Hyun-Ji Bock et al., |
要約 |
| 不眠症は最も一般的な睡眠障害の一つです。従来、その病態生理は主に中枢神経系(CNS)の観点から解釈されてきました。しかし、腸と中枢神経系を双方向的に繋ぐコミュニケーションネットワークである腸内細菌叢-腸-脳軸(MGBA)が、不眠症の発症、維持、治療において重要な役割を果たしている可能性を示唆する証拠が蓄積されています。本レビューでは、腸内細菌叢-腸-脳軸の主要なシグナル伝達経路を概説し、不眠症におけるその潜在的なメカニズムについて考察します。現在のエビデンスは、腸内細菌叢とその代謝産物が、神経系、免疫系、内分泌系、概日リズム経路を介して睡眠・覚醒恒常性に影響を与える可能性を示しています。同時に、不眠症に関連するストレス反応、免疫系の不均衡、生活習慣の乱れが腸内細菌叢に影響を与え、双方向的な調節ネットワークを形成する可能性があります。動物実験および臨床研究は、腸内細菌叢の異常と不眠症との密接な関連性をさらに裏付けています。さらに、本レビューでは、食事、生活習慣、心理社会的ストレス、薬剤、医療曝露など、腸内細菌叢-腸-脳軸に影響を与える可能性のある要因を体系的にまとめています。このことから、食事療法、プレバイオティクスやプロバイオティクス、生活習慣の改善、糞便微生物叢移植、天然薬など、腸内細菌叢-腸-脳軸を標的とした介入は、不眠症の予防と治療のための有望な新たな戦略となる可能性があります。しかしながら、現在のエビデンスは依然として動物実験や横断研究に大きく依存しており、因果関係、長期的な有効性、標準化された治療アプローチを明らかにするためには、さらなる縦断研究や質の高い介入試験が必要です。 |
| 目次(クリックして記事にアクセスできます) |
| 1. はじめに |
| 2. 腸内細菌叢-腸-脳軸とその双方向シグナル伝達経路 |
| 2.1. 腸内細菌叢の構成と機能 |
| 2.2. 神経経路 |
| 2.2.1. 迷走神経経路 |
| 2.2.2. 腸管神経系 |
| 2.2.3. 脊髄求心路 |
| 2.3. 内分泌経路 |
| 2.4. 免疫炎症経路 |
| 3. 腸内細菌叢-腸-脳軸と不眠症のメカニズム的関連性 |
| 3.1. 腸内細菌叢-腸-脳軸と神経伝達物質の不均衡 |
| 3.2. 腸内細菌叢-腸-脳軸および視床下部-下垂体-副腎軸の機能障害 |
| 3.3. 腸内細菌叢-腸-脳軸と免疫炎症異常 |
| 3.4. 腸内細菌叢-腸-脳軸と概日リズム障害 |
| 3.5. 腸内細菌叢と不眠症の関連性を裏付ける実験的および臨床的証拠 |
| 3.5.1. 動物モデルからの証拠:因果関係 |
| 3.5.2. 臨床観察によるエビデンス:関連性 |
| 4. 不眠症における腸内細菌叢-腸-脳軸に影響を与える因子 |
| 4.1. 食事とライフスタイル |
| 4.2. ストレスと心理的要因 |
| 4.3. 薬剤および医療関連因子 |
| 5. 腸内細菌叢-腸-脳軸を標的とした不眠症治療戦略 |
| 5.1. 食事療法 |
| 5.2. プレバイオティクスとプロバイオティクス |
| 5.3. 生活習慣介入 |
| 5.4. 糞便微生物移植 |
| 5.5. 天然薬 |
| 6. 結論と今後の展望 |
| 本文 |
| 1.はじめに |
| 都市化の加速、心理的ストレスの増加、ライフスタイルの変化に伴い、睡眠障害は公衆衛生上の大きな懸念事項となっています[1]。最近の文献の系統的レビューに基づく世界的な分析では、世界中の成人の約16.2%、すなわち約8億5200万人が不眠症に苦しんでいると推定されています[2]。女性[2]、高齢者[3]、およびストレス反応性の高い人[4]は特に影響を受けやすいようです。しかしながら、不眠症の有病率は、使用される診断基準と評価方法によって大きく左右され、研究や地域によって著しいばらつきが生じることに留意する必要があります[5]。例えば、同じ集団における有病率の推定値は、適用される診断基準によって大きく異なり、精神疾患の診断・統計マニュアル第5版(DSM-5)基準では8.5%、国際睡眠障害分類第3版(ICSD-3)基準では20.0%となっています[6]。 |
| 従来の神経生物学的枠組みでは、過覚醒は不眠症の根底にある主要なメカニズムとして広く認識されている[7]。この文脈では、神経伝達物質の不均衡[8]、ストレス関連神経内分泌系の活性化亢進[7]、免疫炎症活性化[9]、概日リズムの乱れ[10]、認知情動調節不全[11]など、複数のプロセスが不眠症の発症と進行に寄与し、過覚醒状態を共同で促進または維持する可能性がある。慢性不眠症は睡眠の質と日中の機能を損なうだけでなく、さまざまな健康上の悪影響とも密接に関連している。不眠症が不安やうつ病などの精神疾患と双方向的に相互作用する可能性があることを示す証拠が増えている[12,13]。さらに、不眠症は代謝疾患や心血管疾患と有意に関連している[14,15]。これらの研究結果は、中枢神経系のみに焦点を当てた従来の仮説では、不眠症の多系統的かつ多次元的な性質を十分に説明できないことを示唆している。このことから、末梢調節系、特に腸内細菌叢(GM)と中枢神経系との双方向コミュニケーションネットワークへの関心が高まっている。 |
| 腸内細菌叢-腸-脳軸は、主に神経、内分泌、免疫経路を介して消化管と中枢神経系を繋ぐ複雑な双方向コミュニケーションシステムである[16]。この軸の重要な構成要素として、腸内細菌叢は宿主の生理的恒常性を維持し、複数のメカニズムを介して脳機能を調節する上で不可欠な役割を果たしている。例えば、腸内細菌叢は短鎖脂肪酸などの代謝産物を生成し、これが神経伝達物質の合成と調節に関与している。さらに、腸内細菌叢は迷走神経求心性シグナル伝達(訳者注:迷走神経求心性シグナル伝達とは、内臓で生じた感覚や生理状態の情報を、末梢から中枢神経系(脳幹の孤束核など)へと伝達する神経回路網です。内受容感覚(内臓感覚)を脳へ伝える主要な経路であり、食欲調節、情動、自律神経反射などの全身恒常性(ホメオスタシス)の維持に不可欠な役割を果たします)を活性化し、免疫炎症反応や神経内分泌反応を調節することで中枢神経系の機能に影響を与える可能性がある[16,17]。 |
| 近年、蓄積された証拠から、健康な人と比較して、不眠症患者は腸内微生物の構成と多様性に著しい変化を示すことが明らかになっている。これらの変化には、アルファ多様性の低下、ベータ多様性の変化、フェカリバクテリウムなどの有益な分類群の相対存在量の減少、および放線菌やバクテロイデスのメンバーを含む日和見的または潜在的に病原性のある分類群の存在量の増加が含まれる可能性がある[18]。このようなディスバイオシスは、神経伝達物質の恒常性に影響を与え、炎症シグナル伝達カスケードを促進し、視床下部-下垂体-副腎(HPA)軸の反応性を高めることにより、睡眠調節を阻害する可能性がある[16,17]。 |
| 本レビューでは、腸内細菌叢-腸-脳軸の主要なシグナル伝達経路、不眠症との潜在的なメカニズム的関連性、および入手可能な臨床的・実験的証拠を概説する。さらに、腸内細菌叢-腸-脳軸を介して睡眠に影響を与える可能性のある因子について議論し、不眠症の病態生理を理解し、潜在的な治療機会を特定するためのより包括的な枠組みを提供することを目的として、新たに登場する微生物叢を標的とした介入戦略の概要を示す。 |
| 2. 腸内細菌叢-腸-脳軸とその双方向シグナル伝達経路 |
| 2.1. 腸内細菌叢の構成と機能 |
| 腸内細菌叢は、ヒトの消化管に生息する複雑な微生物群集であり、細菌、真菌、ウイルス、古細菌などが含まれる[19]。腸内細菌叢は、食物の消化と栄養素の変換に関与するだけでなく、腸上皮細胞、粘液分泌、タイトジャンクションタンパク質の発現を調節することで、腸管バリアの完全性を維持する役割も担っている[20]。さらに、宿主免疫系との相互作用を通じて、腸内細菌叢は免疫寛容の確立と炎症反応の調節に貢献し、免疫恒常性と代謝バランスの維持に不可欠な役割を果たしている[21,22]。 |
| 一般的に、微生物の多様性が高く、コミュニティ構造がより安定していることは、腸内生態系がより健康であることを反映していると考えられているが[23]、ディスバイオシスは、幅広い疾患の発症と進行に関与している。例えば、肥満や糖尿病の患者は、短鎖脂肪酸産生菌の減少や炎症誘発性分類群の増加など、エネルギー代謝に関連する微生物叢の変化を示すことが多い[24,25]。システマティックレビューやメタアナリシスでは、リウマチ性疾患や免疫関連疾患の患者は、免疫調節異常や自己抗原への曝露増加に関連する顕著な腸内微生物の障害を示すことも示されている[26]。神経精神医学の分野では、うつ病や不安障害の患者は、微生物の多様性の低下や特定の分類群の存在量の変化など、腸内微生物組成の異常を示すことが多い[27]。 腸内細菌叢は末梢代謝および免疫恒常性の調節因子であるだけでなく、神経、免疫、および内分泌経路を介して中枢神経系機能の調節にも関与している可能性があることを示唆する証拠が増えている[28]。 |
| 2.2. 神経経路 |
| 神経シグナル伝達は、腸内細菌叢-腸-脳軸(腸管・脳・消化管)内における最も速く直接的な情報伝達経路の一つです。この双方向の情報伝達は主に自律神経系(ANS)によって媒介され、腸と中枢神経系間の継続的な情報交換を可能にしています。腸管で発生した感覚信号は迷走神経(訳者注:迷走神経(めいそうしんけい)は、脳から内臓へ伸びる最も長い脳神経(第10脳神経)です。心拍や消化、呼吸などの生命維持を司る「副交感神経」の大部分を構成し、脳と全身の器官を結ぶ重要な役割を果たしています)および脊髄求心路(訳者注:脊髄求心路(せきずいきゅうしんろ)とは、体の各部位(皮膚、筋肉、内臓など)で受け取った感覚刺激を、末梢から中枢(脊髄や脳)へと伝える神経の通り道のことです)を介して中枢神経系に伝達され、一方、中枢神経系は交感神経(訳者注:交感神経(こうかんしんけい)とは、自律神経系の一部で、体が活動しているときや強いストレスを感じたときに活発になる神経です。自動車でいう「アクセル」の役割を果たし、心身を臨戦態勢(戦闘・活動モード)に整えます)および副交感神経(訳者注:副交感神経は、心身をリラックスさせ、休息や回復を促す自律神経です。心拍数を下げて血管を広げ、胃腸の消化吸収を活性化させる「ブレーキ」の役割を果たします。夜間や休息時に優位に働きます)の遠心性出力を介して腸管運動、分泌、およびバリア機能を調節します[29]。これらの神経回路は、腸-脳コミュニケーションの構造的基盤を提供し、微生物由来の代謝シグナルが神経調節ネットワークと統合されることで、脳機能と行動に影響を与えることを可能にします。 |
| 2.2.1. 迷走神経経路 |
| 迷走神経は、腸管神経系(ENS)と中枢神経系を結ぶ主要な神経経路であり、腸-脳コミュニケーションにおいて重要な役割を果たしています。腸管腔内の機械的刺激、栄養素、化学信号、および微生物代謝産物は、腸管神経系および腸内分泌細胞(EEC)によって感知・統合され、その後、迷走神経求心路を介して脳幹の孤束核(NTS)(訳者注:孤束核(こそくかく)は、脳幹の延髄にある神経核で、味覚情報や心臓、肺、胃腸などの内臓からの感覚情報を中継する極めて重要な器官です。全身の臓器からの信号を統合し、自律神経の調節や情動(こころの制御)に関わっています)に伝達されます[30,31]。迷走神経を介した腸-脳シグナル伝達は、辺縁系および報酬関連脳領域の神経活動にも影響を与える可能性があります[32]。さらに、この経路は海馬関連の記憶プロセスや神経可塑性を調節し、特定の認知機能に影響を与える可能性があります[33]。 |
| さまざまな微生物代謝産物は、受容体を介したメカニズムによって迷走神経活動に影響を与える可能性があります。たとえば、短鎖脂肪酸は、遊離脂肪酸受容体3(FFAR3)を含む経路を介して迷走神経求心性ニューロンの活動を調節する可能性があります[34]。胆汁酸もまた、Gタンパク質共役胆汁酸受容体5(TGR5)を介したメカニズムによって迷走神経シグナル伝達を調節し、それによって腸から脳幹へのシグナル伝達を強化する可能性があります[35]。さらに、腸の腸クロム親和性細胞は大量の末梢5-ヒドロキシトリプタミン(5-HT)(訳者注:5-HTは「5-ヒドロキシトリプトファン」の略称であり、神経伝達物質のセロトニンを指す医学・生理学的な用語です。脳内では精神の安定や睡眠、体温調節に関わり、腸管では消化管の運動や血管収縮などに重要な役割を果たしています。5-HTは以下セロトニンと記載します。)を合成し、これは迷走神経求心性線維に発現する特定のセロトニン受容体サブタイプに作用し、それによって中枢モノアミン作動性システムを調節し、情動およびストレス関連の反応に影響を与える可能性があります[36]。 |
| 2.2.2. 腸管神経系 |
| 腸管神経系はしばしば「第二の脳」と呼ばれ、末梢神経系の中で最大の区分を占めています。多数のニューロンとグリア細胞から構成され、中枢神経系と構造的および機能的に多くの類似点を共有しています。腸管神経系は中枢神経系と同様の神経伝達物質、受容体、転写因子を多数発現し、複雑な局所神経回路を形成しています[37]。 |
| まず、腸管神経系は腸内微小環境の変化を直接感知し、局所反射回路を介して腸の運動、分泌、血流などの複数の生理活動を調節することができます[38,39]。同時に、腸内細菌、腸管上皮細胞、免疫細胞からの信号を統合し、神経伝達物質の放出と局所神経活動を調節することで、腸内恒常性に貢献しています[40]。これまでの研究では、腸管神経系の機能不全は、アルツハイマー病、パーキンソン病、自閉症スペクトラム障害など、さまざまな神経疾患や神経発達障害と関連していることが示されています[37]。これらの知見は、腸管神経系の構造的および機能的変化が、腸由来の信号が中枢神経回路に伝達されるのに影響を与える可能性があることを示唆しています。 |
| 2.2.3. 脊髄求心路 |
| 脊髄求心性神経は、腸管由来の感覚情報を中枢神経系に伝達するもう一つの重要な経路です。これらのニューロンの細胞体は後根神経節(DRG)に位置し、末梢終末は腸壁内に分布しており、機械的伸展、化学伝達物質、炎症、有害刺激など、さまざまな信号を感知します[41]。これらの信号はまず脊髄後角の表層で処理され、局所的な興奮性回路と抑制性回路の複合的な調節の下で、内臓感覚入力および侵害受容性入力と統合されます[42]。その後、関連する信号は脊髄視床路や脊髄網様体路などの上行路を介して脳幹、視床、辺縁系に伝達され、中枢感覚調節に関与します[42]。さらに、中脳水道周囲灰白質および吻側腹内側延髄から始まる下行路は、脊髄後角の興奮性を調節し、双方向の調節ネットワークを形成する[43]。 |
| 主に恒常性および代謝シグナルを伝達する迷走神経とは異なり、脊髄求心性神経は炎症性メディエーターおよび組織損傷に特に敏感である。炎症誘発性サイトカイン、細菌エンドトキシン、および腸内微生物代謝産物は、後根神経節ニューロンの興奮性を変化させ、それによって中枢神経処理および神経免疫相互作用に影響を与える可能性がある[44]。 |
| 2.3. 内分泌経路 |
| 内分泌シグナル伝達は腸内細菌叢-腸-脳軸のもう一つの主要な構成要素であり、主に循環ホルモンと神経活性ペプチドを介して腸脳コミュニケーションを媒介します。腸上皮全体に分布する腸内分泌細胞(EEC)は、腸内細菌叢-腸-脳軸の重要なゲートキーパーとしてますます認識されています[45]。これらの細胞は、腸管腔内の栄養素や微生物代謝産物を感知し、セロトニン、コレシストキニン(CCK)、グルカゴン様ペプチド-1(GLP-1)、ペプチドYY(PYY)など、さまざまな生理活性分子を放出します。これらのシグナル伝達分子は、体液性経路または神経経路を介して中枢調節回路にさらに影響を与え[31]、腸由来のシグナルを宿主の代謝および神経調節プロセスと結びつけます。 |
| これらのメディエーターの中で、セロトニンは神経伝達物質とホルモンの両方として機能します。体内のセロトニンの約90%は、腸クロム親和性細胞によって食事由来のトリプトファンから合成されます。末梢のセロトニンは、迷走神経求心性シグナル伝達、免疫調節、および代謝経路を介して中枢神経ネットワークを間接的に調節する可能性があります[46]。セロトニンに加えて、腸内分泌細胞から分泌されるペプチドホルモンも腸内細菌叢-腸-脳軸で重要な役割を果たします。グルカゴン様ペプチド-1、コレシストキニン、およびペプチドYYは、脳幹および視床下部の受容体に作用し、摂食行動、情動状態、およびエネルギー代謝の調節に関与します[31,47,48]。これらのペプチドの中で、グルカゴン様ペプチド-1は近年特に注目を集めています。研究により、グルカゴン様ペプチド-1はPI3K/Akt(訳者注:PI3K(ホスファチジルイノシトール3-キナーゼ)およびAKTは、細胞の増殖、生存、代謝を制御する最も重要な細胞内シグナル伝達経路(PI3K/AKT経路)の中心的な構成要素です。成長因子などの刺激により活性化され、この機能の異常はがんや糖尿病などの疾患に関与しています)などの細胞内シグナル伝達経路を活性化し、神経細胞の生存を促進し、酸化ストレスと神経炎症を軽減できることが示されています。さらに、グルカゴン様ペプチド-1受容体作動薬は、いくつかの神経変性疾患モデルにおいて神経機能障害を改善し、炎症性損傷を軽減することが示されている[47,49]。 |
| 視床下部-下垂体-副腎軸は、腸と脳をつなぐもう1つの重要な神経内分泌経路であり、ストレス反応における主要なエフェクターシステムとして機能します。腸内細菌叢とその代謝産物は、免疫炎症シグナル伝達と迷走神経求心性入力を介して視床下部-下垂体-副腎軸の活動を共同で調節し、それによって副腎皮質刺激ホルモン放出ホルモン-副腎皮質刺激ホルモン-コルチゾールカスケードに影響を与える可能性があります[50]。コルチゾールの持続的な上昇は、海馬、前頭前皮質、およびその他の感情関連の脳領域に構造的および機能的な変化を引き起こし、神経可塑性の低下と神経栄養因子の発現の低下につながる可能性があります。これらの変化は、精神疾患の発症と進行に密接に関連していると考えられています[51]。 |
| 同時に、視床下部-下垂体-副腎軸の活性化は中枢神経系に影響を与えるだけでなく、腸の生理機能にもフィードバックします。コルチゾールレベルの上昇は、腸管バリアの完全性を損ない、腸管透過性を高め、粘膜免疫活性化と炎症反応を促進し、それによって腸内細菌叢の異常と全身性の低度炎症を悪化させる可能性があります[52]。動物実験では、慢性的なグルココルチコイド曝露が不安様行動やうつ病様行動を誘発し、腸内微生物組成と脳代謝プロファイルの変化を伴うことがさらに示されています[53]。 |
| 2.4. 免疫炎症経路 |
| 腸管は体内で最も多くの免疫細胞を擁しており[54]、腸内細菌叢とその代謝産物の変化を感知することができる。腸管免疫系は、自然免疫応答と獲得免疫応答、炎症性メディエーターの放出、およびバリア機能を調節することにより、末梢および中枢の炎症状態に影響を与えることができる[55]。同時に、中枢神経系は神経機構および内分泌機構を介して腸管免疫応答を調節することができ、それによって腸管-免疫調節系(腸内細菌叢-腸-脳軸)内に双方向性の免疫炎症調節経路が形成される[56]。 |
| 腸内細菌叢の異常や腸管バリア機能の障害が発生すると、腸管免疫恒常性が乱れ、炎症性サイトカインやエンドトキシンのレベルが上昇し、慢性的な低度炎症が促進される可能性がある[57]。インターロイキン(IL)-1β、IL-6、腫瘍壊死因子(TNF)-αなどのこれらの炎症性メディエーターは、血液-脳関門(BBB)を破壊し[58]、ミクログリアを活性化し、サイトカイン受容体関連シグナル伝達経路を介して神経伝達物質恒常性やシナプスリモデリングを変化させ、最終的に中枢神経系機能に影響を与える可能性がある[59]。 |
| 体液性炎症シグナル伝達に加えて、腸管免疫細胞から放出されるサイトカインは、迷走神経求心性終末を直接活性化し、脳幹の孤束核にシグナルを伝達し、それによって高次神経回路に影響を与える可能性がある[60]。一方、中枢神経系は自律神経系を介して末梢免疫応答を調節することができる。重要なメカニズムの1つは、迷走神経シグナル伝達によって媒介されるコリン作動性抗炎症経路であり、アセチルコリンはマクロファージなどの免疫細胞の炎症促進応答を抑制する[61]。対照的に、慢性ストレス下での持続的な交感神経活性化は、炎症反応を増幅し、腸管上皮バリア機能を損なう傾向がある[62]。 |
| さらに、視床下部-下垂体-副腎軸は免疫炎症反応の重要な上流調節因子です。短期的には、ストレス誘発性グルココルチコイド上昇は一般的に免疫抑制作用と抗炎症作用を発揮します[63]。しかし、視床下部-下垂体-副腎軸の慢性的または反復的な過剰活性化は、腸管バリア、微生物組成、炎症環境に影響を与えることで、免疫恒常性を破壊し、腸管および中枢の炎症反応をさらに増幅させる可能性があります[62,63]。 |
| 総合すると、腸内細菌叢-腸-脳軸は神経、内分泌、免疫炎症経路の協調作用を通じて高度に相互接続され動的に調節される双方向のコミュニケーションネットワークを構成し、それによって腸と中枢神経系の間の恒常性の維持に貢献している(図1)。 |
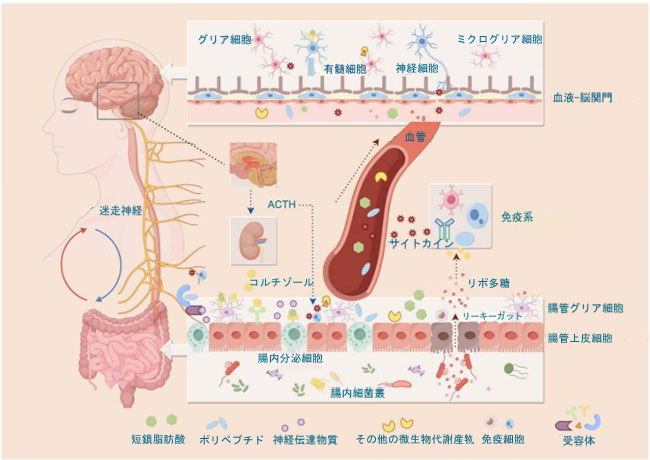 |
図1.腸内細菌叢-腸-脳軸の双方向シグナル伝達経路(赤い矢印は腸から脳へのシグナル伝達を示し、青い矢印は脳から腸へのシグナル伝達を示す)。 中枢神経系は、副腎皮質刺激ホルモン(ACTH)とコルチゾール(青い破線の矢印)の放出を含む迷走神経と視床下部-下垂体-副腎軸を介して腸機能を調節し、それによって腸の透過性、バリアの完全性、および免疫活性を調節する。逆に、腸由来のシグナルは、相互接続されたいくつかの経路を介して脳に到達する: (i)微生物代謝物(短鎖脂肪酸など)によって活性化される迷走神経と脊髄求心性神経、腸内分泌細胞(EEC)由来の神経伝達物質(5-ヒドロキシトリプタミンなど)、および免疫細胞から放出されるサイトカインを含む神経経路。 (ii)免疫炎症経路。腸管バリアの破壊によりリポ多糖(LPS)が免疫細胞を活性化し、サイトカイン産生を誘導する(右の赤い破線矢印)。さらに、免疫細胞とニューロン間の局所的なクロストークを介して媒介される。 (iii)神経内分泌経路。腸管ホルモンやその他の循環メディエーターが中枢シグナル伝達に影響を与える(中央の赤い破線矢印)。 |
| 3. 腸内細菌叢-腸-脳軸と不眠症のメカニズム的関連性 |
| 睡眠の開始と維持は複雑な生理学的プロセスです。従来の神経生物学的枠組みでは、不眠症は一般的に睡眠・覚醒調節系の機能不全に起因すると考えられています[64]。提唱されているメカニズムの中でも、過覚醒は不眠症の中核的な病態生理学的特徴の一つとされており、覚醒促進系と睡眠促進系のバランスの崩れを反映しています[7]。さらに、神経伝達物質の異常、免疫炎症の活性化、ストレス関連の神経内分泌系の機能不全、概日リズムの乱れなども、不眠症の発症と慢性化に寄与すると考えられています[8,64,65]。 |
| 近年、蓄積された証拠は、腸内細菌叢の異常と関連する腸内細菌叢-腸-脳軸の異常が、睡眠覚醒恒常性の重要な末梢調節因子としても機能する可能性があることを示唆している[18,66]。腸内細菌叢-腸-脳軸は、腸内細菌叢、腸管神経系、免疫系、中枢神経系を連結することで、多次元的かつ双方向的な調節ネットワークを形成する。腸内細菌叢-腸-脳軸は、神経伝達物質代謝、ストレス反応性、免疫炎症状態、概日リズムに対する潜在的な影響を通じて、不眠症の発症と進行に関与する可能性がある[66]。 |
| 3.1. 腸内細菌叢-腸-脳軸と神経伝達物質の不均衡 |
| 従来の考え方では主に中枢神経伝達物質に焦点を当てていましたが、腸内細菌叢-腸-脳軸は腸が栄養吸収器官であるだけでなく、複数の神経活性分子の生成、変換、統合の重要な場でもあることを強調しています。腸内細菌叢-腸-脳軸が不眠症に関連する神経伝達物質の不均衡に及ぼす影響は、主に2つの側面から考えられます。一つは、トリプトファン代謝、短鎖脂肪酸、その他の腸内細菌由来代謝産物を調節することで、中枢神経伝達物質の恒常性に影響を与える可能性です。もう一つは、腸由来の神経活性分子は血液-脳関門を容易に通過しないものの、迷走神経、免疫炎症シグナル伝達、神経内分泌調節を介した伝達経路によって、睡眠関連脳領域の機能を間接的に調節する可能性があることです。 |
| まず、腸内細菌叢-腸-脳軸は神経伝達物質の前駆体および関連する代謝経路に影響を与えることで、中枢神経伝達物質の恒常性に関与する可能性がある。トリプトファンはセロトニンの重要な前駆体であり、その腸内代謝は主にセロトニン、キヌレニン、およびインドール経路を介して、宿主と腸内細菌叢によって共同で調節されている[67]。ディスバイオシス状態では、トリプトファン代謝がキヌレニン経路にシフトし、セロトニンの利用可能性が低下し、睡眠調節が損なわれる可能性がある[67]。この経路の代謝産物のうち、キヌレン酸(KYNA)の上昇は、急速眼球運動(REM)睡眠の減少、ノンレム睡眠中のデルタ波パワーの低下、および睡眠紡錘波の減少と関連しており、キヌレン酸合成の阻害はこれらの異常を逆転させることが示されている[68]。インドール誘導体であるインドール-3-酢酸(IAA)も睡眠調節に関与する可能性がある。これまでの研究では、インドール-3-酢酸は腸クロム親和性細胞のアリール炭化水素受容体を活性化することでセロトニンの放出を促進し、睡眠不足に伴う認知機能障害を軽減する可能性があることが示されている[69]。 |
| さらに、微生物発酵によって生成されるチロシン関連シグナルは脳組織に侵入する可能性があり[70]、チロシン自体はカテコールアミン神経伝達物質の重要な前駆体である。睡眠関連神経伝達物質の中で、γ-アミノ酪酸は中枢神経系における主要な抑制性神経伝達物質である。γ-アミノ酪酸は睡眠の開始と維持に密接に関連しており、神経興奮性を低下させ、入眠潜時を短縮し、ノンレム睡眠を延長し、睡眠の質を改善する可能性がある[8,71]。 |
| 第二に、腸クロム親和性細胞、腸管神経系、および腸内細菌叢は、セロトニン、γ-アミノ酪酸、およびドーパミンなどの腸由来神経伝達物質の産生と調節に共同で関与しています。腸由来のセロトニンとγ-アミノ酪酸は、迷走神経求心性終末に存在する受容体に作用し、孤束核に信号を伝達し、さらに背側縫線核、青斑核、視床下部などの睡眠関連脳領域に影響を与え、それによって睡眠の安定性と覚醒レベルを調節する可能性があると提唱されています[36,72]。さらに、腸内細菌叢由来のドーパミン関連代謝物を含む末梢ドーパミン作動性信号は、迷走神経、免疫炎症、および視床下部-下垂体-副腎軸関連メカニズムを介して中枢ドーパミン利用可能性とシグナル伝達に影響を与え、それによって睡眠覚醒調節に寄与する可能性があります[73]。 |
| 3.2. 腸内細菌叢-腸-脳軸および視床下部-下垂体-副腎軸の機能障害 |
| 視床下部-下垂体-副腎軸の機能障害は、不眠症の主要な神経内分泌学的基盤の一つと考えられています。この軸の異常な活性化は、持続的なコルチゾール値の上昇と過覚醒状態を伴い、不眠症の発症と維持に寄与します[74]。不眠症の3Pモデルによれば、ストレスの多いライフイベント、心理的負担、環境的障害はしばしば誘発因子として作用し、睡眠反応性の高さなどの既存の特性は宿主の脆弱性を構成します。これらの因子が重なり合うと、不眠症が誘発され、維持される可能性が高くなります[4,75]。 |
| 最近の研究では、腸内細菌叢の振動が視床下部-下垂体-副腎皮質系(視床下部-下垂体-副腎軸)の日周リズムを調節している可能性が示唆されている。腸内細菌叢の枯渇は、海馬と扁桃体におけるストレス関連の転写および代謝プロファイルを阻害し、グルココルチコイドリズムの異常を引き起こすことが示されているが、糞便微生物叢移植はこれらの変化を部分的に回復させる可能性がある[76]。ストレス条件下では、グルココルチコイドレベルの上昇により腸管透過性と微生物組成が変化し、有益な細菌が減少し、潜在的に病原性のある分類群が増加する可能性がある。これらの変化は、腸-脳軸を介して睡眠および感情に関連する神経調節プロセスにさらに影響を与える可能性がある[52,77]。この文脈では、不眠症はストレス曝露と宿主感受性の相互作用から生じる可能性があり、腸内細菌叢異常が重要な媒介因子として機能している。 |
| 腸内細菌叢関連代謝物のうち、短鎖脂肪酸はエピジェネティック制御と神経内分泌経路を介して視床下部-下垂体-副腎軸の活動に影響を与え、それによってストレス反応を調節し、ストレス関連の過覚醒を軽減する可能性がある[78]。腸由来のγ-アミノ酪酸も視床下部-下垂体-副腎軸の活動を阻害することでストレスを軽減し、それによって感情や睡眠に関連する生理学的プロセスに寄与する可能性がある[79]。さらに、グルカゴン様ペプチド-1、コレシストキニン、ペプチドYYなどのホルモンは、体液性経路または迷走神経経路を介して視床下部シグナル伝達に影響を与え、エネルギー代謝と摂食行動の調節に関与し、睡眠覚醒状態に影響を与える可能性がある[80]。 |
| 重要なことに、不眠症とそれに伴う視床下部-下垂体-副腎軸の過活動は腸内細菌叢にもフィードバックし、悪循環を形成する可能性がある。動物実験では、睡眠不足は視床下部-下垂体-副腎軸の活性化とコルチコステロンレベルの上昇、乳酸菌の減少や短鎖脂肪酸などの有益な代謝物の枯渇といった腸内細菌叢の著しい変化を伴うことが示されている。これらの変化は睡眠障害をさらに悪化させる可能性がある[81]。 |
| 3.3. 腸内細菌叢-腸-脳軸と免疫炎症異常 |
| 免疫炎症調節異常は、不眠症の主要な病理学的基盤の一つと考えられており、腸内細菌叢の変化と中枢神経系機能との重要な関連性を示しています。睡眠と炎症の間には明確な双方向の関係があることを示す証拠が増えています。一方では、末梢および中枢の炎症プロセスが睡眠構造と睡眠恒常性を阻害する可能性があります[82]。他方では、持続的な不眠症自体が免疫バランスの崩れを引き起こし、炎症反応を増幅させ、悪循環を生み出す可能性があります[83]。特に、持続的な炎症活性化は、視床下部-下垂体-副腎軸および概日リズム調節系にも影響を与え、日中のコルチゾール分泌の乱れや生理的覚醒の亢進を伴う可能性があります[84]。 |
| 臨床研究はこの関連性を裏付けている。不眠症患者では、C反応性タンパク質(CRP)、IL-6、TNF-αなどの炎症マーカーのレベルが上昇していることが報告されている[83]。これらの炎症性メディエーターは、神経機能やシナプス可塑性に影響を与えるだけでなく、睡眠に関与する神経伝達物質系を阻害することで睡眠調節異常にも寄与する可能性がある[83]。特に、IL-1βとTNF-αはノンレム睡眠と徐波活動に密接に関連しており、睡眠欲求に影響を与える可能性がある[85]。さらに、アデノシン、ケモカイン、活性酸素種などの他の炎症関連シグナル伝達分子も、睡眠構造と睡眠恒常性に影響を与える可能性がある[86]。 |
| 腸内細菌叢-腸-脳軸内では、腸粘膜バリアの障害は末梢炎症の重要な開始イベントと考えられています。腸内細菌叢の異常は、腸上皮バリアの完全性を弱め、腸管透過性を高め、腸内細菌叢関連産物の循環への侵入を促進し、それによって末梢免疫応答と炎症性サイトカインの放出を引き起こします [20]。これらの炎症シグナルはさらに 血液-脳関門に作用し、その透過性を高め、末梢炎症シグナルの中枢神経系への伝達を促進する可能性があります [58]。その中でも、IL-1β と TNF-α は中枢睡眠調節ネットワークと関連する神経伝達物質系に直接影響を与え、それによって睡眠構造を変化させ、睡眠の質を低下させる可能性があります [86]。 |
| 腸内微生物代謝物もこのプロセスにおいて重要な免疫調節の役割を果たしている。主要な腸内細菌由来代謝物である短鎖脂肪酸は、Gタンパク質共役受容体シグナル伝達とヒストン脱アセチル化酵素(HDAC)の阻害を介して免疫恒常性の維持に役立ち、それによって中枢睡眠調節回路に対する炎症の破壊的影響を軽減する可能性がある[87]。腸由来のセロトニンは、特定のセロトニン受容体を介して樹状細胞、マクロファージ、T細胞などの免疫細胞に作用し、それによって炎症メディエーターの放出と腸内微小環境に影響を与える可能性がある[88]。腸由来のγ-アミノ酪酸は、末梢シグナル伝達と視床下部統合を介して視床下部-下垂体-副腎軸軸の活動を調節し、ストレス関連の神経内分泌反応を減少させ、腸内微小環境を改善し末梢の炎症状態を調節することによって間接的に睡眠関連ネットワークの調節に関与する可能性もある[72]。 |
| さらに、トリプトファン代謝産物である3-ヒドロキシキヌレニン(3-HK)とキノリン酸(QUIN)が不眠症に直接関連しているという証拠は限られているものの、それらの炎症関連および神経毒性生物学的効果は、睡眠障害における潜在的な役割を示唆している。炎症状態では、3-ヒドロキシキヌレニンは活性酸素種の生成とミクログリアの活性化を促進し、それによって神経恒常性を破壊し、睡眠障害の発症に寄与する可能性がある[89]。強力なN-メチル-D-アスパラギン酸受容体アゴニストであるキノリン酸は、カルシウム流入、酸化ストレス、および神経損傷を誘発し、それによって覚醒を高め、睡眠障害に寄与する可能性がある[90]。 |
| 3.4. 腸内細菌叢-腸-脳軸と概日リズム障害 |
| 概日リズムの乱れは、不眠症のもう一つの重要な特徴である。腸内細菌叢-腸-脳軸が宿主の概日リズム調節に双方向的な影響を及ぼすことを示唆する証拠が増えつつある。一方では、宿主の中核時計遺伝子が、摂食・絶食リズム、局所的な腸上皮機能、および免疫微小環境を調節することによって、腸内細菌叢の日周振動パターンを形成する可能性がある[91]。Bmal1欠損症(訳者注:「Bmal1欠損症」という単一の先天性疾患がヒトに存在するという医学的な診断名はなく、これは体内時計(概日リズム)を司る重要な時計遺伝子である「Bmal1」の欠損を指します。Bmal1が欠損した状態では、全身の代謝異常、早期老化、ADHD様症状、炎症の慢性化、筋肉組織の衰弱などが引き起こされることが動物実験(マウス)で確認されています)などの腸上皮時計機能の障害は、微生物のリズム性を著しく減弱させることが示されている[92]。 |
| 一方、腸内細菌叢由来の代謝物、特に短鎖脂肪酸は、宿主の末梢時計システムを調節する可能性がある[93]。短鎖脂肪酸、特に酪酸とプロピオン酸は、エピジェネティックな調節特性を持ち、ヒストン脱アセチル化酵素の阻害を介してPER2(訳者注:PER2(Period Circadian Regulator 2)は、哺乳類の概日リズム(体内時計)の形成において中心的な役割を果たす時計遺伝子およびその翻訳タンパク質です。体内のあらゆる細胞に存在し、約24時間周期の生理現象や代謝のコントロール、さらにはがん抑制に深く関わっています)などの概日遺伝子の発現に影響を与える可能性があり、概日恒常性の維持に潜在的な役割を果たしていることが示唆される[94]。さらに、宿主と腸内細菌叢の共代謝物である胆汁酸は、その合成と輸送において顕著な概日リズムを示し、睡眠不足はこれらのリズムを乱す可能性がある[95]。グルカゴン様ペプチド-1などの腸管由来のホルモン分泌も概日変動を示し、睡眠不足または概日リズムの乱れは分泌リズムを変化させる可能性があり、睡眠覚醒サイクルに関連する代謝調節に関与している可能性が示唆される[96]。 |
| 概日リズムの調節や睡眠開始に関わる重要なホルモンであるメラトニンも、腸内細菌叢の影響を受ける可能性がある。これまでの研究では、腸内細菌叢が短鎖脂肪酸やトリプトファン由来の代謝産物を調節することで、メラトニンの合成と分泌を間接的に促進する可能性が示唆されている[97]。一方、メラトニンは微生物の組成や機能を調節し、相互調節ループを形成する可能性もある[98]。 |
| 3.5. 腸内細菌叢と不眠症の関連性を裏付ける実験的および臨床的証拠 |
| 3.5.1. 動物モデルからの証拠:因果関係 |
| 動物実験は、腸内細菌叢が睡眠調節において因果的な役割を果たしていることを比較的強く示唆する証拠を提供している。まず、腸内細菌叢枯渇モデルにおいて、抗生物質投与または無菌状態は、マウスの睡眠覚醒構造と脳波パワースペクトルの両方を変化させることが示されている。ある研究では、広域抗生物質を4週間投与した後、マウスの盲腸内容物において、神経伝達物質機能に関連するアミノ酸およびビタミン代謝が有意に変化し、セロトニンやビタミンB6などの代謝産物のレベルが低下したことが報告されている[99]。同時に、睡眠分析では、ノンレム睡眠の異常な分布、レム睡眠関連リズムの変化、および睡眠覚醒移行の不安定性が明らかになり、腸内細菌叢が正常な睡眠構造の維持に寄与していることが示唆された[99]。 |
| 第二に、糞便微生物移植は因果推論をさらに強化した。不眠症患者の腸内細菌叢を無菌マウスに移植すると、頻繁な覚醒やノンレム睡眠時間の短縮、血清酪酸レベルの低下など、不眠症に似た表現型が移植動物に誘発された[100]。さらに、睡眠不足のドナーから健康なレシピエントに糞便微生物叢を移植すると、うつ病様行動、心機能障害、心筋線維症などの特定の有害な表現型が伝達された[101]。これらの知見は、腸内細菌叢が睡眠障害に関連する全身的な病理学的影響を媒介する可能性があることを示唆している。 |
| 3.5.2. 臨床観察によるエビデンス:関連性 |
| 臨床研究もまた、腸内細菌叢と不眠症との関連性を支持している。健常対照群と比較して、不眠症患者では腸内細菌叢の構造と多様性に変化が見られることが多い。いくつかの研究では、α多様性の低下に加え、特定の有益な分類群の減少と、潜在的に病原性のある微生物の相対的な増加が報告されている(表1)。報告されている具体的な分類群は研究によって必ずしも一致しているわけではないが、全体的な傾向としては、不眠症と腸内細菌叢異常との関連性が示唆される。 |
| 表1.臨床研究における不眠症患者の腸内細菌叢の構成と多様性の変化 |
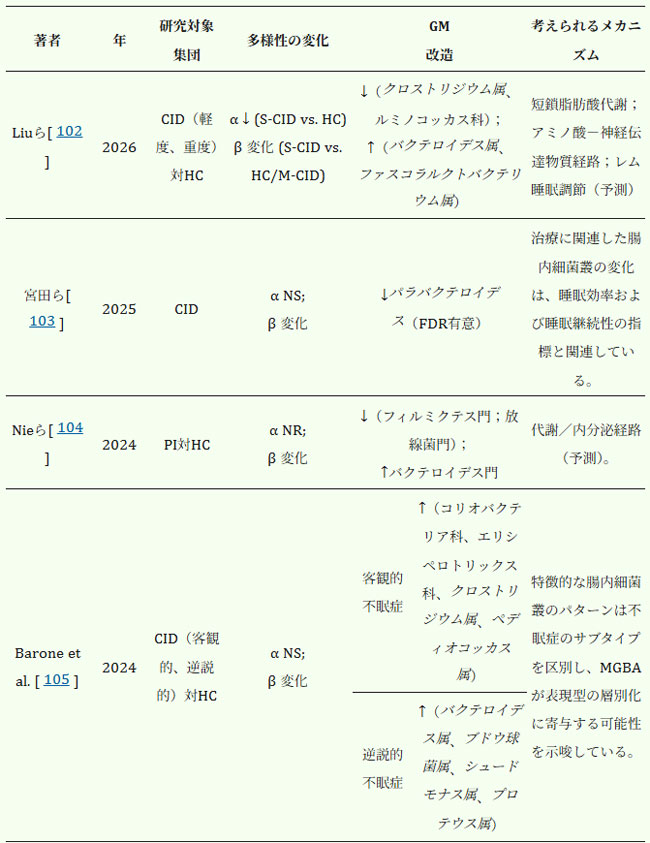 |
 |
| 腸内細菌叢の構造的変化に加え、不眠症患者ではしばしば微生物の代謝機能に異常が見られます。症例対照研究では、酢酸、プロピオン酸、酪酸、吉草酸を含む糞便中の総短鎖脂肪酸が、不眠症患者では健常対照群よりも有意に低く、短鎖脂肪酸レベルはピッツバーグ睡眠質問票(PSQI)および不眠症重症度指数(ISI)の両方と有意な負の相関関係にあることが示されました[110]。睡眠障害患者を対象とした別の研究では、腸内代謝物において「フェニルアラニン、チロシン、トリプトファン生合成」経路が有意に増加しており、神経伝達物質前駆体関連代謝の調節異常が示唆されました[111]。さらに、他の腸内細菌叢関連代謝物にも特徴的な変化が見られました。アデノシン、フェノール、フェニル硫酸塩を含む11種類の血清代謝物が、不眠症患者において有意に変化していた[112]。さらに相関分析を行った結果、アデノシン濃度はラクノスピラ属細菌の存在量および総睡眠時間と正の相関を示し、フェノールおよびフェニル硫酸塩濃度はコプロコッカス属細菌の存在量と負の相関を示し、ッツバーグ睡眠質問票および不眠症重症度指数スコアと正の相関を示した[112]。これらの知見を総合すると、腸内細菌叢関連代謝物の異常は、腸内細菌叢異常と睡眠障害の表現型発現との間の重要な関連性を示す可能性が示唆される。 |
| 4. 不眠症における腸内細菌叢-腸-脳軸に影響を与える因子 |
| 睡眠調節は、外因性因子と内因性因子の両方の影響を受け、その多くは腸内細菌叢-腸-脳軸の調節を介して影響を及ぼす可能性があります。外因性因子には、食事、生活習慣、薬剤曝露、環境因子に関連する概日リズムの乱れなどが含まれ、これらはすべて微生物組成と代謝産物を変化させる可能性があります[113–116]。 |
| 宿主の神経内分泌応答性、免疫状態、遺伝的感受性、および内在性概日リズム機構などの内因性因子は、微生物動態と宿主-微生物叢シグナル伝達をさらに調節する可能性があります[117]。これらの因子は双方向の相互作用を通じて、睡眠恒常性と概日リズムの安定性に影響を与え、それによって不眠症の発症と持続に寄与する可能性があります。 |
| 4.1. 食事とライフスタイル |
| 食事は腸内細菌叢の構成と代謝機能に大きな影響を与えます。高脂肪・高糖質摂取などの不健康な食習慣は、腸内細菌叢の恒常性を乱す可能性があります[118,119]。高脂肪摂取は炎症誘発性細菌の増殖を促進し、腸管バリア機能を損ない、免疫活性化や内因性ストレス反応を引き起こす可能性があり、睡眠恒常性の維持に悪影響を及ぼす可能性があります[118,120]。過剰な糖質摂取は血糖値の不安定化を招き、夜間覚醒の増加や睡眠断片化との関連が指摘されています[121]。マクロ栄養素に加えて、人工甘味料や乳化剤などの超加工食品成分への曝露は、微生物生態系やバリア機能を変化させる可能性があり[122]、睡眠関連の腸内細菌叢-腸-脳軸シグナル伝達における潜在的な役割を示唆しているが、不眠症に特化した証拠はまだ限られている。 |
| 生活習慣も腸内環境と睡眠の両方に影響を与える可能性があります。適度な運動を定期的に行うことは、一般的に腸内細菌叢の健全性と睡眠の質の向上につながるとされています[123]。一方、座りがちな生活は腸内細菌叢の構成と機能の変化につながり[124]、不眠症のリスク増加と関連付けられています[125]。短期間のアルコール摂取は鎮静作用をもたらす可能性がありますが、就寝前のアルコール摂取を繰り返すと、睡眠構造が乱れ、夜間の覚醒回数が増加し、睡眠の質が低下する可能性があります[126]。アルコールはまた、腸管透過性と炎症シグナル伝達を亢進させ、腸内細菌叢の構成を変化させ、間接的に感情、認知、睡眠関連行動に影響を与える可能性があります[127,128]。横断研究では、バランスの取れた食事、定期的な運動、禁煙、飲酒量の制限、座っている時間の短縮、適正体重の維持など、複数の健康的な行動を実践している人は、睡眠の質が向上し、睡眠時間が適切になり、不眠症のリスクが低くなることが示されました[129]。これらの知見は、腸内細菌叢-腸-脳軸の調節における外因性因子の重要性を強調しています。 |
| 環境曝露も見過ごしてはならない要因の一つです。新たな証拠によると、PM2.5などの微粒子状物質は腸管バリアの完全性を損ない、炎症反応を促進する可能性があり、それによって睡眠の安定性に関わる腸内細菌叢-腸-脳軸経路に影響を与える可能性があることが示唆されています[130]。さらに、交代勤務や夜間の人工光への曝露などによる概日リズムの乱れは、微生物の概日リズムと宿主の概日リズムの同期を崩し、睡眠調節を阻害する可能性があります[131]。 |
| 4.2. ストレスと心理的要因 |
| ストレスは、腸内細菌叢-腸-脳軸に影響を与える重要な心理社会的要因です。職業上のストレス、対人関係の葛藤、人生における大きな出来事などの心理社会的ストレス要因は、交感神経系を活性化させ、視床下部-下垂体-副腎軸を阻害することで、睡眠調節を損なう可能性があります[132,133]。不安やうつ病などの精神疾患は、不眠症と併発することが多く、双方向的に不眠症と相互作用する可能性があります[12,134]。ネガティブな感情、不適応な認知、慢性的な知覚ストレスは、睡眠の質を直接的に低下させるだけでなく[135–137]、腸内細菌叢の不均衡を伴い、心理的要因と睡眠障害の相互作用に寄与する可能性があります。研究によると、不安やうつ病の患者は腸内細菌叢の構成に異常を示すことが多く、一部の報告では、微生物の多様性の低下や有益な分類群の減少が示されています。[27,138]。これらの知見は、腸内細菌叢が心理的要因と睡眠との関連に関与している可能性を示唆している。さらに、臨床的証拠は、認知行動療法が不眠症の症状を改善し、併存する不安や抑うつ症状にも有益な効果をもたらすことを示しており[139,140]、不眠症の包括的な管理において心理的要因に対処することの重要性を強調している。 |
| 4.3. 薬剤および医療関連因子 |
| 薬剤および医療への曝露は、腸内細菌叢-腸-脳軸の安定性に影響を与える可能性のある外因性因子のもう一つのカテゴリーです。抗生物質は、腸内細菌叢を最も大きく撹乱する因子の一つであり、その使用は微生物の構成、多様性、代謝能力を著しく変化させ、ディスバイオシス(腸内細菌叢の不均衡)や免疫および神経化学シグナル伝達の障害を引き起こす可能性があります。[141,142] これらの影響は、新生児や小児において特に顕著であり、長期的な影響を及ぼす可能性があります。[143] |
| 動物実験では、抗生物質による腸内細菌叢の減少が、神経伝達物質関連の代謝を阻害し、睡眠構造を変化させることが示されている[99]。長期にわたる曝露は、代謝経路の変化や、不安様行動を含む神経行動異常を伴う可能性もある[144]。 |
| 抗生物質に加えて、エストロゲン、プロゲスチン、甲状腺ホルモンなどのホルモン剤も、微生物の増殖、付着、バイオフィルム形成、短鎖脂肪酸プロファイルに影響を与える可能性があります[145]。デキサメタゾンなどのグルココルチコイドは、微生物の量と多様性を減少させる可能性があり、免疫抑制と代謝の変化に関連しており[146]、視床下部-下垂体-副腎軸、免疫、および腸内細菌叢間のフィードバックループを阻害することにより、睡眠に影響を与える可能性があります。鎮痛剤、特にオピオイドも、ディスバイオシスを誘発し、行動調節に影響を与える可能性があります。一方、疼痛治療自体が睡眠構造の変化と密接に関連しています[147,148]。 |
| 臨床観察によると、がん治療に関連する神経精神症状は、睡眠関連症状を含む腸内細菌叢の異常と関連している可能性があり、化学療法、放射線療法、その他の医療曝露によって誘発されるディスバイオシスが睡眠障害の一因となっている可能性がある[149]。手術や麻酔も同様に腸内細菌叢の変化を誘発し、トリプトファン-キヌレニン代謝を阻害し、バリア機能の障害や術後の睡眠障害につながる可能性がある[150]。 |
| 5. 腸内細菌叢-腸-脳軸を標的とした不眠症治療戦略 |
| 腸内細菌叢は、神経伝達物質代謝、視床下部-下垂体-副腎軸機能、免疫炎症調節、概日リズムに関わる経路を介して不眠症の発症と維持に関与している可能性があることから、腸内細菌叢-腸-脳軸を標的とした介入は不眠症治療の有望な方向性として浮上している(表2)。 |
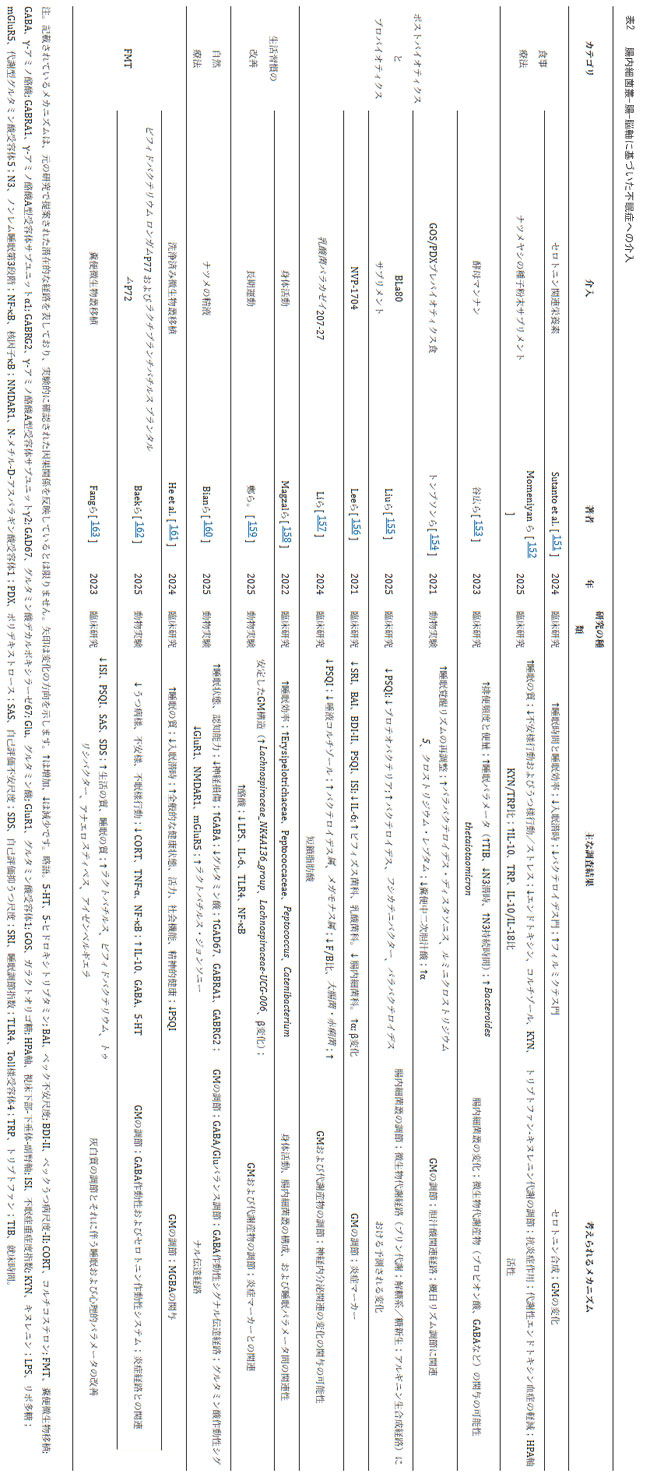 |
| 5.1. 食事療法 |
| 食事療法は、腸内細菌叢バランス(腸内細菌叢-腸-脳軸)を調節するための基本的かつ容易に実施可能な戦略です。観察研究では、野菜、全粒穀物、豆類、ナッツ類、オリーブオイルを多く摂取する地中海食を実践することで、睡眠の質が向上し、不眠症の重症度が軽減されることが示されています[164,165]。さらに、早朝の時間制限食など、日中を中心とした規則的な食事パターンは、腸内細菌叢と宿主の概日リズムの同期を維持するのに役立つ可能性があります。一方、夜間の食事や不規則な食事時間は、概日リズムの乱れ、腸内細菌の概日リズムの変動を阻害し、ディスバイオシス(腸内細菌叢の不均衡)を促進することで、不眠症のリスクを高める可能性があります[166,167]。 |
| 全体的な食事パターンに加えて、特定の栄養素も睡眠の質と関連付けられています。レビューでは、高炭水化物食や、チェリーなどのトリプトファン、メラトニン、植物栄養素が豊富な食品は、セロトニンとメラトニンの調節を含むメカニズムを介して、睡眠結果の改善と関連していることが示唆されています[168]。カテキン、アントシアニン、レスベラトロールなどの食事性ポリフェノールは、腸内微生物組成を調節し、酸化ストレスと炎症反応を軽減することで、睡眠の質に潜在的な利益をもたらす可能性があります[169,170]。さらに、高繊維食は有益な細菌の量を増やし、短鎖脂肪酸の産生を促進する可能性があり、腸粘膜バリアの完全性を維持し、炎症を軽減し、腸脳軸内の代謝シグナル恒常性を維持するのに役立つ可能性があります[171,172]。 |
| 5.2. プレバイオティクスとプロバイオティクス |
| プレバイオティクスは、選択的に発酵可能な基質として、有益な微生物の増殖を促進し、腸内環境を改善することができます[173]。動物実験では、プレバイオティクスを豊富に含む食事が糞便中の微生物組成を変化させ、睡眠不足後の睡眠回復を促進することが示されています[174]。慢性的な概日リズム障害のモデルでは、プレバイオティクスの介入が微生物組成と胆汁酸プロファイルを調節し、睡眠と概日リズムの再同期を促進し、併発する不安や抑うつ症状を軽減する可能性も示されています[154]。 |
| 現在、プロバイオティクスは睡眠障害に対する非薬物療法としてますます注目を集めている。いくつかのランダム化比較試験では、特定のプロバイオティクス株の補給が睡眠の質を改善したり、睡眠時間を延長したりする可能性があることが示されている[153,157]。メカニズム的には、プロバイオティクスは短鎖脂肪酸の産生を促進し、γ-アミノ酪酸とセロトニンを含む神経伝達物質関連経路を調節し、腸管バリアの完全性と炎症状態を改善することで、中枢神経系の機能、睡眠、感情状態に影響を与える可能性がある[175,176,177,178]。 |
| しかし、これらの介入の効果は、菌株の特異性、投与量、治療期間、宿主の特性によって大きく左右される。抗生物質治療を受けている新生児を対象とした無作為化二重盲検試験では、プロバイオティクス補給により睡眠時間が長くなり、泣き声が減少することが示されたが、統計的に有意な差は認められなかった[179]。したがって、標準化された治療プロトコルを確立するためには、大規模かつ厳密に管理された臨床試験が依然として必要である。 |
| 5.3. 生活習慣介入 |
| 定期的な運動は、一般的に睡眠の質の向上と不眠症症状の軽減に関連しています。運動は、体温調節、ストレス軽減、概日リズムの強化、気分の改善など、複数のメカニズムを通じて睡眠を促進する可能性があります[180]。また、運動が腸内細菌叢の構成を変化させる可能性を示唆する新たな証拠も得られています。定期的な中強度の運動は、腸内細菌叢の多様性の増加、短鎖脂肪酸産生菌の増加、腸管バリア機能の改善と関連付けられています[181]。 |
| さらに、最近のレビューでは、睡眠、腸内細菌叢、および心身医学の間の本質的な関係について体系的に議論されており、ヨガ、瞑想、マッサージなどの心身介入は、視床下部-下垂体-副腎軸、自律神経系活動、および神経免疫経路を調節することによって微生物生態系に影響を与え、それによって睡眠の質を改善する可能性があることが示唆されています[182]。 |
| 5.4. 糞便微生物移植 |
| 睡眠調節における腸内細菌叢の因果的役割を裏付ける前臨床的証拠に基づき、睡眠障害に対する糞便微生物移植の治療可能性を探る臨床研究が始まっている。これまでの研究結果によると、健康なドナーから採取した洗浄済み腸内細菌叢を睡眠障害患者に移植すると、入眠潜時が有意に短縮され、睡眠時間が延長し、睡眠の質が全体的に改善するとともに、腸内細菌叢の構造変化も伴うことが示されている[161]。さらに、前向き研究では、糞便微生物移植がCOVID-19急性期後の不眠症症状に有効である可能性が示唆されており、複雑な睡眠障害に対する治療可能性が強調されている[183]。実臨床研究でも、糞便微生物移植は慢性不眠症患者の睡眠と情緒状態を改善し、腸内細菌叢の再構築も伴うことが示されている[163]。興味深いことに、同じ研究の不眠症ではないサブグループでは、糞便微生物移植は主観的な睡眠の質の改善とも関連しており、微生物叢を標的とした介入はより広範な睡眠調節効果を持つ可能性があることを示唆している[163]。 |
| これらの予備的な結果は有望ではあるものの、糞便微生物移植は不眠症の分野においてはまだ探索段階にあり、その有効性を確認し、長期的な安全性を判断するためには、厳密な無作為化比較試験が依然として必要である。 |
| 5.5. 天然薬 |
| 腸内細菌叢-腸-脳軸の調節を介して不眠症を改善するために、伝統的な中国医学とその有効成分を使用する研究が増加している。関連するレビューでは、ナツメ種子、霊芝、茯苓多糖類、ジンセノサイド、および黄耆多糖類が、腸内細菌叢の組成を調節し、有益な代謝産物の産生を促進し、粘膜バリアの完全性を高め、炎症および神経伝達物質関連経路を調節することによって、睡眠リズムと睡眠の質の調節に関与する可能性があることが示唆されている[184]。 |
| さらに、ラクトバチルス・プランタラムP8と伝統的な中国漢方成分(ナツメ、ユリの球根、クチナシ、ブクリョウ)を組み合わせた評価を行った研究では、この組み合わせ介入により睡眠時間が有意に延長し、入眠潜時が短縮し、脳組織中のセロトニンとγ-アミノ酪酸の放出が増加することが示されました。その睡眠促進効果は、腸内細菌叢によって媒介される腸内細菌叢リモデリングと代謝および神経シグナル伝達関連経路の調節に関連している可能性があります[185]。これらの知見は、天然薬が単独で、または腸内細菌叢ベースの製剤と組み合わせて、不眠症介入のための新しい腸内細菌叢-腸-脳軸標的戦略となる可能性を示唆しています。 |
| 6. 結論と今後の展望 |
| 蓄積された証拠は、腸内細菌叢-腸-脳軸が不眠症の発症と維持において重要な調節システムであることを裏付けています。従来の中枢神経系中心モデルを超えて、腸内細菌叢-腸-脳軸は神経系、内分泌系、免疫炎症系、概日リズムの経路を統合し、末梢の微生物シグナルと中枢の睡眠覚醒調節を結びつける協調的なネットワークを形成します。腸内細菌叢-腸-脳軸は、神経伝達物質代謝、視床下部-下垂体-副腎軸活性、免疫恒常性、概日リズムに影響を与えることで、不眠症の複雑な病態生理を理解するための体系的な枠組みを提供します。重要なのは、これらの経路は独立して機能するのではなく、動的かつ相乗的に相互作用し、それによって過覚醒状態を維持し、不眠症の発症と慢性化を促進するということです(図2)。 |
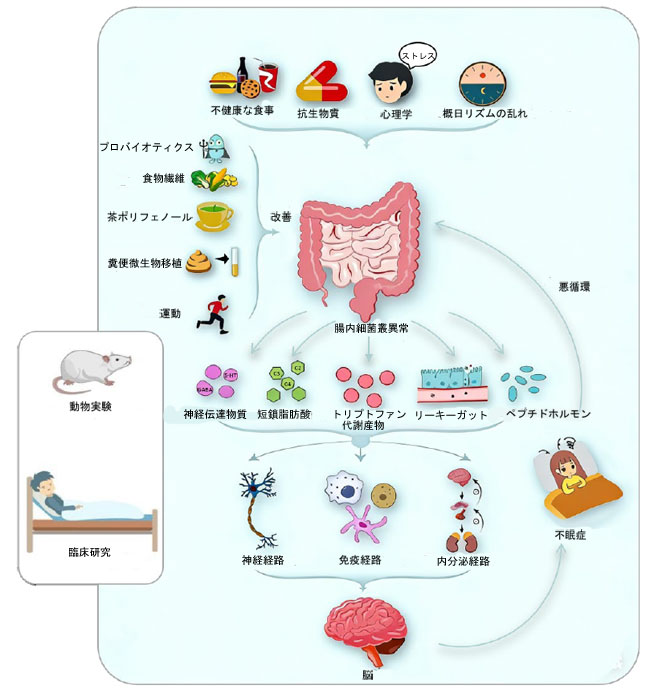 |
図2. 不眠症のメカニズムと腸内細菌叢を標的とした介入。 動物実験や臨床研究の証拠から、不健康な食習慣、抗生物質への曝露、心理的ストレス、概日リズムの乱れなどの要因が腸内細菌叢の異常を引き起こす可能性があることが示唆されている。この異常は、神経伝達物質、短鎖脂肪酸、トリプトファン代謝物、ペプチドホルモンなどの腸内細菌叢由来代謝物の産生を変化させるとともに、腸管透過性(リーキーガット)を増加させる可能性がある。これらの変化は、神経系、免疫系、内分泌系の経路を介して中枢神経系に影響を与え、不眠症の発症と進行に寄与する可能性がある。さらに、不眠症は腸内細菌叢をさらに乱し、悪循環を形成する可能性がある。高繊維食、茶ポリフェノール、定期的な運動、プロバイオティクス補給、糞便微生物叢移植など、腸内細菌叢を標的とした介入は、不眠症の症状を緩和する治療効果をもたらす可能性がある。 |
| 臨床応用および臨床的観点から、腸内細菌叢-腸-脳軸を標的とすることは、不眠症管理において有望かつ潜在的に改善可能なアプローチとなる。食事療法、プレバイオティクスおよびプロバイオティクス、生活習慣の改善、糞便微生物移植、天然薬など、腸内細菌叢を標的とした様々な介入は、睡眠の質および関連する神経心理学的アウトカムの改善において予備的な効果を示している(図2)。注目すべきは、これらの戦略は、短鎖脂肪酸産生の促進、神経活性代謝物の調節、腸管バリア機能の回復、全身性炎症の軽減など、収束的なメカニズムを介して作用する可能性があることである。 |
| これらの進歩にもかかわらず、いくつかの重要な限界を認識しておく必要があります。入手可能なヒトを対象とした研究のほとんどは横断研究または症例対照研究であり、因果関係の推論が制限されています。不眠症のさまざまな段階における動的な腸内細菌叢の変化を捉えた縦断的データは依然として不足しています。さらに、介入研究は、サンプルサイズが小さい、追跡期間が短い、アウトカム指標が標準化されていないといった制約を受けることが多く、結果の不一致につながっています。加えて、年齢、性別、代謝状態、心理的要因、およびベースラインの腸内細菌叢構成に関連する個人間の大きなばらつきが、結果の解釈をさらに複雑にしています。これらの限界は、現在のエビデンス基盤における既存のギャップと、腸内細菌叢-腸-脳軸研究を臨床実践に活かす上での課題を浮き彫りにしています。 |
| 今後の研究では、因果関係を確立し、臨床的に関連のある微生物の特徴を特定するために、大規模な多施設共同縦断研究と、適切に設計されたランダム化比較試験を優先的に実施すべきである。メタゲノミクス、メタボロミクス、トランスクリプトミクス、神経画像解析を含むマルチオミクスアプローチの統合は、腸内細菌叢と中枢睡眠調節系との動的な相互作用を解明するために不可欠となるだろう。さらに、微生物叢に基づく層別化と反応表現型の探索は、不眠症における精密医療戦略の開発を促進する可能性がある。 |
| 結論として、腸内細菌叢-腸-脳軸は不眠症に関する従来の神経生物学的パラダイムを超えた理解を広げるだけでなく、革新的で個別化された介入戦略を開発するための多次元的なプラットフォームを提供する。メカニズムに関する知見と臨床応用を結びつけることは、腸内細菌叢-腸-脳軸を標的としたアプローチを効果的な治療法へと転換し、将来の精密睡眠医療におけるその役割を確立するために不可欠となるだろう。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
| この文献は、Nutrients 2022, 14(20), 4300に掲載されたThe Microbiota–Gut–Brain Axis in Insomnia: Mechanisms and Intervention Strategies.を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |