Yingchun Zhao et al., |
概要 |
| アロニアメラノカルパ(Aronia melanocarpa:以下アロニアと表示)は、さまざまな生物活性を持つ天然の薬用植物であり、その果実は食品や医薬品によく使用されます。アロニアメラノカルパ多糖類(Aronia melanocarpa polysaccharide:AMP、以下アロニア多糖類と表示)は、アロニア果実の主成分です。この研究では、アロニア果実から得られたアロニア多糖類のD-ガラクトース誘導による老化マウスの遅延と保護を評価し、老化マウスの腸内細菌叢の代謝に対するアロニア多糖類の補給の効果を調査しました。老化モデルは、D-ガラクトース(200 mg / kg〜1000 mg / kg)を3日1回12週間腹腔内注射することによって確立されました。 アロニア多糖類(100および200 mg / kg)は、6週間のD-ガラクトース誘発後、強制経口投与により毎日投与されました。結果は、アロニア多糖類投与が8アーム迷路テストによって決定された老化マウスの空間学習と記憶障害を有意に改善したことを示しました。 H&E染色は、アロニア多糖類が脳組織の病理学的損傷と構造障害を有意に逆転させたことを示しました。 アロニア多糖類は、アデノシン5'-リン酸(ATP)活性化プロテインキナーゼ(AMPK) / サーチュイン1(SIRT1) /核因子カッパ-B (NF-κB)および核関連因子-2(Nrf2 )/ ヘムオキシゲナーゼ-1(HO-1)シグナル伝達経路を調節することにより、老化した脳組織の炎症と酸化ストレスによる損傷を軽減しました。特に、アロニア多糖類は、ホスファチジルイノシトール3-キナーゼ(PI3K)/ プロテインキナーゼB (AKT )/哺乳類のラパマイシン標的タンパク質(mTOR)シグナル伝達経路とその下流のアポトーシスタンパク質ファミリーを活性化することにより、脳細胞のアポトーシスと神経学的欠損を軽減しました。重要なことに、16S rDNA分析は、アロニア多糖類投与が腸内細菌叢の組成と有益な細菌の豊富さを改善することにより、老化プロセスを大幅に遅らせることを示しました。要約すると、この研究は、アロニア多糖類が炎症を抑制し、腸内微生物を調節することによってマウスの脳の老化を遅らせることを発見しました。これは、老化と関連する代謝性疾患の改善と治療の可能性を提供します。 |
序章 |
老化は、生物とそれに関連する神経の機能に影響を与える変性疾患です(1)。避けられない生理学的プロセスとして、加齢は身体機能の段階的な喪失を引き起こし、糖尿病、認知障害、癌、肝障害、パーキンソン病、アテローム性動脈硬化症など、多くの加齢性疾患を引き起こします(2,3)。先進国と発展途上国はどちらも、科学技術の進歩により人口が高齢化しています。したがって、老化を遅らせることは多くの研究の焦点となっています(4)。アンチエイジングの研究は、主にテロメアの短縮の抑制、脂質過酸化への抵抗、フリーラジカルの除去、免疫内分泌分泌の調節、DNA損傷とオートファジーの低減に焦点を当てています(5–9)。アデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)は、生物学的エネルギー代謝の調節における重要な分子であり、細胞の成長、増殖、生存、およびエネルギー代謝の調節に重要です(10–12)。 アデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)は、哺乳類細胞の老化の調節に関与するサーチュイン1(S IRT1)、核因子カッパ-B (NF-κB)、p53(訳者注:がん抑制遺伝子の1種)などの一連の加齢に伴うシグナル伝達経路の調節に関与しています(13)。サイレンシングメッセージレギュレーター2関連酵素1(SIRTUIN 1)は、生体に見られるニコチンアミドアデニンジヌクレオチド(NAD) +依存性ヒストンデアセチラーゼのクラスです。 サーチュイン1(SIRT1)は、細胞の生存、アポトーシス、ストレス耐性、炎症、その他の生理学的活性化に関与しています。 サーチュイン1(SIRT1)の活性化は、生物学的寿命を延ばす重要な理由であり、p53および核因子カッパ-B (NF-κB)シグナル伝達経路を直接調節することにより、細胞の抗ストレス能力に影響を与えます(14,15)。エネルギー代謝におけるアデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)の調節機能と上記のシグナル伝達経路の直接的または間接的な調節は重要であるため、アデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)と加齢に伴うシグナル伝達経路との関係を調べて、老化の考えられるメカニズムを見つける必要があります。 |
| D-ガラクトースの長期投与はげっ歯類の老化を促進することが知られているため、d-ガラクトースによる老化モデルは、老化メカニズムの研究に徐々に使用されてきました(16)。 体内の過剰なD-ガラクトースは、脳内に活性酸素種(ROS)の蓄積を引き起こし、カタラーゼ(CAT)、スーパーオキシドジスムターゼ(SOD)、グルタチオン(GSH)などの抗酸化酵素の活性を阻害する可能性があります(17)。 それはレドックスバランス防御システムを破壊し、酸化ストレスによる損傷につながる可能性があります(18,19)。 重要なことに、D-ガラクトースによって誘発される脳の老化は、ミトコンドリアの機能障害を引き起こし、炎症性損傷、細胞アポトーシス、および脳由来の神経栄養因子の減少を誘発することにより、認知能力の低下につながる可能性があります(20)。 抗酸化防御システムの不均衡は、老化の過程で重要な役割を果たします(21)。 したがって、レドックスの動的バランスを維持することは、D-ガラクトースによって誘発される老化を遅らせるための有用な治療アプローチである可能性があります(22)。 |
| アロニアは北米原産であり、現在中国東北部で大規模に導入・栽培されています(23)。 2018年9月、アロニアは、中国の国民健康評議会によってノベルフードとして承認されました。主にフルーツジュース、ジャム、ソース、フルーツティー、栄養補助食品、ワインの製造に使用されます。また、ロシアや東欧諸国では、食品着色料の天然源として、また降圧薬、抗アテローム性動脈硬化薬としても使用されています(24–27)。 アロニアは、フラボノイド、ポリフェノール、多糖類、有機酸、食物繊維、その他の栄養素が豊富です。多糖類は、抗腫瘍、肝保護、抗炎症、抗ウイルス、抗酸化、抗菌に有効です(28,29)。現在の研究では、アンジェリカ、クコ、レンゲの多糖類など、伝統的な漢方薬の多糖類がアンチエイジングに使用されていることもわかりました。メカニズムの研究は、フリーラジカルを除去し(30–32)、染色体の末端のテロメアの長さに影響を与え、体の免疫系を調節することによって機能することを示しています(33–35)。この研究では、動物行動実験を使用して、空間学習およびD-ガラクトース誘発老化マウスの記憶能力に対するアロニア多糖類の効果を観察しました。また、アデノシン5'-リン酸(ATP)活性化プロテインキナーゼ(AMPK) / サーチュイン1(SIRT1 )/核因子カッパ-B (NF-κB)シグナル伝達経路によるNLRP3(訳者注:NLRP3は、樹状細胞(DC)、単球、マクロファージ等の抗原提示細胞(APC)に発現するタンパク質です)インフラマソーム上のアロニア多糖類のメカニズムについても調査しました。 16S rDNAは、マウスの各グループの糞便の腸内細菌叢が有意に異なることを示しました。これは、腸内細菌叢の違いの違いが加齢において重要であることを示唆しています。 |
| アロニア多糖類の単糖組成 |
| 精製された多糖類はDEAE-52によって得られました。 単糖組成は、アロニア多糖類がフコース(0.14%)ラムノース(0.73%)、 アラビノース(7.14%)、ガラクトース(10.61%)、グルコース(76.16%)、キシロース(2.31%)、マンノース(1.25%)、ガラクトウロン酸(1.43%)、グルクロン酸(0.16%)、およびマンヌロン酸(0.07%)で構成されていることを示しました。(図1A、B)。 グルコースが主成分で、ウロン酸の総比率は1.66でしたが、ウロン酸前のピークは溶媒のピークでした。 |
図1.アロニア多糖類の構造の分析 |
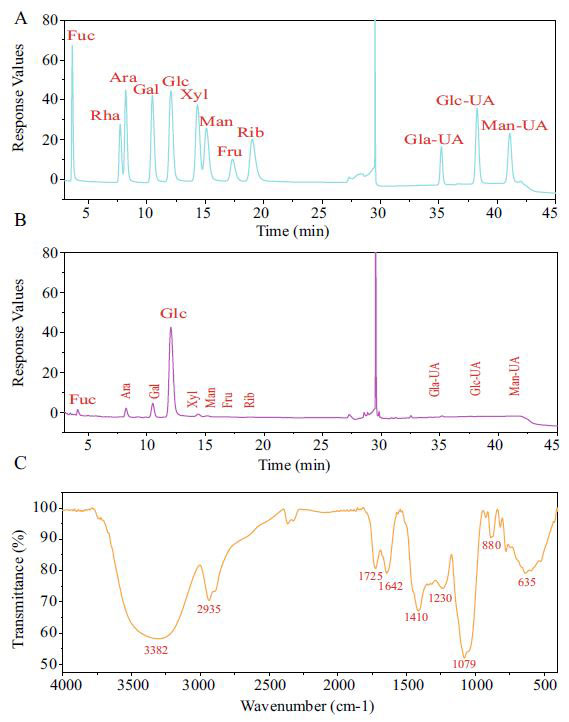 |
| (A)標準的な単糖。 (B)単糖構成。 (C)アロニア多糖類のFI-IR分析。 IRスペクトルはJASCOFT /で分析しました。IR-620分光計(http://www.jasco.co.jp/)。 [略語] Fuc:フコース,Rha:ラムノース,Ara:アラビノース,Gal:ガラクトース,Glc:グルコース, Xyl:キシロース,Man:マンマンノース,Gal-UA:ガラクツロン酸.Glc-UA:グルクロン酸, Man-UA:マンヌロン酸 |
| アロニア多糖類のFT-IRスペクトル分析 |
| アロニア多糖類の特徴的な構造を図1Cに示した。 3600〜3200 cm-1のピークは、-OHの伸縮振動吸収ピークであり、この領域の吸収ピークは糖の特徴的なピークでした。 詳細は次のとおりです。 382cm-1は、糖の特徴的なピークであるO–H伸縮振動吸収ピークでした。 2933 cm-1の吸収ピークは、C–H伸縮振動に起因していました(30)。 1650 cm-1の吸収ピークは、C = O伸縮振動に起因していました(32)。 1556 cm-1の吸収ピークは、C = O非対称伸縮振動に起因していました(34)。 1402 cm-1に吸収ピークがあり、これはC–O伸縮振動に起因していました。 1029 cm-1に吸収ピークがあり、これはO–H可変角度振動に起因していました。 1000〜800のいくつかの弱いピークは、αおよびβグルコシドが結合してピラノース環を形成していることを示しています(35)。 |
| アロニア多糖類はマウスの全身状態を改善します |
| マウスの初期および最終体重を記録し、体重増加率を分析した。 表1に示すように、 正常群と比較して、モデル群の体重は非常に有意な影響を及ぼしました(p <0.01)。 高濃度アロニア多糖類(AMP-H)グループはモデルグループと比較して有意に異なっていました(p <0.05)。 データは、D-ガラクトースよりもアロニア多糖類投与がマウスの体重に影響を与える可能性があることを示しました。 さらに、マウスの心臓、肝臓、脾臓、腎臓、脳を含むマウスの臓器指数を表2に記録しました。 結果は、アロニア多糖類がD-ガラクトースによって誘発されたマウスの臓器指数を効果的に改善し、各臓器の改善効果が異なることを示した。 |
表1.老化モデルマウスの体重に対するアロニア多糖類の効果 |
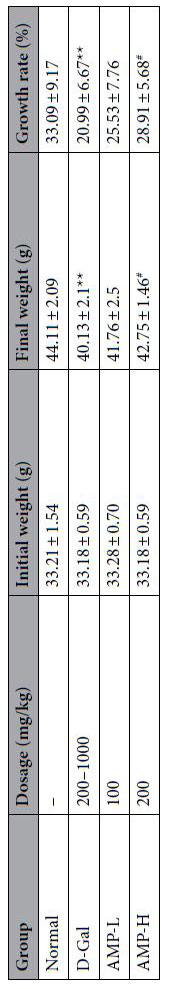 |
| 値は平均±S.D.、n = 10を表します。 * p <0.05または** p <0.01vs。通常のグループ; #p <0.05または## p <0.01 vs.D-ガラクトースグループ。 テーブルはを使用して描かれました。Microsoft Excel 2016バージョン3.3.2.13(https://www.microsoft.com/ es- cl / micro soft-365)。 |
表2.老化したマウスの臓器指数に対するアロニア多糖類の効果 |
 |
| 値は平均±S.D.、n = 10を表します。* p <0.05または ** p <0.01vs。通常のグループ; #p <0.05または## p <0.01 vs.D-ガラクトースグループ。 テーブルはMicrosoftExcelを使用して作成されました 2016バージョン3.3.2.13(https://www.microsoft.com/ es- cl / micro soft-365)。 |
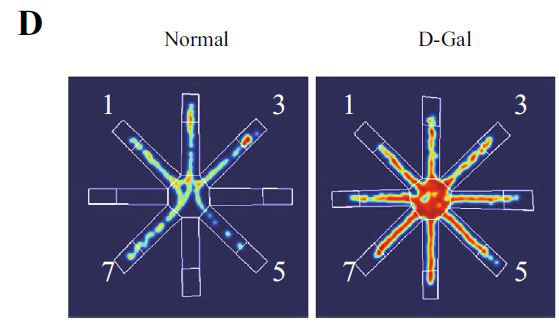
| 正常群のマウスは毛皮が滑らかで精神的に活発であり、D-ガラクトース群のマウスは丸くなり、毛皮はまばらで光がなく、精神状態は悪かった。 アロニア多糖類投与グループは、D-ガラクトースグループよりも毛皮と精神が優れていました。マウスの脳の外観から、D-ガラクトースグループは表面に出血スポットがあり、マウスの脳組織構造はぼやけていました(図2B)。 アロニア多糖類投与グループはD-ガラクトースグループよりも優れているようでした。脳の病的状態を調査するために、200倍および400倍の倍率で海馬歯状回の病理学的損傷を分析しました(図2C)。正常群と比較して、D-ガラクトースグループの顆粒細胞は不規則に配置され、核の形状が変化し、核が細胞質から分離し、細胞の形状が不完全であり、細胞の老化を示している(33,36)。 アロニア多糖類投与グループは、特に高濃度アロニア多糖類投与グループ(AMP-H)において、細胞の形態と配置を大幅に改善しました。 |
図2.アロニア多糖類はマウスの全身状態を改善します |
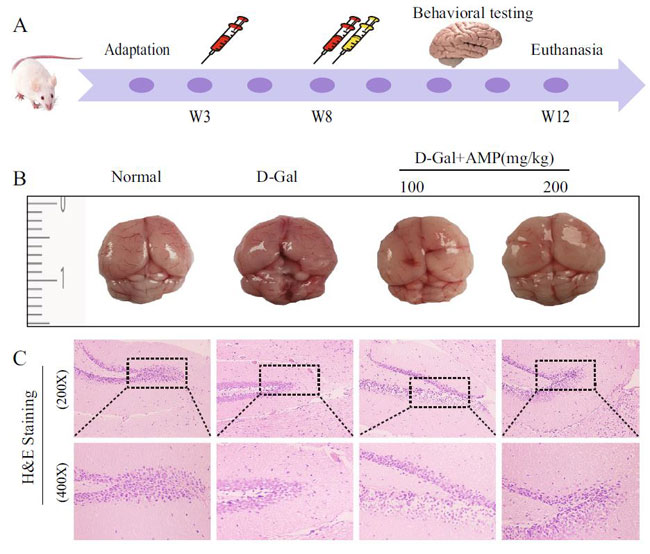 |
| (A)マウスにおけるD-ガラクトース誘発性肝線維症モデルの実験計画。 (B、C)アロニア多糖類は老化したマウスの脳全体の外観を改善し、H&E染色は海馬の病理学的損傷を示しました。 画像はCanonEOS 1600D \ u002Fを使用して取得されました 200D II(http://www.canon.com.cn/)、 Adobe Photoshop CC 2018(https://www.adobe.com/ product / photoshop.html)で処理されます。 図面は、Adobe Illustrator CS 11.0(https://www.adobe.com/ cn / product / illustrator.html)を使用して作成されました。 染色された切片は、光学顕微鏡Leica DM750(https://www.leica-microsystems.com.cn/ cn / product / light- micro scopes /)によって収集されました。 |
| アロニア多糖類はD-ガラクトースによって誘発される空間学習と記憶を改善します |
| 海馬は、ナビゲーションと空間記憶において重要な役割を果たします(33,37–39)。 各グループのマウスの空間記憶機能を研究するために、脳の空間学習と記憶機能を正確かつ高感度に反映する8アーム迷路テストを実施します。 このテストは、げっ歯類の空間記憶を検出するために使用される重要な実験方法です(40–42)。 結果は、D-ガラクトースグループのマウスの移動距離と潜時がより長く、アロニア多糖類投与がこの現象を大幅に改善したことを示しました(図3A–D)。 ただし、各象限のマウスの潜時に有意な関係はありませんでした。 この結果を検証するために、マウスの脳組織のアセチルコリンエステラーゼ(AchE)テストを実行しました(図3E)。 D-ガラクトースグループは正常群よりも有意に高く、アロニア多糖類投与後に減少した。 上記の結果はすべて、アロニア多糖類がD-ガラクトースによって引き起こされる老化現象を大幅に改善したことを示しています。 |
| 図3.アロニア多糖類は、D-ガラクトースによって誘発される空間学習と記憶を改善します |
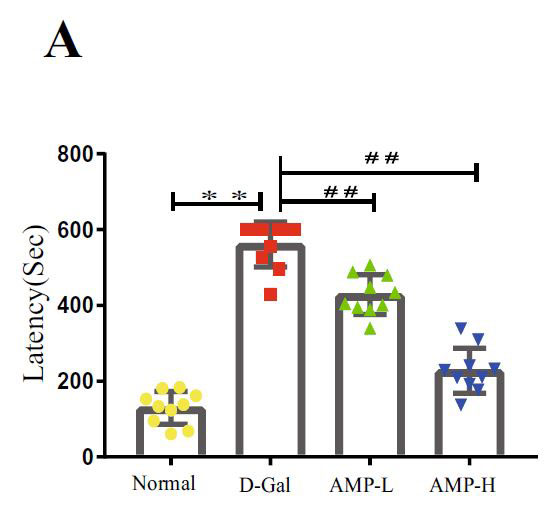 |
(A)迷路内のマウスの潜伏期間 |
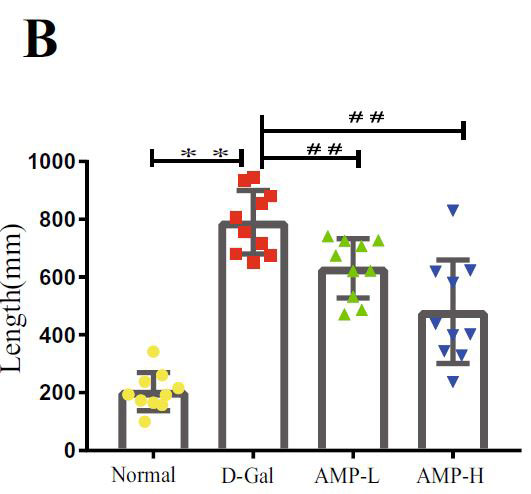 |
(B)迷路内のマウスのトラックの長さ |
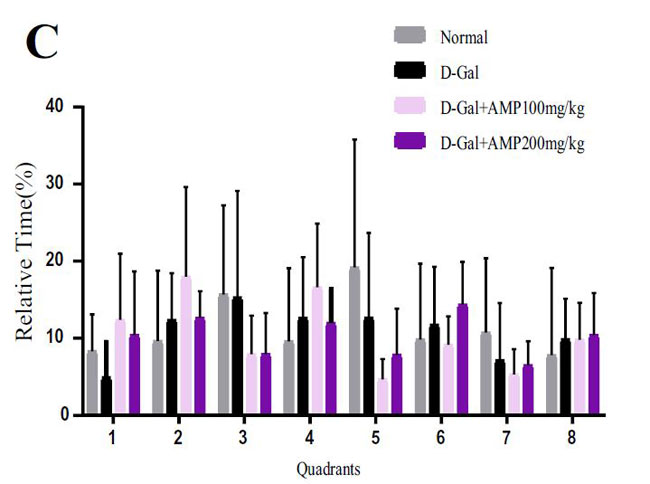 |
(C)各象限でマウスが費やした時間のパーセンテージ |
 |
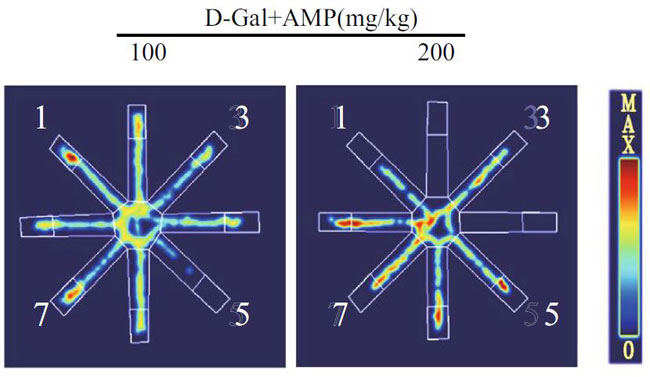 |
(D)迷路内のマウスの行動軌跡図 |
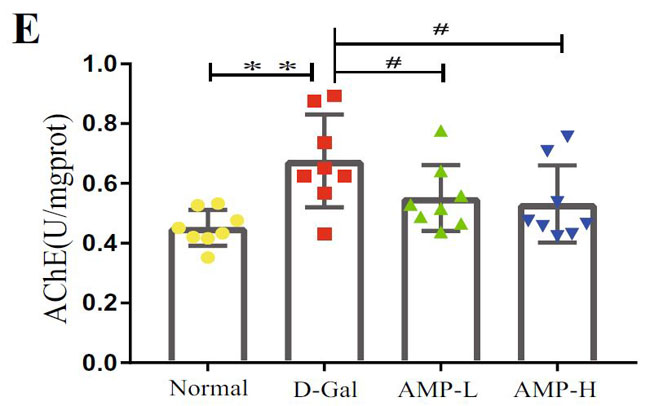 |
(E)マウス脳組織のアセチルコリンエステラーゼ(AchE)レベル |
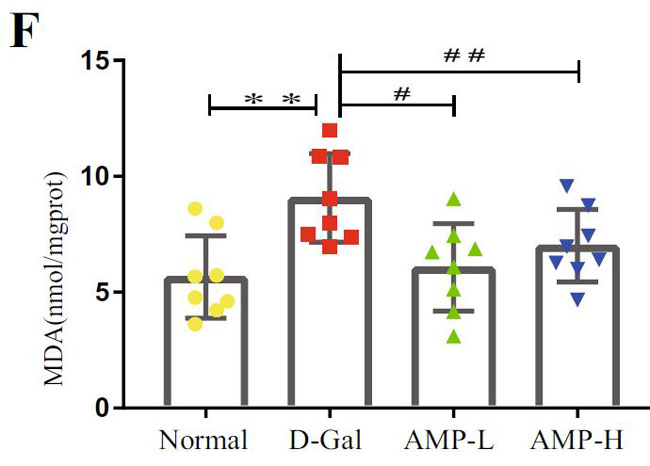 |
(F)D-ガラクトース誘発老化におけるマロンジアルデヒド(MDA)の脳レベル |
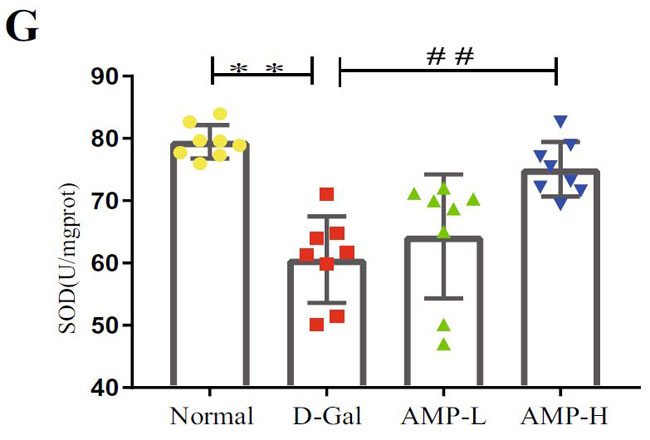 |
(G)D-ガラクトース誘発老化におけるスーパーオキシドジスムターゼ(SOD)の脳レベル |
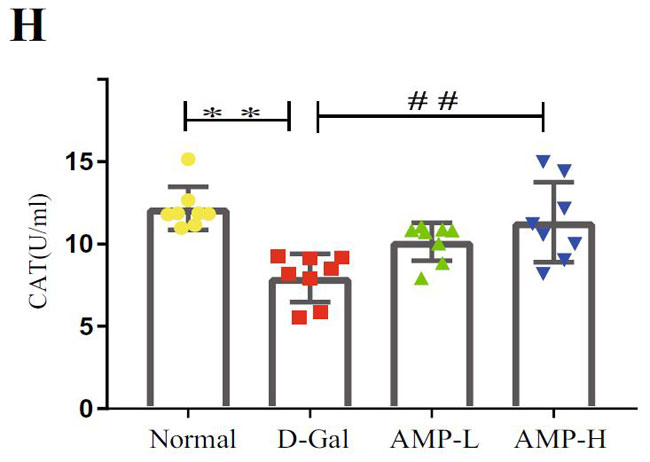 |
(H)D-ガラクトース誘発老化におけるカタラーゼ(CAT)の脳レベル |
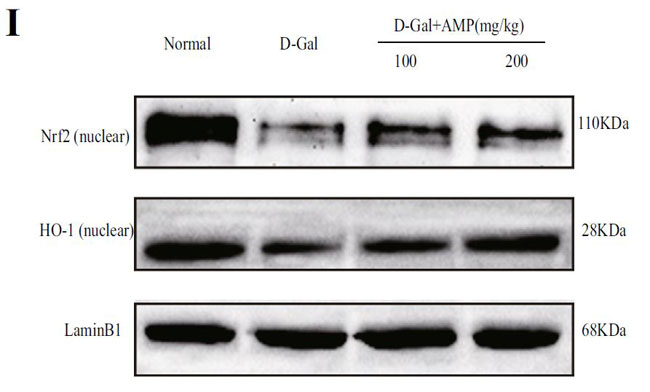 |
(I)核関連因子-2(Nrf2)およびヘムオキシゲナーゼ-1(HO-1)タンパク質の発現 |
|
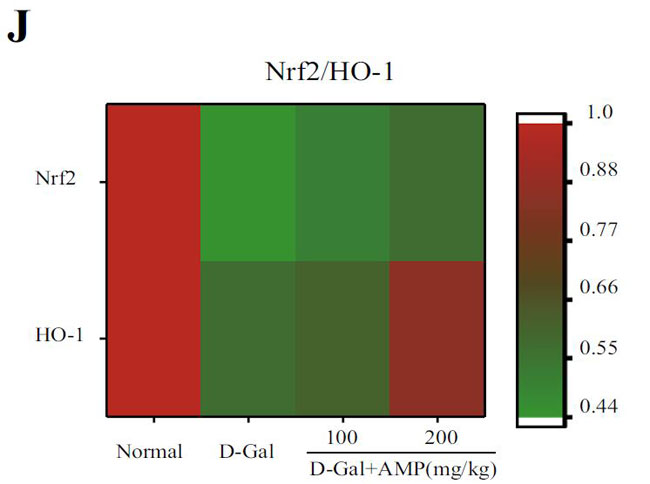
(J)核関連因子-2(Nrf2)およびヘムオキシゲナーゼ-1(HO-1)タンパク質の発現レベルのヒートマップ分析。 |
| データは、平均±標準偏差(S.D)、n = 10として表されます。* p <0.05または** p <0.01vs。通常のグループ。 #p <0.05または## p <0.01 vs.D-ガラクトースグループ。 図は、GraphPad Prism Softwareバージョン6.04(https://www.graphpad.com/)を使用して作成されました。化学イメージングは、Bio-Rad ImagingSystemバージョンのVersaDoc3000(https://www.bio-rad.com/)を使用して収集されました。 ヒートマップは、オンラインツールMorpheus(https://software.broadinstite.org/morph eus /)を使用して作成されました。軌道イメージングは、Thermal Imaging AnalysisソフトウェアバージョンRMT-100(https:// tmvmc.com/)を使用して撮影されました。 |
| アロニア多糖類はD-ガラクトースによって誘発される酸化ストレスを改善します |
| 酸化ストレスは老化プロセスにおいて重要な役割を果たしているため、マロンジアルデヒド(MDA)、スーパーオキシドジスムターゼ(SOD)、カタラーゼ(CAT)などの脳組織の関連指標を測定しました。 結果は、マロンジアルデヒド(MDA)が反対であるのに対し、D-ガラクトース(D-Gal)グループのスーパーオキシドジスムターゼ(SOD)とカタラーゼ(CAT)のレベルが大幅に減少したことを示しました(図3F–H)。 アロニア多糖類投与グループは状況を大幅に改善し、マロンジアルデヒド(MDA)レベルを低下させ(p <0.01またはp <0.05)、スーパーオキシドジスムターゼ(SOD)およびカタラーゼ(CAT)レベルを上昇させました(p <0.01)。 これらの結果は、アロニア多糖類がD-ガラクトースによって誘発される酸化ストレスに対して保護効果を持っていることを示しました。 |
| 老化したマウスに対するアロニア多糖類の抗酸化作用をさらに調査するために、核関連因子-2(Nrf2) /ヘムオキシゲナーゼ-1(HO-1)シグナル伝達経路をウエスタンブロッティングによって分析しました。 結果は、アロニア多糖類がD-ガラクトースグループの核関連因子-2(Nrf2)およびヘムオキシゲナーゼ-1(HO-1)核タンパク質の発現を有意にアップレギュレーションしたことを示しました(p <0.01)(図3I、J)。 上記の結果はすべて、アロニア多糖類がD-ガラクトースによって誘発される酸化ストレス損傷を改善したことを示しています。 |
| アロニア多糖類はアデノシン5'-リン酸(ATP)活性化プロテインキナーゼを介してNLRP3インフラマソームの産生を阻害します |
| アデノシン5'-リン酸(ATP)活性化プロテインキナーゼ(AMPK)とサーチュイン1(SIRT1)の間の循環調節メカニズムは、老化プロセスにおけるエネルギー代謝バランスの重要な役割を強化します。 サーチュイン1(SIRT1)は、P53遺伝子の活性を低下させることにより、老化プロセスの調節に関与します(14,15)。 そこで、老化関連タンパク質をテストし、その結果を図4A〜Dに示しました。 モデルグループのアデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)とサーチュイン1(SRIT1)の発現は有意に少なかったが、P53はその反対であった。 |
| 図4.アロニア多糖類は、D-ガラクトースによって誘発される酸化ストレスを改善します |
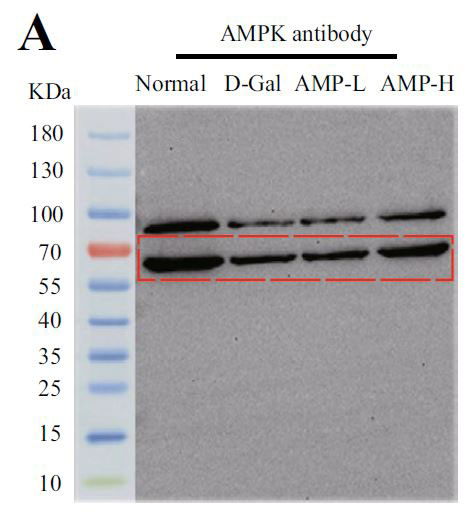 |
(A)アデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)の発現 |
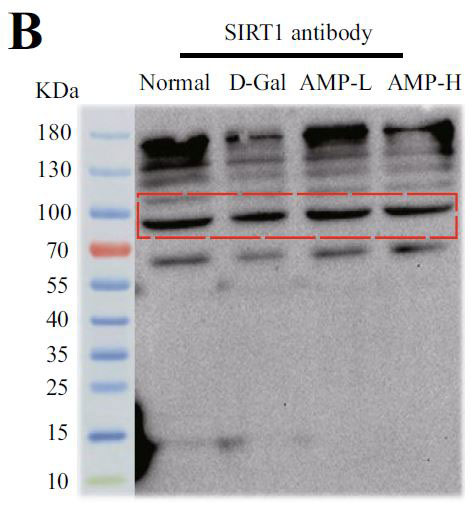 |
(B)サーチュイン1(SIRT1)の発現 |
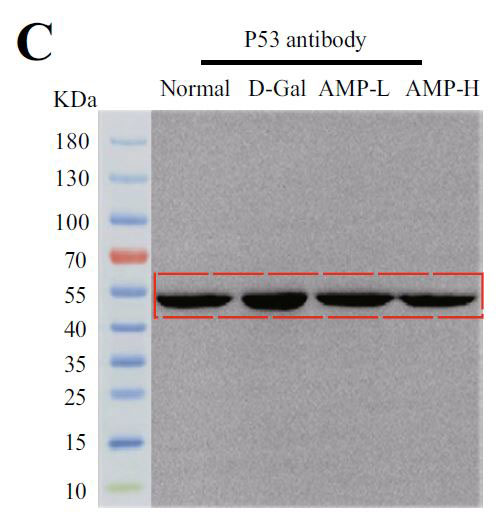 |
(C)P53タンパク質の発現 |
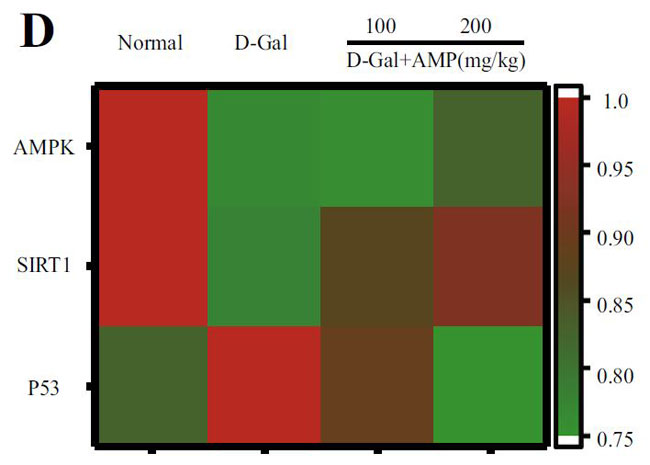 |
(D)アデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)、サーチュイン1(SIRT1)、およびP53タンパク質シグナル伝達経路の発現レベルのヒートマップ分析 |
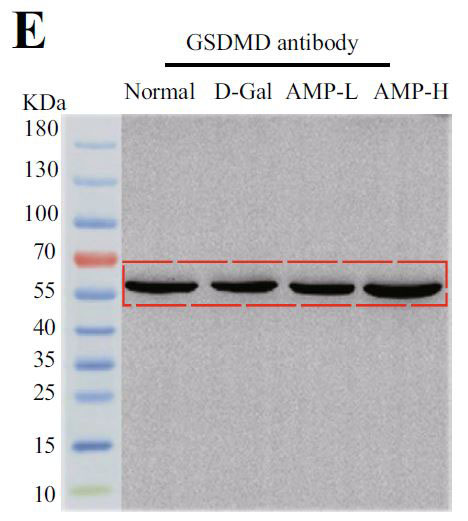 |
(E)ガスデルミンD (GSDMD)のインフラマソームタンパク質発現 |
|
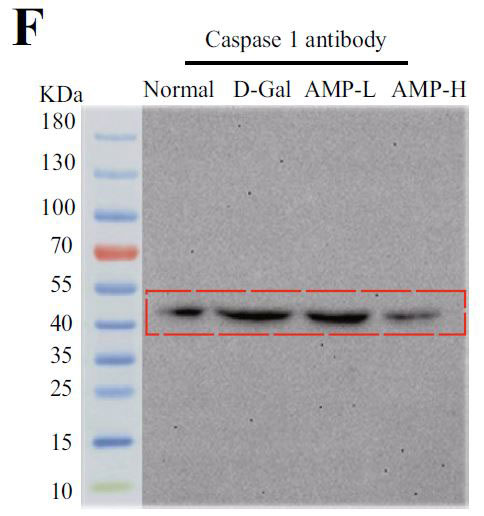
(F)カスパーゼ1のインフラマソームタンパク質発現 |
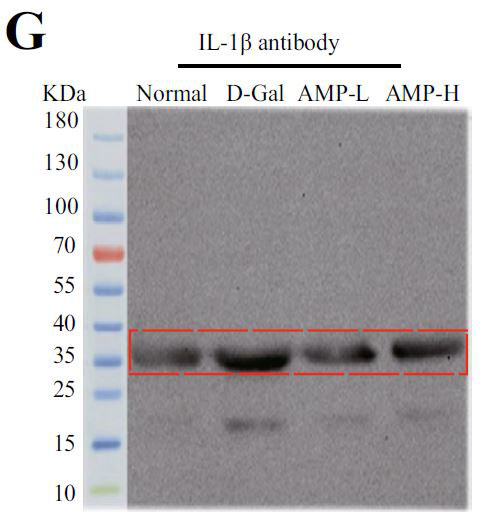 |
(G)組換えラットIL-1β(IL-1β)のインフラマソームタンパク質発現 |
 |
(H)ガスデルミンD、カスパーゼ1、組換えラットIL-1βインフラマソームタンパク質発現レベルのヒートマップ分析 |
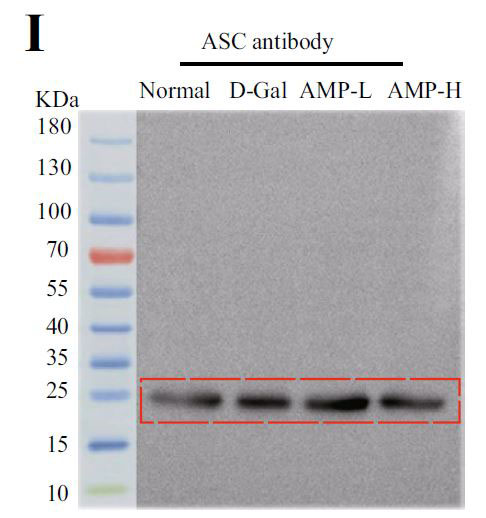 |
(I)脂肪由来幹細胞(ASC)のインフラマソームタンパク質発現 |
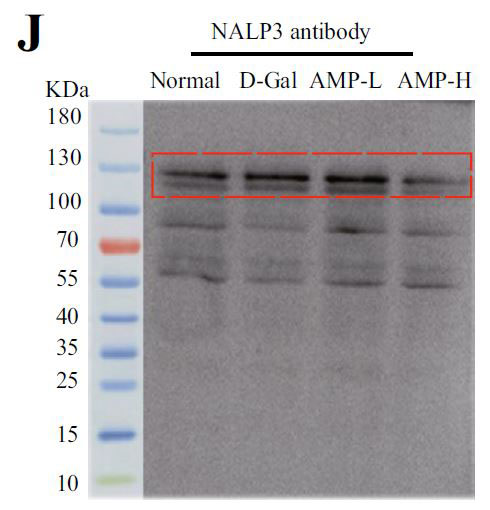 |
(J)NALP3のインフラマソームタンパク質発現、およびインフラマソームタンパク質発現レベルのヒートマップ分析 |
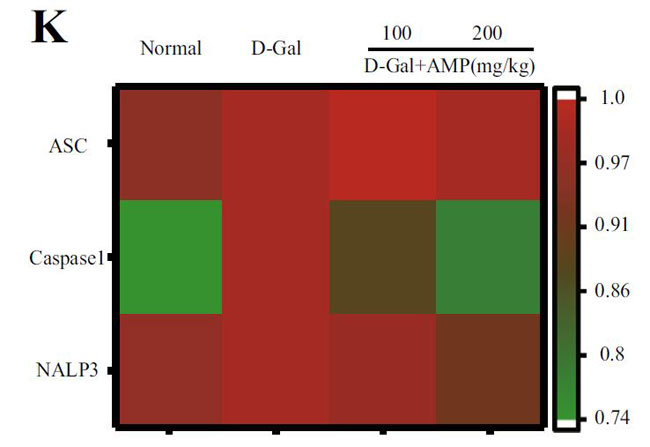 |
(K)ASC、およびNALP3のインフラマソームタンパク質発現レベルのヒートマップ分析 |
| データは、平均±標準偏差(S.D)、n = 10として表されます。* p <0.05または** p <0.01vs。通常のグループ。 #p <0.05または## p <0.01 vs.D-ガラクトースグループ。 化学イメージングは、Bio-Rad ImagingSystemバージョンのVersaDoc3000(https://www.bio-rad.com/)を使用して収集されました。ヒートマップは、オンラインツールMorpheusを使用して作成されました(https://software.broadinstite.org/morph eus /)。 |
| アロニア多糖類がD-ガラクトース誘発性老化に対して抗発熱作用を有するかどうかを判断するために、カスパーゼ-1によって媒介される古典的な発熱経路における関連タンパク質のレベルを検出しました。 D-ガラクトースグループのガスデルミンD(GSDMD)、カスパーゼ-1、およびIL-1βのタンパク質発現はすべて増加し、アロニア多糖類投与グループはこの現象を変える可能性があります(図4E–H)。 熱分解の発生は、インフラマソームの蓄積を伴うことがよくあります。 熱分解に対するアロニア多糖類の分子メカニズムをさらに調査するために、NLRP3およびASC(訳者注:脂肪由来幹細胞)タンパク質の発現に焦点を当てました。 正常グループと比較して、D-ガラクトースグループのNLRP3およびASCのタンパク質含有量は増加し、アロニア多糖類投与後にそれらの発現は減少しました(図4I–K)。 すべての結果は、アロニア多糖類がD-ガラクトースによって誘発される熱分解を軽減し、それによってマウスの老化状態を遅らせることを示しました。 |
| アロニア多糖類はD-ガラクトース誘発性炎症を軽減します |
| アロニア多糖類がd-ガラクトース誘発性老化に対して抗炎症作用を有するかどうかを調べるために、関連タンパク質のレベルを核因子カッパ-B NF-κBの古典的な炎症経路で測定しました。 D-ガラクトースを使用すると核因子カッパ-B(NF-κB)のリン酸化タンパク質とその上流のレギュレーターであるIκBの発現が増加しました(p <0.01)。 アロニア多糖類投与グループは、核因子カッパ-B (NF-κB)の高発現を有意に抑制し、上流の調節因子IκB-αリン酸化タンパク質の発現の増加をブロックすることができました(p <0.01)(図5A、B)。 これらの結果は、アロニア多糖類が抗炎症効果を通じてD-ガラクトース誘発性の老化症状を改善したことを示しています。 |
| 図5. アロニア多糖類は、核因子カッパ-B (NF-κB)を介した炎症とPI3K / AKT / mTORの抗アポトーシス経路を調節することにより、マウスの老化防止効果を発揮します |
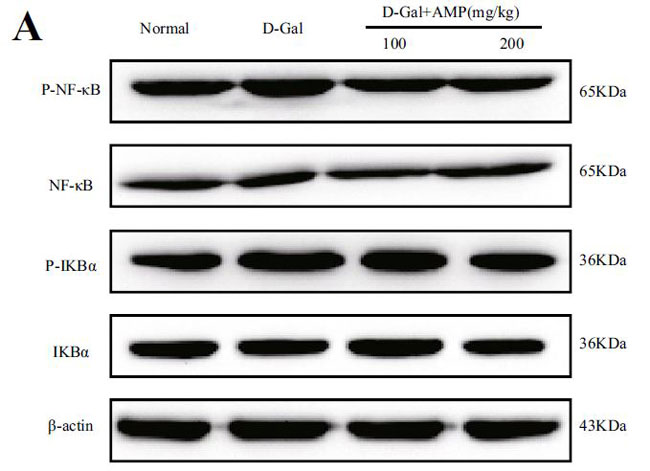 |
(A)脳組織のマウスにおける核因子カッパ-B (NF-κB)およびIκB-αのタンパク質発現 |
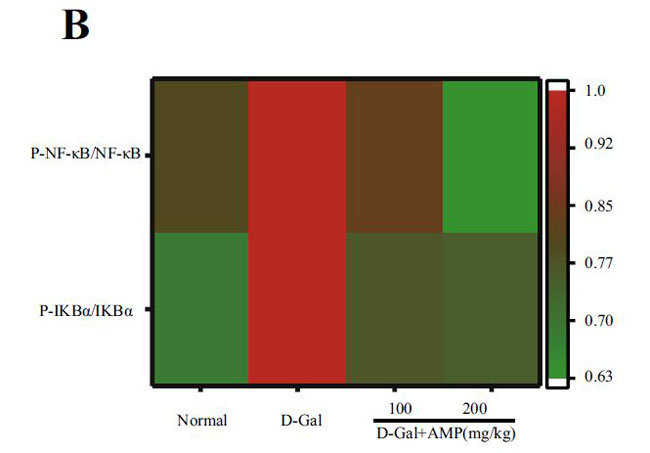 |
(B)脳組織のマウスにおける核因子カッパ-B (NF-κB)シグナル伝達経路発現タンパク質レベルのヒートマップ分析 |
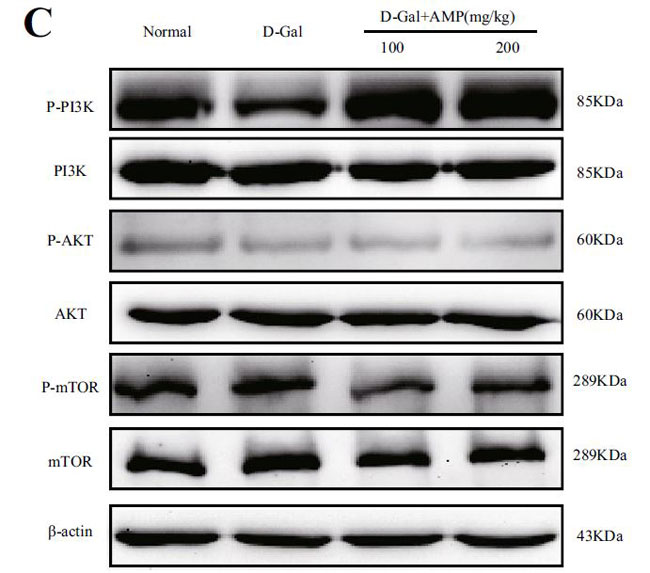 |
(C)脳組織のマウスにおけるp-PI3K / PI3Kのタンパク質発現 |
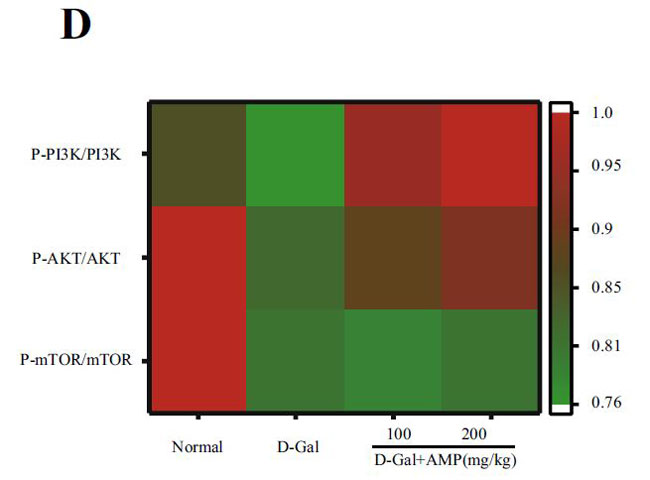 |
(D)PI3K / AKT /のヒートマップ分析 |
 |
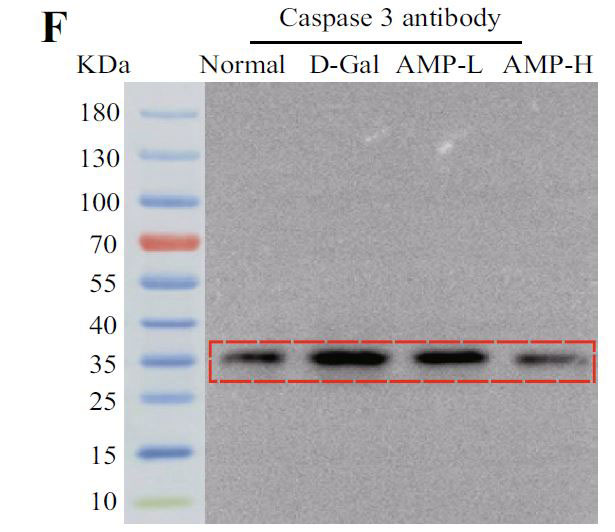
(E)脳組織のマウスにおけるアポトーシスタンパク質(Bax)のタンパク質発現 |
 |
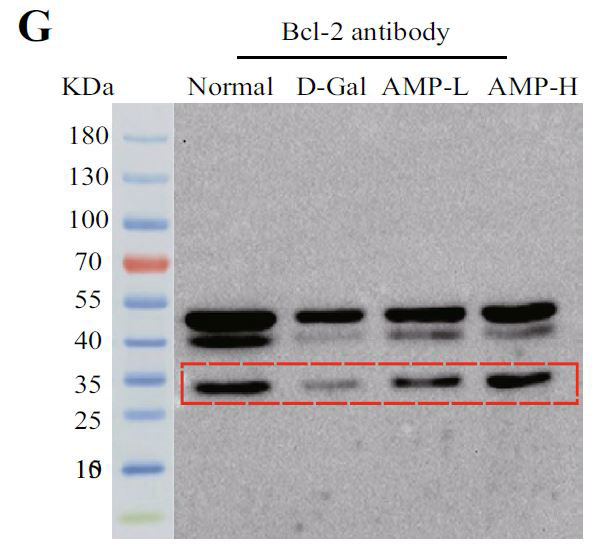
(F)脳組織のマウスにおける、Caspase 3のタンパク質発現 |
 |
(G)脳組織のマウスにおけるBcl-2のタンパク質発現 |
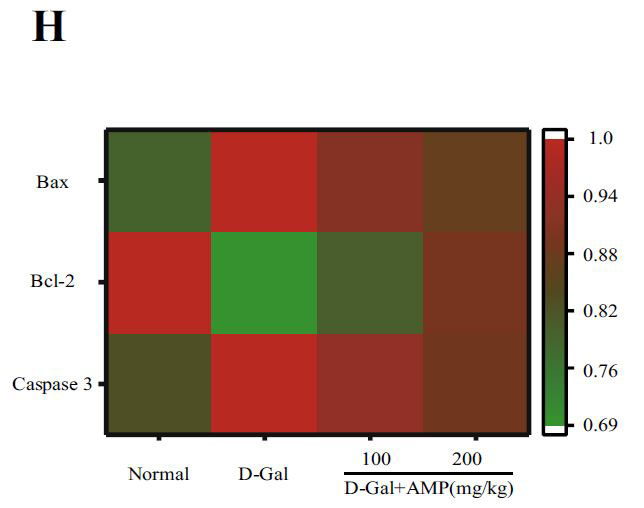 |
(H)PI3K / AKT /のヒートマップ分析 mTORとその下流のタンパク質レベル |
| データは、平均±標準偏差(S.D)、n = 10として表されます。* p <0.05または** p <0.01vs。通常のグループ。 #p <0.05または## p <0.01 vs.D-ガラクトースグループ。 化学イメージングは、Bio-Rad ImagingSystemバージョンのVersaDoc3000(https://www.bio-rad.com/)を使用して収集されました。ヒートマップは、オンラインツールMorpheus(https://software.broadinstite.org/morph eus /)を使用して作成されました。 |
| アロニア多糖類は、PI3K / AKT経路を介してD-ガラクトース誘導アポトーシスを低減します |
| 下流のアポトーシスタンパク質Bax、Bcl-2、およびカスパーゼ3を含むPI3K / AKT関連タンパク質を実行しました。 アロニア多糖類投与グループは、D-ガラクトースグループと比較してP-PI3K、P-AKT、およびP-mTORタンパク質の発現を有意に増加させました(p <0.05またはp <0.01)(図5C–H)。 さらに、その下流のアポトーシスタンパク質については、アロニア多糖類)はBaxおよびカスパーゼ3タンパク質の発現を有意に減少させましたが、Bcl-2の結果は反対でした(p <0.05またはp <0.01)。 これらの結果は、アロニアメラノカルパ多糖類(AMP)がD-ガラクトース誘導アポトーシスを阻害したことを示しています。 |
| アロニア多糖類は、マウスの腸内細菌叢を調節することにより、D-ガラクトースによる老化を改善します |
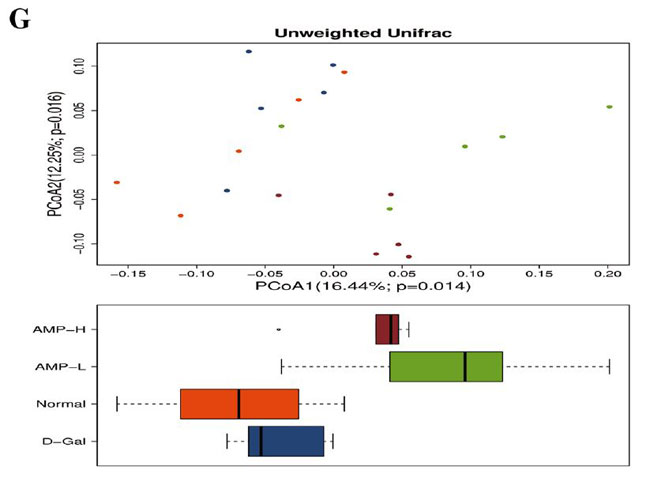
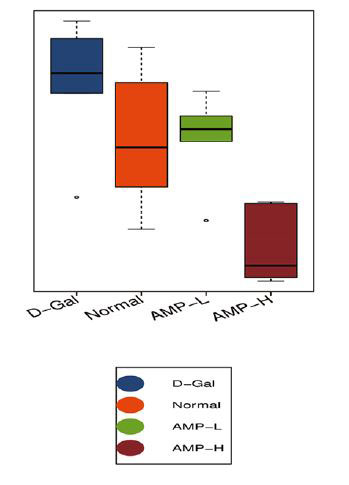
| ベン図は、正常グループ、D-ガラクトース(D-Gal)グループ、低濃度アロニア多糖類(AMP-L)、および高濃度アロニア多糖類(AMP-H)グループのOUT(訳者注:細菌の必須遺伝子(一般に,16SリボソームRNA遺伝子)の塩基配列をコンピュータ上でその類似度を指標に分類したときに得られる単位をいう)の数が、それぞれ473、520、509、および467であることを示しています。 通常グループとD-ガラクトースグループには435のOTUがありました。 低濃度アロニア多糖類(AMP-L)、高濃度アロニア多糖類(AMP-H)グループには合計458および434 OTUがあり、低濃度アロニア多糖類(AMP-L)、高濃度アロニア多糖類(AMP-H)グループには合計429 OTUがありました(図6A)。 種の存在量の表によると、バクテロイデス属とファーミキューテス属の門の相対的な存在量は、4つのグループで比較的増加しました。 アロニア多糖類投与後、バクテロイデス門の相対的な存在量は増加しましたが、ファーミキューテスの相対的な存在量は減少しました(図6B)。 4つのグループで有意に濃縮された優勢な細菌グループは、ラクトバチルス、アッケルマンシア、バクテロイデス、プレボテラ、バルネシエラ、アリスティペスでしたが、グループ間に有意差はありませんでした(図6F)。 |
| 図6. アロニア多糖類は、マウスの腸内細菌叢を調節することにより、D-ガラクトースによる老化を改善します |
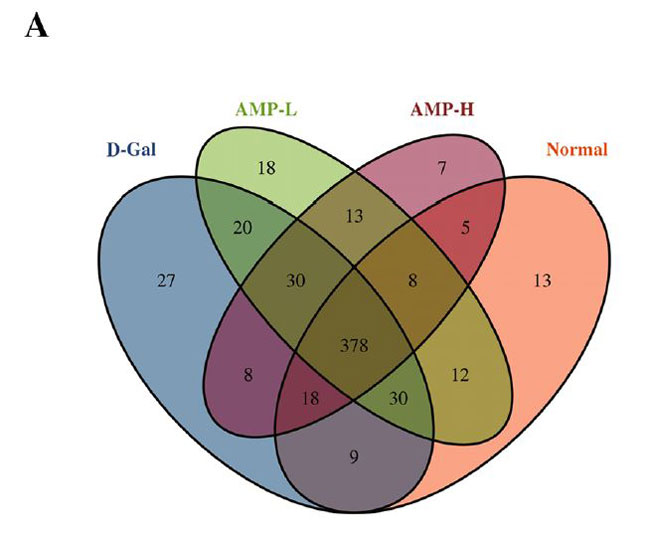 |
(A)ベン図 |
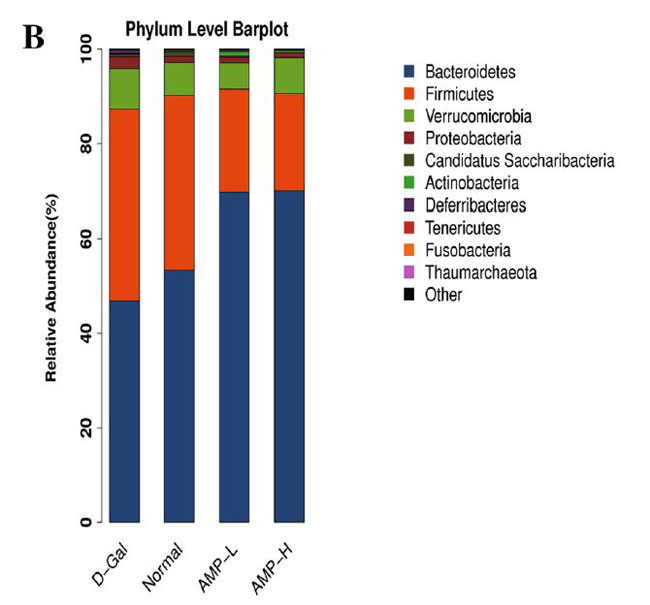 |
(B)門レバー棒グラフ |
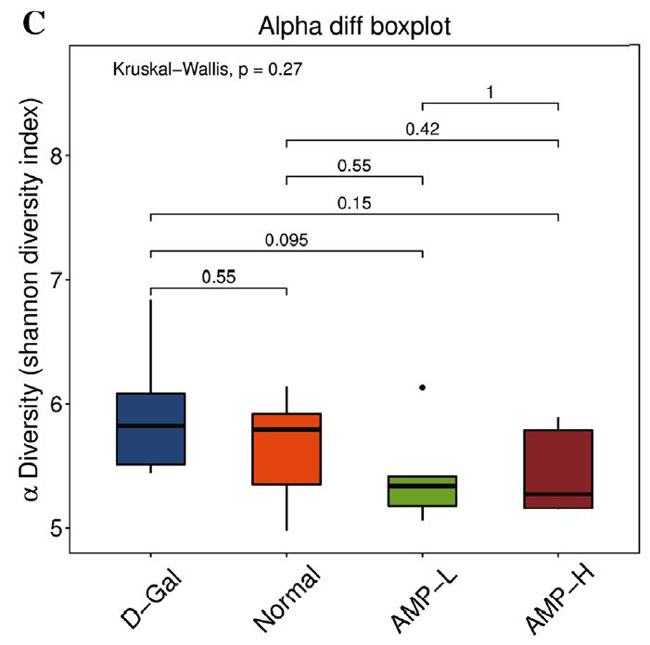 |
(C)アルファ差分箱ひげ図 |
 |
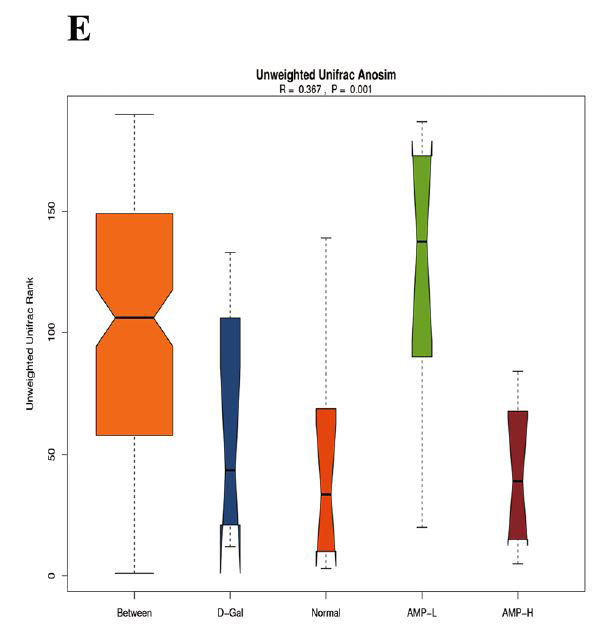
(D)アルファ差分箱ひげ図 |
 |
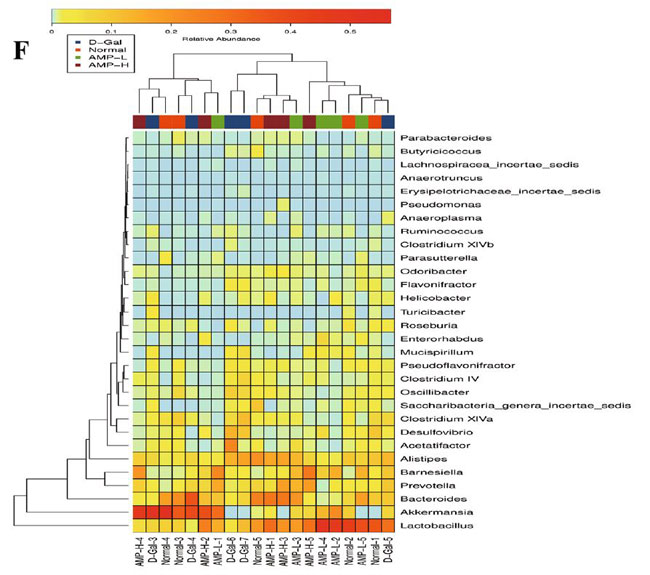
(E)重み付けされていないUniFracANOSIM |
 |
(F)相対的な存在量と重み付けされていないUnifrac |
 |
|
(G)相対的な存在量と重み付けされていないUnifrac |
| データは、平均±標準偏差(S.D)、n = 10として表されます。* p <0.05または** p <0.01vs正常群。 #p <0.05または## p <0.01 vsD-ガラクトースグループ。 OTUは、UPARSEバージョン7.1(http://drive5.com/ uparse /)を使用してクラスター化されました。画像は、R 3.6.3(https://www.r-project.org/)とpheatmapパッケージ(https://cran.r-project.org/web / packa ges / pheat map /)を使用して生成されました。 |
| 図6C、Dに示すように、αダイバーシティ(シャノンおよびシンプソン指数)は0.27から0.99に増加しました。 加重Unifracクラスタリング(pCoA、ANOSIM)に基づくβ多様性分析は、マウスの4つのグループ間の差がグループ内の差よりも有意に大きく、有意差があったことを示しました(R = 0.367、p = 0.001)(図.6E)。 pCoAダイアグラムは、4つのグループの植物相がさまざまな程度で変化したものの、低濃度アロニ多糖類(AMP-L)グループの微生物叢は正常なマウスのグループと類似しており、高濃度アロニア多糖類(AMP-H)グループの微生物叢は他のグループとは大幅に異なることを示しました。 (図6G)。 |
| D-ガラクトースは還元糖であり、酸化ストレスや組織の損傷を引き起こし、細胞のアポトーシスやミトコンドリアの機能障害を引き起こし、抗酸化防御システムの機能を低下させる可能性があります(37–39)。 齧歯動物に使用され、長期の前臨床老化モデルを実現します(43,44)。 実質的な研究により、薬用多糖類が老化を効果的に防ぐことができることが実証されています(45,46)。 一方、アロニア多糖類は、その抗高血圧および抗アテローム性動脈硬化作用についてのみ報告されています。 加齢の指標として広く使用されている体重、臓器指数、マロンジアルデヒド(MDA)、スーパーオキシドジスムターゼ(SOD)、カタラーゼ(CAT)のレベルを測定することにより、アロニア多糖類の老化防止効果を評価しました。 D-ガラクトースグループと比較して、アロニア多糖類投与グループは老化したマウスの体重を有意に増加させ、それらの心臓、肝臓、腎臓、および脳はすべて異なるレベルの改善を示しました。 さらに、8アーム迷路テストは、マウスの空間作業と参照記憶を評価するために使用されました。 研究は、アロニア多糖類がマウスの老化と老化したマウスの摂食距離と時間を有意に遅らせることを示しました。 毛皮、精神状態、活動などの外見の観察、および加齢に対するアロニア多糖類の保護効果も実証されました。 H&E分析は、アロニア多糖類投与がD-ガラクトーズ誘発性脳損傷を軽減し、神経機能も改善することを示しました。 |
| 腸内細菌はしばしば「第2のゲノム」と呼ばれます。 それらは、微生物、ゲノムDNA、タンパク質、代謝物など、老化中に体に影響を与える可能性があります(47–49)。 老化したマウスでは、腸内細菌叢の多様性が減少し、有益な細菌が減少し、通性嫌気性菌が増加し、認知機能と記憶機能が低下します(43,44,50,51)。 アロニア多糖類がマウスの微生物叢を改善することによって体の老化状態を緩和したかどうかを判断するために、マウスの糞便を収集し、これらを16SrDNA分析に使用しました。 バクテロイデス属の有益なバクテリアの割合は大幅に増加しました。 加重Unifracクラスタリング(pCoA、ANOSIM)に基づくβ多様性分析は、4つのグループ間の差(R = 0.367、p = 0.001)がグループ内の差よりも有意に大きいことを示しました。 要約すると、アロニア多糖類は腸内細菌叢に明らかな影響を及ぼし、老化研究の新しい分野を切り開きます。 |
| メカニズムの研究では、老化に関連することが示された酸化ストレス、炎症、およびアポトーシスの側面を検討しました。 バイオエナジー代謝の調節における重要な要素として、アデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)は、代謝関連疾患の鍵でもある抗炎症および成長調節において重要な役割を果たします(52,53)。アデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)は、サーチュイン1(SIRT1)、N核因子カッパ-B (F-κB)、哺乳類のラパマイシン標的タンパク質(mTOR)、P53(54)などの複数のシグナル伝達経路にも関連しています。 P53腫瘍抑制タンパク質は、細胞増殖、周期、アポトーシスを調節する多機能転写因子です。 細胞が損傷を受けて癌化すると、p53は細胞の老化を誘発し、腫瘍細胞へのさらなる悪性形質転換を防ぎます。 これは、P53のレベルが55歳まで上昇することを示しており、私たちの研究でもこの現象が確認されています。 |
| ピボットタンパク質として、加齢と加齢に伴う慢性炎症に関連するAMPK / SIRT1 /NF-κBシグナル伝達経路の役割に焦点を当てました。 加齢に伴う核因子カッパ-B (NF-κB)複合体のトランス調節活性の増加、および核因子カッパ-B (NF-κB)活性の増加を伴う慢性炎症との関連、および糖尿病やアテローム性動脈硬化症などの代謝性疾患は、加齢プロセスにおける核因子カッパ-B (NF-κB)シグナル伝達の役割を示唆しています(56)。 メトホルミンは、ヨーロッパでの臨床試験に基づいており、アンチエイジング剤であり、アデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)のアゴニストです。また、抗炎症作用のための核因子カッパ-B (NF-κB)シグナル伝達経路を阻害します(57)。 核因子カッパ-B (NF-κB)はNLRP3インフラマソーム分子の転写と翻訳を促進し、インフラマソーム活性化因子に応答してNLRP3インフラマソームを活性化し、炎症性因子を放出します(58)。 ASCと結合し、IL-1β炎症性因子の成熟を促進し、細胞のピロトーシスを誘発します。炎症性の体はピロトーシスを伴います。 このため、カスパーゼ1とGSDMDの発現レベルも測定しました。 GSDMDは、ピロプトーシスの「死刑執行人」とも呼ばれています(59)。 この結果は、アロニア多糖類がD-ガラクトース誘発老化マウスの脳組織のインフラマソームに有意な抑制効果をもたらし、それによって老化のプロセスを延期することを示しており、これは以前の研究結果と一致していました(60)。 興味深いことに、この研究は、N核因子カッパ-B (NF-κB)に対するアデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)の間接的な調節効果を持っていたアデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)経路を調べることによって結果を検証しました。 この結果は、加齢に伴うアデノシン5'-リン酸活性化プロテインキナーゼ(AMPK)の活性の低下を説明し、エネルギー代謝の不均衡と炎症反応を引き起こす可能性があります(61)。 |
| 酸化ストレスは、神経変性疾患の病因における重要な要因です(62)。核関連因子-2(Nrf2)は酸化ストレスを調節する重要な転写因子であり、通常の生理学的条件下で細胞質に存在します。活性化されると、核に入り、下流のタンパク質の発現を調節し、抗炎症効果があります(63)。ヘムオキシゲナーゼ-1(HO-1)は、体内で最も広く分布している抗酸化酵素の1つであり、抗炎症作用があります(64,65)。 我々は、アロニア多糖類投与が核関連因子-2核(Nrf2 Nuclear)への参入を促進し、ヘムオキシゲナーゼ-1(HO-1)の発現をアップレギュレーションすることを発見しました。 酸化に関連する指標(マロンアルデヒド(MDA)、スーパーオキシドジスムターゼ(SOD)、カタラーゼ(CAT))をテストしたところ、結果は予想と同じでした。 PI3K / Akt / mTOR経路は、核関連因子-2(Nrf2)核転座と第2段階の抗酸化酵素の発現を調節するための最も重要な上流シグナルの1つです(66–68)。PI3K / Akt / mTOR経路関連タンパク質を探索し、下流のアポトーシスタンパク質をテストしました。 BaxやBcl-2を含むアポトーシスタンパク質ファミリーは、細胞増殖とアポトーシスの典型的な調節因子です(69)。この研究では、アロニア多糖類はP-PI3KとP-AKTの発現を増加させ、Baxとカスパーゼ3の発現を抑制し、PI3K / AKT経路を介してBcl-2の発現を増強しました。これらのデータは、アロニア多糖類がPI3K / AKT / mTORシグナル伝達経路を調節し、老化プロセスを遅らせることにより、老化した脳組織におけるD-ガラクトース誘発性の酸化ストレス損傷を有意に改善し、細胞アポトーシスを阻害したことを示しました。 |
| 結論として、アロニア多糖類は、マウスの基本的な特性と指標、行動空間、学習、記憶機能、および腸内細菌叢の分析によって、アンチエイジングに有効であることが確認されました。 酸化ストレス、炎症、アポトーシス、その他の側面で考えられるメカニズムを分析しました。 アロニア多糖類は、AMPK / SIRT1 /NF-κBシグナル伝達経路によってNLRP3インフラマソームを阻害し、マウスの老化の腸内細菌叢の遅延を調節しました。 これらの証拠は、老化を遅らせるためのアロニア多糖類の潜在的なメカニズムを明らかにしました。 しかし、老化は全身の包括的な症状であり、この研究は脳のみを検出し、他の臓器へのアロニア多糖類の影響はさらなる研究のままです。 |
| 化学薬品および材料 |
| アスパラギン酸アミノトランスフェラーゼ(AST)、アラニンアミノトランスフェラーゼ(ALT)、還元型グルタチオン(GSH)、スーパーオキシドジスムターゼ(SOD)、およびマロンジアルデヒド(MDA)スレオニンは、南京江城生物工学研究所リグニン-エオシン(H&E)市販アッセイキットから入手しました。 D-Gal(純度≥95%)はSigma(セントルイス、ミズーリ州、米国)から購入しました。 ウエスタンブロット関連の抗体は、Proteintech and Arigo(Changchun、China)から購入しました。この研究で使用した他のすべての化学物質と試薬は、分析グレードのものでした。 |
| アロニア多糖類の準備 |
| アロニア多糖類の分離と精製新鮮な果物を洗浄し、蒸留水で煎じて多糖類を抽出しました。煎じ薬を元の容量の8分の1に濃縮し、次に95%エタノールの3倍の容量を加えて、4°Cの冷蔵庫で一晩沈殿させて沈殿物を得ました。水を加えて溶解し、脱脂、色素脱失、マクロポーラス樹脂D101による予備精製、恒量までの真空凍結乾燥、AMPとしての記録を経て、アルコール沈殿を3回繰り返して粗多糖類を得た。マクロポーラスレジンで予備精製した後、AMP 1 gを脱イオン水に溶解し、脱イオン水(流速1.0 mL / min)をDEAE-52ファイバーカラム(3×40cm)に通して溶出しました。 DEAE-52セルロースの直径は300メッシュでした。溶出液は自動コレクターによって収集されました。各試験管(10 mL)は、10分ごとに収集され、合計150本になりました。溶出液中の多糖類は硫酸フェノール法により検出された。吸光度の値を測定し、溶出曲線を描いてMWPの組成を決定しました。真空凍結乾燥後、淡褐色-黄色の粉末が得られ、これをその後の構造同定に使用した。 |
| フーリエ変換赤外(FT-IR)分光分析 |
| サンプルの重さを2mgにし、サンプルと臭化カリウム200mgを圧搾して錠剤にしました。 ブランクの対照錠剤は、臭化カリウム粉末のみを使用してプレスされた。 サンプルをスキャンし、フーリエ変換赤外分光計(JASCO FT / IR-620分光計; JASCO、八王子、日本)で記録しました。 |
| 単糖組成分析 |
| アロニア多糖類の単糖組成は、イオンクロマトグラフィーによって決定された。 5 mgのサンプルとTFAを121°Cで2時間加熱し、窒素で乾燥させ、残留TFAをメタノールで除去し、これを3回繰り返しました。 |
| クロマトグラフィーシステムは、Thermo ICS5000 +イオンクロマトグラフィーシステム(ThermoFisher Scientific、USA)、Dionex™CarboPac™PA10(250×4.0 mm、10μm)液体クロマトグラフィーカラムを使用しました。 注入量は20μL、カラム温度は30°Cでした。 移動相A(H2O)および移動相B(100 mMNaOH)。 |
| フコース(Fuc)、ラムノース(Rha)、アラビノース(Ara)、ガラクトース(Gal)、グルコース(Glc)、キシロース(Xyl)、マンノース(Man)、ガラクツロン酸(Gal-UA)、グルクロン酸(Glc-UA) 、およびマンヌロン酸(Man-UA)を標準的な単糖として使用しました。 それらの処理方法は、サンプル分析に使用されたものと同じでした。 定性分析と定量分析は、クロマトグラフィーピークの保持時間に応じて調整されました。 |
| 多糖類の老化防止活性の分析 |
| 動物と実験計画合計40匹の成体雄ICRマウス(6〜8週齢、体重22〜25 g)を、品質証明書SCXK(JI)2019-0008(Changchun)を付けてChangchun Yisi Experimental Animal Co.、Ltd。から購入しました。 、 中国)。 マウスは、12:12時間(L:D)の光周期で病原体のない条件下で一定の温度と湿度の下に維持され、食物と水に無制限にアクセスできました。 すべての動物調査プロセスは、実験動物の注意と使用に関するガイドに従って行われ、吉林農業大学の動物調査道徳委員会によって許可されました。 倫理承認番号は2019-08-28-001でした。 |
| 図2Aに示すように、実験操作の基本的な流れを確立しました。 適応の3週間後、マウスをランダムに4つのグループに分けました:正常グループ、D-Galグループ、AMP低用量グループ、高用量グループ、各グループに10匹のマウス。 D-Gal群では、腹腔内注射により200 mg / kg増量し、1000 mg / kgに増量して同じ用量を維持し、3日1回12週間、AMP(100および200 mg)を投与しました。 / kg)は、6週間のD-Gal誘発後、強制経口投与により毎日投与されました。 同量の生理食塩水が正常群に与えられた。最後の投与から24時間後、すべてのマウスを処刑し、解剖して血液上清を得ました。これは、4°Cで10分間の遠心分離後に分離され、分析まで-80°Cで保存されました。脳、心臓、腎臓、脾臓、肝臓の組織を採取して、臓器指数を計算します。臓器重量指数=臓器重量(mg)/体重(kg)。生理食塩水を1:9の比率で使用し、粉砕組織グラインダーを4°Cで使用しました。分離したホモジネート上清を4000×g、4°Cで10分間遠心分離し、-80°Cで保存してコリン作動性機能を測定しました。脳組織サンプルは組織学的検査のためにホルマリンで固定され、他の切片は液体窒素で急速に凍結保存されました。 |
| 組織の収集とサンプルの準備マウスの初期および最終重量をマウスの糞便とともに収集し、すぐに液体窒素に入れ、その後の腸内細菌叢分析のために凍結しました。 12時間絶食した後、軌道採血と塩基性酸化指数の測定により採血しました。 脳はすぐに取り出され、生化学的分析のために-80°Cで保存されました。 マウスの臓器を収集し、臓器指数分析のために秤量した。 |
| 生化学的分析 |
| 新鮮な血液を3000rpm、4°Cで30分間遠心分離し、血清を収集し、グルタチオン(GSH)、スーパーオキシドジスムターゼ(SOD)、およびマロンジアルデヒド(MDA)の含有量を使用して酸化を評価しました。 マウスの脳を0.9%生理食塩水でホモジナイズし、アセチルコリンエステラーゼ(AchE)含有量を測定して、脳内のニューロンの損傷を評価しました。 |
| 組織学的分析 |
| マウスを解剖した後、マウスの新鮮な脳を採取し、パラフィンに包埋し、製造元の指示に従ってヘマトキシリンおよびエオシン(H&E)染色キットを使用して検出しました。 結果は、光学顕微鏡(Leica DM750、ドイツ)を使用して得られました。 |
| ウエスタンブロット |
| タンパク質抽出キット(Thermo)を使用して、脳の上清を取得しました。 次に、サンプルをSDSポリアクリルアミドゲルで分離し、PVDFメンブレンに転写し、BSAで1.5時間ブロックし、TBSTで5分間3回洗浄し、さまざまな抗体と一晩インキュベートしました。 メンブレンをTBSTで3回、毎回5分間リンスした後、HRP標識抗体と1時間インキュベートしました。 バンドはECL化学発光溶液によって検出され、タンパク質発現はBio-Rad VersaDoc 3000 Imaging System(Bio-Rad、Mississauga、ON、Canada)によって表示されました。 元の画像は補足情報で取得されました。 |
| 行動テスト |
| マウスの空間と学習能力をテストするために、8本の腕の迷路が使用されました。 テストの前に、環境に適応するために一定時間マウスを順応させました。 それらは12時間絶食され、その後10分間テストされ、4つのアームが砂糖の錠剤の誘導のためにランダムに選択されました。 RMT-100分析ソフトウェア(Chengdu Taimeng Software Co.、Ltd。)を使用してデータを収集し、運動軌道パラメーターを記録しました。 実験中にマウスが移動した距離、潜伏期間を分析し、マウスが各象限を通過する時間/合計時間を計算しました。 |
| 16SrDNAハイスループットシーケンス |
| 腸内細菌叢分析のために、すべてのグループから合計20の糞便サンプルがランダムに選択されました(グループあたり5つのサンプル)。 16S V4領域のハイスループットシーケンスを使用して、腸内細菌叢を分析および比較しました。 Uparseソフトウェアを使用して、97%の類似性のOTUしきい値に基づいて、シーケンスは操作上の分類法(OTU)にクラスター化されました。 シーケンスリードとOTUに基づいて、αダイバーシティ、βダイバーシティ、線形判別分析の効果量(LEfSe)を分析しました。 |
| 統計分析 |
| 統計分析は、SPSS統計ソフトウェアを使用して実行されました。 数値比較は、2つのグループ間の差異を分析するためのANOVAテストによって実行されました。 結果は平均±SDとして示され、有意水準はp <0.05として定義されます。 |
| 参考文献(本文中の文献No.は原論文の文献No.と一致していますので、下記の論文名をクリックして、原論文に記載されている文献を参考にしてください) |
| この文献は、Scientifc Reports | (2021) 11:20558に掲載されたAronia melanocarpa polysaccharide ameliorates infammation and aging in mice by modulating theAMPK/SIRT1/NF κB signaling pathway and gut microbiota. を日本語に訳したものです。タイトルをクリックして原文を読むことが出来ます。 |