトロレダクターゼです。 この酵素は、ニトロ化合物の芳香族アミンへの還元を担っています。反応性ニトロソ、N-ヒドロキシ中間体、芳香族アミンなど、これらの反応から得られる反応性の高い中間体および最終生成物は、変異原性および発癌性の影響を示します。芳香族ニトロ化合物とアゾ化合物の還元は、腸内細菌叢の活動によって達成できます[159]。 |
| 乳製品に含まれるプロバイオティクス乳酸菌は、腸内細菌叢に何らかの変化を引き起こすことにより、腸の発癌を防ぐことができます。プロバイオティクス乳酸菌は、腸内細菌叢で発癌性物質を発癌性物質に変換する細菌の増殖を抑制し、腸内の発癌性物質の数を減らすことによって、これらの効果を示します[160]。Lactobacillus kefiriの効果は、いくつかの種類の癌(胃癌細胞、乳癌細胞、およびヒト末梢血単核細胞)で調査されました。この乳酸菌株は胃癌細胞にアポトーシスを誘導し、アポトーシスはミトコンドリア膜電位の分極の減少とBcl2(訳者注:カスパーゼ活性阻害でアポトーシスを抑制する遺伝子)発現の減少に関連していることが示されています。しかし、他の癌細胞では、Lactobacillus kefiriはアポトーシスを誘導していません。これに関連して、この菌株は胃癌の治療における潜在的な薬剤である可能性があることが強調されました[161]。 |
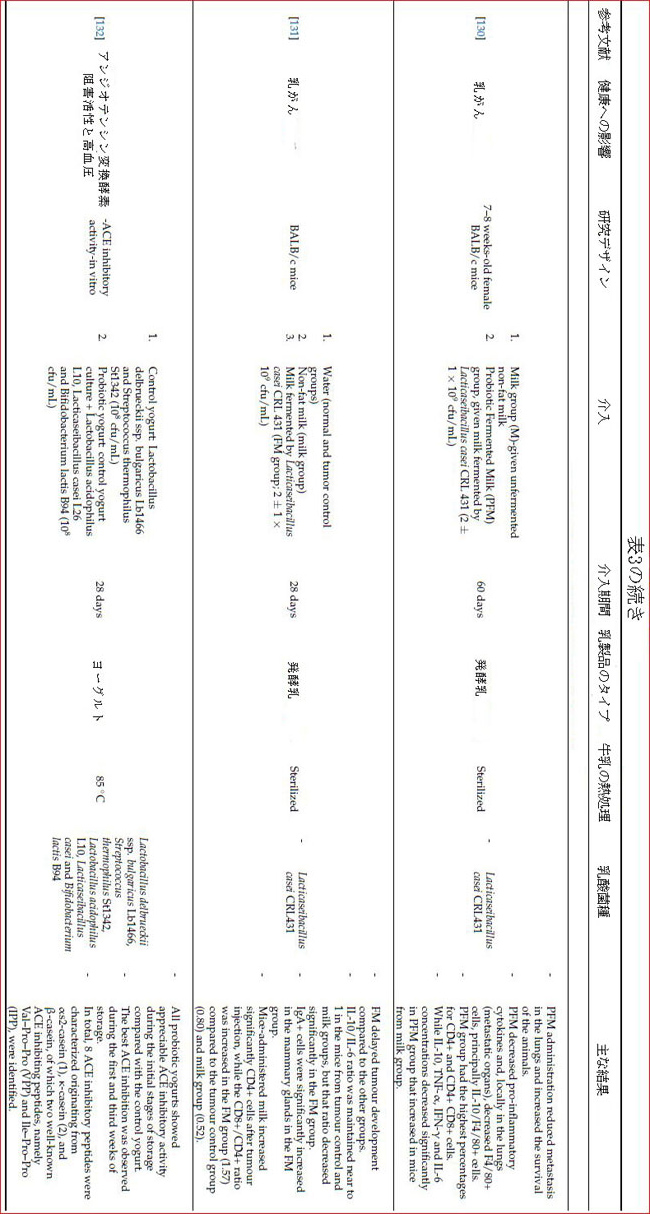
| 研究によると、乳製品の乳酸菌は、消化器系の癌だけでなく乳癌にも有益な効果があることが示されています[128]。最近の研究によると、Lacticaseibacillus caseiの菌株で発酵させたミルクの投与は、マウスモデルの乳癌腫瘍に対する免疫応答を調節しました。 [131]。 Lactobacillus helveticusR389で発酵させたミルクは、乳房腫瘍の成長を遅らせることが報告されました。この発酵乳は、血清、乳腺、免疫細胞のIL-6を減少させながら、IL-10とIL-4を増加させることがわかっています[160]。高いIL-6全身レベルは、乳癌患者の予後不良と相関しており、腫瘍量のバイオマーカーとなる可能性があります[130]。 Lacticaseibacillus casei CRL431株を含む発酵乳がBALB /cマウスの乳癌に及ぼす影響に関する研究では、発酵乳は腫瘍の発生を遅らせ、炎症誘発性マーカーを減らし、免疫応答を改善し、細胞レベルでいくつかの正の変化をもたらします(表3)[128,129]。 |
| 癌患者は、化学療法や放射線療法など、生活の質を低下させ、免疫系を損なう可能性のある治療を受けることがよくあります。 乳製品乳酸菌を用いた研究は有望であるように思われますが、これらの製品に使用されるプロバイオティクスの有効性と安全性は、特定の条件下で評価する必要があります[129]。しかし、癌に対する生物療法としてのプロバイオティクスの適用がテストされている十分な人間の研究はまだありません[131]。 |
| 4.5 炎症性腸疾患(IBD) |
| 炎症性腸疾患は、消化管の非感染性慢性炎症性疾患です。彼らの臨床的特徴によると、それは2つの慢性特発性疾患、潰瘍性大腸炎(UC)とクローン病(CD)を含みます[162]。炎症性腸疾患は世界的な病気であり、その有病率は世界中で増加していますが、その病理はまだ理解されていません。最近のデータは、遺伝的要因、腸内細菌叢、および環境要因がこの病気の病因に役割を果たすことを示唆しています。ヨーロッパと北アメリカは、炎症性腸疾患の有病率が最も高い地域として報告されています[163]。近年、炎症性腸疾患の発生率は工業化を始めたばかりの国で増加しています[164]。臨床試験は限られていますが、発酵食品は炎症性腸疾患の症状にプラスの影響を与える可能性があると報告されています。特に乳酸菌を含む発酵食品は、腸内細菌叢を積極的に変化させることにより、炎症性腸疾患にプラスの影響を与える可能性があると考えられています[165,166]。これに関連して、Saez-Lara etら(2014)は、乳製品に含まれ、プロバイオティクスとして発現する乳酸菌およびビフィズス菌が、腸内細菌叢を変化させることにより、炎症性腸疾患の予防と治療に有益な効果をもたらす可能性があると報告しました[166]。 |
| 炎症過程、特にTh1細胞とTh17細胞は、炎症性腸疾患の病態生理に関与していることが知られています。ある研究では、Th1 / Th17応答に対する発酵乳の影響が、軽度の炎症性腸疾患のマウスモデルで評価されました。 Limosilactobacillus fermentumで発酵させたミルクは、6週目に、それに含まれる代謝物または細胞成分(特に乳酸菌および細胞成分)を介して炎症反応を軽減することが示されています[167]。プロバイオティクス乳酸菌は、IL-1β、IL-6、IL-17、TNF-α、IFN-γなどの炎症誘発性サイトカインをダウンレギュレートすることにより、細胞レベルでいくつかの可能な経路を介して抗炎症効果を発揮する可能性があります[168,169]。さらに、生物活性ペプチドは、NF-kB経路を介して単球細胞におけるリポ多糖誘導性サイトカイン産生をダウンレギュレートすることにより抗炎症活性を提供することができます[170]。炎症性腸疾患患者におけるプロバイオティクスヨーグルトの抗炎症効果を観察するために、参加者にLactocaseibacillus rhamnosus GR-1とLimosilactobacillus reuteri RC-14を含むヨーグルトを30日間投与しました。プロバイオティクスヨーグルトの摂取は、炎症性腸疾患患者の有意な抗炎症転帰と、推定される制御性T(Treg)細胞の末梢プールの拡大と関連しています[165]。 |
| ケフィアとケフィア画分には抗炎症作用と抗酸化作用があり、ケフィアはラットのデキストラン硫酸ナトリウム(DSS)誘発性大腸炎に治癒効果があることが示されています[171]。Sevencan ら(2019)は、毎日10mLのケフィアが、トリニトロベンゼンスルホン酸によって引き起こされた大腸炎のラットの臨床症状と結腸の肉眼的損傷を軽減することを報告しました。それでも、同じ研究で、30mLのケフィアの1日量が大腸炎を悪化させることが示されています。 したがって、大腸炎誘発症状を改善するためのケフィア(Lactococcus lactis subsp., Leuconostoc subsp., Streptococcus thermophilus, Lactobacillus subsp.,およびケフィア酵母)の効果は用量依存的です[172]。 ケフィアから分離されたLactobacillus kefiri CIDCA8348株が、活動性炎症性腸疾患患者の腸T細胞に及ぼす免疫調節効果を調べると、Lactobacillus kefiriの存在、粘膜固有層T細胞の増殖率は低く、TNF-α、IL -6の分泌、IFN-γ、およびIL-13が減少しました。 さらに、Lactobacillus kefiriは、IL-10レベルが高いCD4 + FOXP3 (訳者注:制御性T細胞のマスター転写因子)+ 粘膜固有層 T細胞(LPTC)の頻度の増加を誘導しました。これらの発見は、Lactobacillus kefiri CIDCA8348菌株が炎症性腸疾患患者の腸細胞に免疫調節作用を及ぼす可能性があることを示唆しています[173]。 |
| 5 結論 |
| 乳製品、特にそれらの発酵製品は、乳製品乳酸菌の貴重な供給源です。これらの微生物は、発酵中の食品マトリックスへの生物活性化合物の生成を含むさまざまなレベルで作用することにより、腸内環境における生細胞と共生微生物叢との相互作用;複雑な生理学的コミュニケーションを仲介することができるシグナル伝達分子の放出および/または誘導など、宿主に対する有益な応答に関与しています。ただし、乳製品乳酸菌のプロバイオティクス効果は、食品-腸-脳軸の観点から、発酵乳製品の潜在的な健康上の利点とほとんど混同される可能性があります。それらが発酵のためのプロバイオティクスまたはスターターカルチャーであり、製品の機能性を高めるための新しい食品添加物であるかどうかにかかわらず、それらはあらゆる点で健康にプラスの健康調節効果をもたらします。 |
| 近年、プロバイオティクスおよび生物活性代謝物の供給源としての乳製品乳酸菌消費の有益な効果が、抗菌性および免疫調節性の可能性に特に注意を払って広く研究されています。ただし、非感染性疾患への貢献に関する最近の証拠は、さらなる解明が必要な新しい治療および予防の展望を開きます。それでも、文献には多くの観察データがありますが、特に食事療法の用途では、使用の可能性(用量、期間、種類、安定性など)の観点から深く明らかにする必要がある多くの問題があることも明らかです。 特に、腸内細菌叢の組成に対する食事スタイルの影響を明らかにするために、人間の臨床試験によるプロバイオティクスサプリメントの根底にある生化学的メカニズムや腸内細菌叢の長期コロニー形成を誘発するためのプロバイオティクス摂取の影響など、将来の調査に取り組む必要があります。 |
| 乳製品乳酸菌は、非感染性疾患の治療における微生物叢を標的とした潜在的な選択肢である可能性がありますが、特定の菌株を特定し、臨床試験で有益な結果をもたらす可能性のあるメカニズムを解明するには、さらなる研究が必要です。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
|
|
| この文献は、Foods 2021, 10, 3099に掲載されたDairy Lactic Acid Bacteria and Their Potential Function in Dietetics: The Food–Gut-Health Axisを日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |